
Abstract
Tumor necrosis factor α (TNFα) and other members of the TNF family affect adipose tissue metabolism and contribute to the obesity-related inflammation of adipose tissue. Here, we sought to identify the effects of TRAIL (TNF-related apoptosis-inducing ligand) on fat cell biology. TRAIL-receptor 2 (TRAIL-R2) and its mouse homolog DR5 were regulated upon acute and chronic energy imbalance in murine and human adipose tissue. TRAIL inhibited insulin-stimulated glucose uptake and de novo lipogenesis in human adipocytes. Interestingly, TRAIL did not interfere with the phosphorylation of insulin-stimulated kinases such as Akt or Erk and did not activate the NF-κB pathway. Instead, TRAIL activated cleavage of caspase-8 and caspase-3. The subsequent cleavage of PPARγ led to its inactivation and resulted in reduced expression of lipogenic genes, such as Glut-4, FASN, and ACC. Taken together, we discovered a so far unknown function of the death ligand TRAIL in regulating adipocyte metabolism. Our results imply that TRAIL/TRAIL-R system might provide a new target for the prevention and treatment of obesity and its co-morbidities.
Similar content being viewed by others
Main
Long regarded as a mere storage organ, white adipose tissue (WAT) is now recognized as a highly active endocrine organ.1 It secretes a plethora of different factors, including hormones and inflammatory cytokines, and thereby regulates metabolism locally and systemically.2 Adipose tissue is also a target for several hormones and cytokines, which control its physiological functions, that is, lipid storage and fatty acid release. Any disturbance of these processes can eventually lead to the co-morbidities widely observed to be associated with obesity, especially insulin resistance and type 2 diabetes mellitus.3 In obesity, the abnormally increased adipose tissue mass is accompanied by local inflammation and characterized by increased infiltration of immune cells and enhanced production of inflammatory cytokines.4 This leads to activation of inflammatory pathways in adipocytes and might contribute to both local and systemic insulin resistance.5
Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) is a member of the tumor necrosis factor (TNF) family.6 Upon binding to its receptors (R1 and R2) the death-inducing signaling complex (DISC) is assembled. In turn, DISC formation leads to cleavage and activation of caspases and finally to induction of apoptosis.7 As TRAIL selectively induces apoptosis in cancer cells, but not in non-malignant cells, it is regarded as a promising anticancer agent and its safety and efficacy is currently investigated in several phase I/II studies.8, 9 TRAIL as well as it receptors are expressed in many normal tissues10 suggesting additional, so far uncharacterized, biological functions.
Members of the TNF family have been implicated in the pathogenesis of obesity-related disorders.11 The finding that TNFα is overexpressed in obese WAT and is involved in the development of insulin resistance revolutionized the field of obesity research almost two decades ago.12 Recently, Wueest et al.13 demonstrated that adipocyte-specific deletion of CD95 results in improved glucose homeostasis, less triglyceride accumulation in the liver, and decreased systemic inflammation when the mice were challenged with a high-fat diet. Interestingly, elevated serum concentrations of TRAIL are associated with serum lipid levels and adiposity in humans.14
Based on these findings, we hypothesized that TRAIL may exert so far unknown effects on adipose tissue. We showed previously that human preadipocytes and adipocytes are resistant to TRAIL-induced apoptosis.15, 16 In this study, we investigated the impact of TRAIL on adipocyte metabolism.
Results
Regulation of expression of TRAIL and its receptors in mice and humans by acute and chronic energy imbalance
We analyzed the expression of TRAIL and its receptors (TRAIL-R1 and R2) in adipose tissue during different states of energy imbalance. Data from a searchable microarray gene expression database for mouse models of obesity17 revealed that Trail as well as Dr5 (homolog to human TRAIL-R2) mRNA were significantly increased in genetically obese ob/ob mice (Figure 1a). We next challenged wild-type FVB mice with an overnight fast followed by re-feeding. Both Trail and Dr5 were significantly downregulated by fasting in the inguinal fat depot (Figure 1b). Expression levels returned to initial values within 2 h of re-feeding.

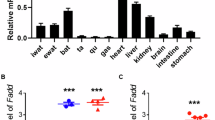
Expression of TRAIL and its receptors are regulated by chronic and acute energy imbalance. (a) Expression of Trail and Dr5 (mouse homolog to human TRAIL-R2) in adipose tissue of wild-type and ob/ob mice. Data were obtained from microarray gene expression database.17 (b) Total RNA of inguinal white adipose tissue of fed (n=5), overnight fasted (n=5), and re-fed (n=5) mice was prepared and reversely transcribed. Expression levels of Trail and Dr5 were analyzed with qPCR and normalized to Hprt. Data are the mean+S.E.M. *P<0.05 (fasted versus fed or re-fed). Total RNA of subcutaneous adipose tissue of 10 patients (age 40–64, BMI 20–67 kg/m2) was prepared and reversely transcribed. Expression levels of (c) TRAIL and its receptors ((d) TRAIL-R1, (e) TRAIL-R2) were measured with qPCR and normalized to SDHA (succinate dehydrogenase complex, subunit A) followed by a Spearman rank order correlation analysis with the BMI. Correlation coefficient (R) and P-values are given in the respective graphs. Expression of (d) TRAIL-R1 and (e) TRAIL-R2 correlated positively with BMI
We prepared total mRNA from subcutaneous adipose tissue of 10 patients (age 40–64, body mass index (BMI) 20–67 kg/m2) and analyzed TRAIL, TRAIL-R1, and TRAIL-R2 expression by qPCR followed by a spearman correlation analysis with BMI of the patients. In contrast to the murine data, TRAIL expression in human adipose tissue was not correlated with obesity (Figure 1c). However, we found a strong, positive correlation of TRAIL-R1 (R=0.685, P=0.252; Figure 1d) and TRAIL-R2 (R=0.644, P=0.0377; Figure 1e) with BMI.
This set of data clearly demonstrates that the expression of TRAIL receptors in the adipose tissue is associated with the nutritional status of the body underlining our hypothesis of a putative role of the TRAIL/TRAIL-R system in adipocyte metabolism.
TRAIL receptors are expressed in human preadipocytes and adipocytes
To investigate the effects of TRAIL on adipocyte metabolism, we used the human preadipocyte cell strain SGBS (Simpsom Golabi Behmel syndrome), a well characterized in vitro model of human adipocyte biology.15, 16 TRAIL-R1 and TRAIL-R2 mRNA was expressed in SGBS preadipocytes and downregulated upon differentiation into adipocytes (Figures 2a and b, for detailed expression analysis, see Supplementary Figure S1). The same expression pattern was detected in human primary preadipocytes and adipocytes isolated from three different donors (Figures 2c and d). Both receptors were present at the cellular surface of preadipocytes as measured by flow cytometry (Figure 2e). Although TRAIL-R1 was absent in mature adipocytes, TRAIL-R2 was clearly present, but downregulated by∼47% compared with precursor cells.

TRAIL receptors are expressed in human preadipocytes and adipocytes. SGBS preadipocytes were differentiated into adipocytes. Total RNA was prepared on d0 (preadipo) and d14 (adipo) and reversely transcribed. qPCR analysis was performed using primer pairs specific for (a) TRAIL-R1 and (b) TRAIL-R2. Data are represented as relative amount of mRNA expression normalized to GAPDH and are presented as mean+S.E.M. of three independent experiments. *P<0.05 (preadipo versus adipo). Primary preadipocytes were isolated from subcutaneous adipose tissue of three different donors and differentiated into adipocytes for 14 days. (c) TRAIL-R1, and (d) TRAIL-R2 mRNA expression was analyzed by qPCR. Data are expressed as mean±S.E.M. of three different donors. *P<0.05 (preadipo versus adipo). (e) Surface expression levels of TRAIL-R1 (left panel) and TRAIL R2 (right panel) were analyzed by flow cytometry for SGBS preadipocytes (upper panel) and adipocytes (lower panel). Black: isotype control, blue: TRAIL-R1, red: TRAIL-R2
TRAIL affects insulin-mediated metabolic functions of fat cells via TRAIL-R2
To study the effects of TRAIL on important insulin-stimulated metabolic pathways of fat cells, we pre-treated adipocytes with TRAIL for 24 h. As a result, insulin-stimulated glucose uptake was significantly decreased by 45%±9 with 100 ng/ml TRAIL (Figure 3a). De novo lipogenesis, studied by measuring the incorporation of radioactively labeled, metabolizable glucose into cellular lipids, was significantly decreased with 100 ng/ml TRAIL (55%±8; Figure 3a). A comparable effect was detected in human primary adipocytes differentiated ex vivo (Supplementary Figure S2). Both basal glucose uptake as well as basal lipogenesis was not affected by TRAIL treatment (Supplementary Figure S3).

TRAIL inhibits glucose uptake and de novo lipogenesis in human fat cells via TRAIL-R2. SGBS adipocytes were treated with increasing doses of TRAIL for 24 h. (a) Glucose uptake was stimulated with 10−8M insulin for 15 min. The cellular uptake of 14C-deoxy-glucose uptake was measured on a β-counter. Results are presented as percentage of control and the mean+S.E.M. of five independent, triplicate experiments. *P<0.05 (TRAIL versus vehicle control). (b) SGBS adipocytes were treated with increasing doses of TRAIL. After 24 h, 10−8M insulin and 14C-glucose was added for another 24 h. The incorporation of 14C-glucose into cellular lipids was measured. Results are presented as percentage of control and the mean+S.E.M. of five independent, triplicate experiments. *P<0.05 (TRAIL versus vehicle control). SGBS adipocytes were treated with 10 μg/ml agonistic antibodies against TRAIL-R1 (mapatumumab, Mapa) or TRAIL-R2 (lexatumumab, Lexa). (c) Insulin-stimulated glucose uptake and (d) de novo lipogenesis was determined as described above. Results are presented as percentage of control and the mean+S.E.M. of three independent, quadruplicate experiments. *P<0.05 (Mapa or Lexa versus vehicle control)
To elucidate which TRAIL receptor mediates the effect on adipocyte metabolism, we used specific, agonistic antibodies for TRAIL-R1 (HGS-ETR1, mapatumumab) or TRAIL-R2 (HGS-ETR2, lexatumumab). These antibodies are currently tested for anticancer activity in phase I/II studies.18 Insulin-stimulated glucose uptake was unaffected when TRAIL-R1 was targeted with mapatumumab (Figure 3c). When adipocytes were pretreated with the TRAIL-R2 agonist, lexatumumab, we observed a reduced insulin-stimulated glucose uptake (inhibition by 28%±8; Figure 3c). In line with this finding, the insulin-stimulated de novo lipogenesis was only inhibited by TRAIL-R2 stimulation (34%±9; Figure 3d). With this set of experiments, we identified TRAIL-R2 as the receptor responsible for mediating the TRAIL-related effects on adipocyte metabolism.
TRAIL-mediated effects on adipocyte metabolism are independent of nuclear factor kappa B (NF-κB)
NF-κB is a key molecule in TNFα signaling in many cell types.19 In murine adipocytes, TNFα leads to a downregulation of adipocyte-specific genes, thereby causing insulin resistance; the activation of NF-κB is critically involved in these processes.20 We therefore wanted to elucidate if TRAIL mediates its effects on metabolism via NF-κB-activated changes in gene expression. Indeed, we detected a significant downregulation of genes responsible for glucose uptake and lipogenesis, including glucose transporter-4 (Glut-4), acetyl CoA-carboxylase (ACC), and fatty acid synthase (FASN) after 24 (Supplementary Figure S4) and 48 h of TRAIL treatment (Figure 4a). There was no sign of TRAIL-induced nuclear translocation of p65 (Figure 4b) as well as no evidence of TRAIL-induced NF-κB activation in human adipocytes, while TNFα clearly caused a shift in the electrophoretic mobility shift assay (Figure 4c).

TRAIL effects on adipocyte metabolism are independent of NF-κB and insulin-stimulated kinases. SGBS adipocytes were incubated with vehicle or increasing concentrations of TRAIL. (a) After 48 h total RNA was prepared and reversely transcribed. qPCR was performed using specific primers for Glut-4, ACC, and FASN. Results are standardized to GAPDH and related to vehicle control. Data are presented as mean+S.E.M. of five independent experiments. *P<0.05 (TRAIL versus vehicle control). (b) SGBS adipocytes were treated with 100 ng/ml TRAIL or 100 ng/ml TNFα for 6 h. Then, p65 translocation to nucleus was analyzed by immunofluorescence microscopy. One representative experiment out of three performed is shown. (c) DNA binding activity of NF-κB after stimulation with 100 ng/ml TRAIL for 30 or 120 min or 100 ng/ml TNFα for 120 min was determined by electrophoretic mobility shift assay. One representative experiment of three performed is shown. (d and e) SGBS adipocytes were incubated with vehicle or 100 ng/ml TRAIL. After 24 h, cells were stimulated for 15 min with or without 10−8M insulin and protein lysates were prepared. Western blot analysis was performed to study the phosphorylation of known insulin-stimulated kinases Akt, ERK, and p38 (d) and TNFα-stimulated kinases JNK and AMPK (e). One representative experiment out of three performed is shown
Phosphorylation of insulin-stimulated kinases is not affected by TRAIL
TNFα induces insulin resistance in adipocytes via multiple pathways, including the inhibition of expression and/or phosphorylation of insulin-stimulated kinases.21, 22 Likewise, CD95 triggering downregulates Akt expression and activity in 3T3-L1 adipocytes.23 Interestingly, neither expression nor phosphorylation of typical insulin-activated kinases such as Akt, p38 mitogen-activated protein kinase (MAPK), or ERK (p42/44 MAPK) was significantly affected by treatment with TRAIL as judged by western blot analysis with phosphor-specific antibodies (Figure 4d). TNFα induces phosphorylation of AMP-activated protein kinase (AMPK) and c-Jun N-terminal kinase (JNK), thereby contributing to the development of insulin resistance.24 However, we did not find comparable significant effects of TRAIL on AMPK and JNK phosphorylation (Figure 4e). Additional performed MAPK phosphorylation arrays did not reveal any significant changes related to phosphorylation of intracellular kinases (Supplementary Figure S5).
Caspases are involved in mediating the effects of TRAIL on adipocyte metabolism
Canonically, TRAIL binding to its cognate receptor is thought to initiate apoptosis induction, resulting in receptor trimerization and formation of the DISC, subsequently leading to cleavage of caspases and finally ending up in cell death.7 We knew from earlier studies that TRAIL causes formation of the DISC without inducing apoptosis, which was in line with the finding that TRAIL only induces apoptosis in malignant cells.16 We now hypothesized that caspases might be involved in the TRAIL-induced inhibition of insulin-stimulated metabolism in fat cells. We therefore incubated SGBS adipocytes with the pan-caspase inhibitor zVAD.fmk during TRAIL treatment. Notably, the inhibitory effect of TRAIL on insulin-stimulated glucose uptake and lipogenesis was completely abolished by the caspase inhibitor (Figures 5a and b). Likewise, the inhibition of Glut-4, ACC, and FASN mRNA expression by TRAIL was abrogated in the presence of zVAD.fmk (Figure 5c).

TRAIL-induced effects on adipocyte metabolism are caspase dependent. (a) SGBS adipocytes were incubated with 100 ng/ml TRAIL in the presence or absence of 10 μM zVAD.fmk. Glucose uptake was stimulated with 10−8 M insulin for 15 min. The cellular uptake of 14C-deoxy-glucose uptake was measured on a β-counter. Data are presented as mean+S.E.M. of four independent, triplicate experiments. *P<0.05 (zVAD+TRAIL versus TRAIL). (b) SGBS adipocytes were treated with 100 ng/ml TRAIL in the presence or absence of 10 μM zVAD. After 24 h, 10−8 M insulin and 14C-glucose was added for another 24 h. The incorporation of 14C-glucose into cellular lipids was measured. Data are presented as mean+S.E.M. of four independent, triplicate experiments. *P<0.05 (zVAD+TRAIL versus TRAIL). (c) SGBS adipocytes were incubated with 100 ng/ml TRAIL in the presence or absence of zVAD (10 μM) for 48 h. qPCR analysis was performed using primer pairs specific for Glut-4, ACC, and FASN. Data are represented as relative amount of mRNA expression normalized to GAPDH are presented as mean+S.E.M. of three independent experiments.*P<0.05 (TRAIL versus TRAIL+zVAD). (d) SGBS adipocytes were treated with 100 ng/ml TRAIL. Protein lysate were prepared after indicated time points and subjected to western blot analysis for caspases-1, -3, -6, and -8. One representative experiment out of three performed is shown
These observations raise the question which caspase is critically involved in mediating these effects of TRAIL. We therefore monitored cleavage of several caspases known to be functionally important in adipocytes.25, 26, 27 In line with our previous work, TRAIL induced cleavage of caspase-8 into the p43/41 intermediate and the active p18 fragment within 30 min (Figure 5d). Cleavage of caspase-3 into the active p17 fragment was evident at 2–6 h. No difference was detectable between control and TRAIL treatments on caspase-1 or caspase-6 (Figure 5d). It is important to mention that there is no significant induction of apoptosis (<8%) even after long-term stimulation for 72 h with 100 ng/ml TRAIL (Supplementary Figure S6).
Caspase-8 and caspase-3 mediate the effect of TRAIL on adipocyte metabolism
To verify whether caspase-8 and caspase-3 are involved in mediating the effects of TRAIL on metabolism, we took advantage of lentiviral-mediated overexpression of shRNA in order to knock down the expression levels of both caspases. Using two different constructs for each caspase, we were able to downregulate the expression of caspase-3 and caspase-8 by∼80% in mature SGBS adipocytes compared with control cells overexpressing a nonsense shRNA sequence (Figure 6a). Although TRAIL significantly inhibited de novo lipogenesis in hyper random sequence control cells by∼25%, this inhibitory effect was totally blunted in capase-3- and caspase-8-deficient adipocytes (Figure 6b), proving that caspase-8 and caspase-3 mediate the effects of TRAIL on adipocyte metabolism.

Knock down of caspase-8 and caspase-3 overcomes the effect of TRAIL on de novo lipogenesis. SGBS adipocytes were transduced with lentivirus containing either a nonsense shRNA sequence (ctrl) or shRNA targeting human caspase-3 (C3.1 and C3.2) and caspase-8 (C8.1 and C8.2). (a) Knock down of caspase-3 and caspase-8 was analyzed by western blot. (b) Nonsense controls (ctrl) and caspase-3- or caspase-8-deficient adipocytes were treated with 100 ng/ml TRAIL. After 24 h, 10−8 M insulin and 14C-glucose was added for another 24 h. The incorporation of 14C-glucose into cellular lipids was measured. Results are presented as percentage of insulin-stimulated cells, which were set to 100%. Data are shown as mean+S.E.M. of three independent, triplicate experiments. *P<0.05 (insulin versus insulin+TRAIL)
TRAIL induces caspase-mediated cleavage of peroxisome proliferator-activated receptor gamma (PPARγ) and FASN
Next, we aimed to elucidate the critical downstream events upon TRAIL-induced caspase activation. Recently, caspase-mediated cleavage of PPARγ was reported to be an important aspect of TNFα-induced insulin resistance in adipose tissue.25, 26, 27 Indeed, we detected a∼47 kDa cleavage product of PPARγ at 2–6 h of TRAIL treatment (Figure 7a). This cleavage product disappeared at later time points, accompanied by a downregulation of full-length PPARγ. As expected from mRNA expression data (Figure 4a), Glut-4 protein expression was downregulated upon TRAIL treatment. In addition, we detected a so far unknown cleavage product of FASN with an estimated size of ∼100 kDa. Potential cleavage sites of FASN are summarized in Supplementary Table S1. Both the cleavage of PPARγ and FASN were inhibited by the pan-caspase inhibitor zVAD.fmk (Figure 7b). As expected from Guilherme et al.,26 the cleavage of PPARγ resulted in alteration of its subcellular localization. Although PPARγ was located in the nucleus in control cells, it was dispersed in the cytosolic compartment in TRAIL-treated cells (Figure 7c). Cytosolic PPARγ co-localized with the cleaved form of caspase-3, further underlining the role of caspase-3 in PPARγ cleavage. Taken together, these experiments demonstrate that cleavage of PPARγ leads to its inactivation and is accompanied by nuclear export and cytosolic degradation.26

TRAIL induces caspase-mediated cleavage of PPARγ and FASN. (a) SGBS adipocytes were treated with 100 ng/ml TRAIL. Protein lysate were prepared after indicated timepoints and subjected to western blot analysis for PPARγ, FASN, and Glut-4. One of three performed experiments is shown. (b) SGBS adipocytes were treated with 100 ng/ml TRAIL in the presence or absence of the caspases-inhibitor zVAD (10 μM). After 6 h, protein lysate was prepared and subjected to western blot analysis for PPARγ and FASN. One experiment out of three performed is shown. (c) SGBS adipocytes (day 14) grown on chamber slides were incubated with vehicle or 100 ng/ml TRAIL. After 6 h, cells were fixed, permeabilized, and stained with anti-PPARγ, anti-active caspase-3, and DAPI to visualize endogenous PPARγ (red) and active caspase-3 (green) and nucleus (blue). Upper rows magnification 100-fold, lower row magnification 400-fold
Discussion
In the present study, we discovered a so far unknown function of the death ligand TRAIL in regulating human adipocyte metabolism. By binding to TRAIL-R2, it activates cleavage of caspase-8 and caspase-3, which in turn cleaves and inactivates PPARγ resulting in reduced expression of lipogenic genes, such as Glut-4, FASN, and ACC. Consequently, insulin-stimulated processes such as glucose uptake and lipogenesis are significantly inhibited. Most interestingly, TRAIL-R2 and its mouse homolog DR5 are regulated upon acute and chronic energy imbalance in murine and human adipose tissue.
TRAIL was identified in 1995/96 by two independent groups (for review, see Koschny et al.8). Together with other death ligands of the TNF superfamily such as TNFα and CD95L, it regulates immune responses, in part, by inducing apoptosis. As summarized by Falschlehner et al.,28 the TRAIL/TRAIL-R system has immunoregulatory, immunosuppressive, pro-viral or anti-viral, and tumor immunosurveillance functions. TRAIL is expressed in immune cells such as macrophages, T cells, NK cells, and dendritic cells.28 As it selectively kills transformed cells sparing non-malignant cells, TRAIL received enormous attention as a therapeutic agent for the treatment of cancer.29 TRAIL receptors are expressed in a variety of cells and tissues (for more information, see www.proteinatlas.org). In this study, we demonstrate that TRAIL-R2 is expressed on the cellular surface of human preadipocytes and is still present in significant amounts on lipid-laden adipocytes, where TRAIL-R1 is absent. Therefore it was not surprising that only the TRAIL-R2 agonist, lexatumumab, and not the TRAIL-R1 agonist, mapatumumab, mimicked the effects of TRAIL on adipocyte metabolism.
TRAIL serum levels are increased in human obesity.14 Since obesity is characterized by the excessive accumulation of WAT, this prompted us to study the effects of TRAIL on adipocyte metabolism. In our in vitro studies, activation of TRAIL-R2 with recombinant human TRAIL resulted in a robust inhibition of insulin-stimulated glucose uptake and insulin-stimulated de novo lipogenesis. To our knowledge, this is the first study demonstrating direct metabolic effects of this death ligand on adipocytes at the cellular level. Earlier studies focused on the functions of TRAIL in the immune system and its anti-cancer properties. Only recently, TRAIL was also implicated in proliferation and differentiation of non-immune cells30, 31, 32. For instance, TRAIL promotes survival, migration, and proliferation of vascular smooth muscle cells31 or stimulates human intestinal cell differentiation.32 Some studies investigated the role of TRAIL in type 1 diabetes mellitus, which is characterized by autoimmune destruction of insulin-producing β-cells in the pancreas. In NOD mice, blocking TRAIL function by a soluble TRAIL receptor increased the onset of diabetes.33, 34 TRAIL knockout mice were more susceptible to streptozotocin-induced diabetes and displayed enhanced islet inflammation compared with wild-type animals.33 From these findings, Lamhamedi-Cherradi et al.33 concluded that TRAIL deficiency accelerates autoimmune type 1 diabetes and enhances autoimmune responses in the pancreas. However, the role of TRAIL in the pathogenesis of type 2 diabetes mellitus has not been addressed so far. The levels of circulating TRAIL correlate with total body fat and serum lipid levels.14 In a recent study, the serum concentrations of TRAIL correlated positively with energy intake and energy balance.35 These clinical findings show that TRAIL is linked to adiposity, and the question arises whether it is causally connected to the development of obesity-induced insulin resistance. Our data demonstrates that TRAIL induces insulin resistance in adipocytes in vitro. Adipose tissue insulin resistance is an early event in the pathogenesis of obesity-related systemic insulin resistance. We claim that TRAIL is an important player in these processes.
In human adipocytes, TRAIL induced cleavage and activation of the initiator caspase-8 and the effector caspase-3. We knew from earlier studies that TRAIL causes formation of the DISC,16 which is usually the first step of the apoptosis cascade. Despite subsequent cleavage of caspase-8 and -3, there is no induction of apoptosis upon selective TRAIL-R activation alone. Additional sensitization by either blocking protein biosynthesis with cycloheximide or incubation with the phytochemical resveratrol is needed to finally execute the apoptotic program.15, 16 Resistance to death receptor-mediated apoptosis in human fat cells is mediated by an auto-/paracrine insulin-like growth factor-1 (IGF-1)/IGF-1 receptor loop.15 Locally, high levels of IGF-1 lead to activation of intracellular survival pathways, including the PI3K/Akt signaling cascade and induce the expression of anti-apoptotic proteins, such as Fas-associated death domain-like interleukin-1 beta-converting enzyme-inhibitory protein (c-FLIP).15, 36 Thus, adipocytes possess an efficient survival machinery to prevent apoptosis. However, once activated, caspases have the ability to cleave other substrates, such as PPARγ. Furthermore, we identified FASN as a new caspase substrate. Our results suggest that cleavage of FASN negatively regulates its function and contributes to its downregulation. Additional experiments will be required to clarify this.
Caspase-mediated cleavage of PPARγ, induced by TNFα, was previously reported by two groups.25, 26 Using specific inhibitors He et al.25 identified caspase-1 as responsible for this cleavage, while data from Guilherme et al.26 point to a role of caspase-8, -3, and -6. In our study with human adipocytes, TRAIL only stimulated cleavage of caspase-8 and -3. As expected,26 PPARγ was no longer located to the nucleus which has two important consequences: (A) it no longer activates transcription of genes important for glucose uptake and lipogenesis; and (B) once transported to the cytosol, it is degraded in the proteasome.26
We identified TRAIL as a new player in the development of adipose tissue insulin resistance as it inhibited insulin-stimulated glucose uptake and lipogenesis. This is well conceivable as TRAIL belongs to the TNF superfamily and shares not only 23% of sequence homology with TNFα, but also marked similarities in terms of regulation and signal transduction.6, 7 Yet TRAIL had very specific effects on intracellular signaling in human adipocytes. We expected changes, such as downregulation of Akt expression and phosphorylation and activation of NF-κB.20 However, none of these typical effects induced by TNFα took place in adipocytes underlining the specific role of the TRAIL/TRAIL-R system in adipocytes.
Although TRAIL was upregulated in genetic obesity and acutely regulated by fasting/re-feeding in mice, there was no correlation of TRAIL expression with BMI in human adipose tissue. This could be either due to species-specific differences in TRAIL expression or due to the small sample size. As TRAIL is mainly expressed on immune cells and obesity is associated with macrophage infiltration into adipose tissue, we expected an increase of TRAIL with increasing BMI. However, our data suggest that another cellular source outside of adipose tissue is responsible for the obesity-related increase in circulating TRAIL.
Our results have several important implications. First, we propose that the TRAIL/TRAIL-R system might bear new molecular targets for the treatment of obesity-associated co-morbidities. This hypothesis is based on the finding that adipocyte-specific deletion of CD95 in mice resulted in improved glucose homeostasis, less triglyceride accumulation in the liver, and decreased systemic inflammation when fed with a high-fat diet.13 In line, an antibodies blocking TNFα (infliximab), which is typically used to treat patients with colitis ulcerosa or rheumatoid arthritis, was successfully used to inhibit high-fat diet-induced insulin resistance in mice.37 There is evidence that insulin sensitivity improves under infliximab therapy in patients who suffer from type 2 diabetes and simultaneous psoriatic arthritis.38 As TRAIL is increased with obesity, we wonder whether blocking TRAIL signaling would also prevent the development of insulin resistance. In vivo experiments are certainly needed to further test this hypothesis.
Second, the metabolic effects of TRAIL should be taken into account when TRAIL agonists are applied for the treatment of cancer. Besides a report on hepatotoxicity,39 no severe side effects of TRAIL therapy were published so far. TRAIL concentrations used in our in vitro study were 1–100 ng/ml. Physiological levels of circulating TRAIL may vary between 50 and 70 pg/ml.14 In Phase I studies, 0.5–15 mg/kg rhTRAIL were administered as 1 h intravenous infusion for 5 consecutive days, leading to a peak serum concentrations of almost 1000 μg/ml.40 Thus, adverse effects on glucose homeostasis should be considered, especially under long-term treatment and in overweight or obese patients.
Third, using TRAIL for cancer therapy might be a double-edged sword with our new discovery of TRAIL-induced cleavage of PPARγ leading to its inactivation. PPARγ has a major role in adipogenesis but is also expressed in several other cell types besides adipocytes, particularly in several cancer cells. Recently, the role of PPARγ in tumorigenesis has been critically discussed, it might exert anti- as well as pro-tumorigenic effects depending on the cell type or other conditions.41 The possibility of TRAIL-induced PPARγ cleavage with either positive or adverse consequences should be taken into account when treating cancer patients with recombinant TRAIL or agonistic TRAIL antibodies.We further propose that the newly identified pathway of TRAIL-induced PPARγ cleavage might have an impact on differentiation, maturation or activation processes in the immune system (summarized in Falschlehner et al.28). TRAIL is an important factor in T-cell homeostasis and differentiation, and PPARγ was implicated in the differentiation of naive T cells into their effector subsets (overview in Schmidt et al.41). The direct regulatory connection between TRAIL and PPARγ opens a completely new perspective, and this pathway should receive further attention regarding TRAIL-dependent immune processes.
In conclusion, we identified a novel metabolic function of the death ligand TRAIL. By binding to TRAIL-R2, TRAIL activates the cleavage of caspase-8 and -3, which in turn cleaves and inactivates PPARγ. This causes changes in gene expression of lipogenic genes and finally leads to the inhibition of insulin-stimulated glucose uptake and lipogenesis. Most interestingly, TRAIL-R2 and its mouse homolog DR5 are regulated upon acute and chronic energy imbalance in murine and human adipose. This leads us to conclude that the TRAIL/TRAIL-R system might bear new molecular targets for the treatment of obesity-associated co-morbidities. Furthermore, consideration should be given to this newly identified pathway with respect to possible negative side effects of TRAIL in cancer therapy and TRAIL-regulated processes in the immune system.
Materials and Methods
Materials
Cell culture media and supplements were from Invitrogen (Darmstadt, Germany). Recombinant human TRAIL was purchased from R&D Systems (Wiesbaden, Germany). The fully human agonist monoclonal antibodies against TRAIL-R1 and TRAIL-R2, mapatumumab and lexatumumab, respectively, were kind gifts from Human Genome Sciences.42 All other chemicals and reagents were obtained from Sigma Aldrich (München, Germany), if not otherwise stated.
Cell culture
Primary human preadipocytes isolated from adipose tissue and SGBS cells were cultured and their adipogenic differentiation was induced as described elsewhere.43, 44 After 14 days in adipogenic medium, cells were microscopically investigated and used for experiments if rate of adipogenic differentiation was >80%.
Basal and insulin-stimulated glucose uptake and de novo lipogenesis
After washing twice with PBS, SGBS adipocytes on day 14 of adipogenic were incubated in serum-free medium supplemented with rhTRAIL or PBS+0.05% BSA (vehicle) for 24 h. For glucose uptake, cells were stimulated with or without insulin (10 nM) for 15 min and 2-deoxy-D-[14C]-glucose (0.2 μCi/well) was added for another 15 min. For lipogenesis, insulin (10 nM) and D-[14C]-glucose (0.1 μCi/well) were added for 24 h. After incubation with glucose, cells were washed twice in ice-cold PBS, harvested with 100 mM NaOH, and incorporation of 2-deoxy-D-[14C]-glucose or D-[14C]-glucose was measured on a β-counter.
Western blot analysis
Western blot analysis was performed as described previously.36 Enhanced chemiluminescence was used for detection (Amersham Bioscience, Freiburg, Germany). All western blots shown are representative of at least three independent experiments. The following antibodies were used: rabbit anti-phospho Akt, mouse anti-phospho ERK, rabbit anti-panAkt, rabbit anti-phospho p38, mouse anti-p38, rabbit anti-phopsho JNK, rabbit anti-caspase-1, rabbit anti-caspase-3, rabbit anti-caspase-6, rabbit anti-FASN, rabbit anti-PPARγ (all from Cell Signaling, Beverly, MA, USA), JNK, mouse anti-caspase-8 (Alexis, Gruenberg, Germany), mouse anti-GLUT4 (R&D), rabbit anti-ERK (Sigma, München, Germany), and mouse anti-β-actin (Sigma). HRP-conjugated goat anti-mouse IgG and goat anti-rabbit IgG were obtained from Santa Cruz Biotechnology, Heidelberg, Germany.
Reverse transcription-PCR
Total RNA was prepared using RNeasy Lipid tissue kit (Qiagen, Hilden, Germany). cDNA synthesis was performed as described previously.36 qPCR on specific genes was carried out on a Roche lightcycler, Grenzach, Germany. The mRNA levels of genes were normalized to those of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) using ΔCt method. Fold changes of gene expression were calculated by the ΔΔCt method. Primer sequences are GAPDH forward 5′-GAAGGTGAAGGTCGGAGTC-3′, GAPDH reverse 5′-GAAGATGGTGATGGGATTTC-3′, Glut-4 forward 5′-TTCCAACAGATAGGCTCCGAAG-3′, Glut-4 reverse 5′-AAGCACCGCAGAGAACACAG-3′, ACC forward 5′-CTGTGGCTTCTCCAGCAGAATTTGTTACTC-3′, ACC reverse 5′-GATCTGCCATCTTAATGTATTCTGCATTGGCT-3′, FASN forward 5′-CTACCTGAGCATAGTGTGGAAGACGCTG-3′, FASN reverse 5′-CATCCCACTGGTACACCTTCCCACTCAC-3′. All primers were obtained from Thermo-Fischer Scientific (Ulm, Germany).
Surface expression of TRAIL receptors
To determine the surface receptor expression of TRAIL receptors, SGBS preadipocytes and adipocytes were incubated with mouse anti-human TRAIL-R1 and -R2 monoclonal antibody (all from ApoTech Corporation, Epalinges, Switzerland) for 30 min at 4 °C, washed in PBS containing 1% FCS, incubated with rabbit anti-mouse-F(ab’)2IgG/Biotin (BD Biosciences, Heidelberg, Germany) for 20 min at 4 °C in the dark, washed in PBS containing 1% FCS, incubated with streptavidin-PE (BD Biosciences) for 20 min at 4 °C in the dark, and analyzed by flow cytometry using CellQuest software.
Nuclear protein extraction and electrophoretic mobility shift assay
Adherent cells were collected from 10-cm dishes by scraping and centrifugation (10 000 × g for 5 min at 4 °C). After washing once with ice-cold PBS, cell pellets were resuspended in 200 μl low-salt buffer (10 mM HEPES-OH, pH 7.9; 1.5 mM MgCl2; 10 mM KCl) and incubated for 10 min on ice. After addition of 20 μl of a 10% Nonidet P-40 solution, samples were mixed vigorously for 30 s. Nuclei were collected by centrifugation and resuspended in 50 μl high-salt buffer (20 mM HEPES-OH, pH 7.9; 420−mM NaCl; 1.5 mM MgCl2; 0.2 mM EDTA; 25% glycerol). Both buffers were supplemented with a protease- inhibitor cocktail (Sigma); 0.2 mM PMSF; 0.5 mM dithiothreitol (DTT) and 1 mM sodium-ortho-vanadate before use. Nuclei were incubated 15 min on ice and vortexed periodically. Nuclear extracts were obtained by centrifugation at 12 500 g for 10 min at 4 °C and stored at −80 °C. Protein concentration was determined with the BCA protein assay kit (Pierce, Rockford, IL), according to the manufacturer’s instruction. Oligonucleotides were purchased from Biomers.net (Ulm, Germany): standard NF-κB probe: 5′-AGTTGAGGGGACTTTCCCAGGC-3′ (sense) and 5′-GCCTGGGAAAGTCCCCTCAACT-3′ (antisense); Single-stranded oligonucleotides were labeled with γ-32P-ATP (Amersham, Freiburg, Germany) by T4-polynucleotide kinase (MBI Fermentas, St. Leon-Rot, Germany). A twofold molar excess of unlabeled complementary oligonucleotides was annealed, and double-stranded oligonucleotides were purified on spin columns (Micro Bio-Spin P30; Bio-Rad, Munich, Germany). Binding reactions were performed for 30 min on ice in 20 μl buffer (1 mM MgCl2, 0.5 mM EDTA, 0.5 mM DTT, 50 mM NaCl, 10 mM Tris-HCl, pH 7.5; 4% glycerol) containing 5 μg nuclear extract protein, 1 μg poly(dI/dC; Sigma), and 10 000 cpm-labeled oligonucleotide.
Immunofluorescence
For immunofluorescence, SGBS cells were grown and differentiated on chamberslides (BD Falcon, Heidelberg, Germany). At day 14 of adipogenic differentiation, cells were treated with vehicle or 100 ng/ml TRAIL. After 6 h, monolayers were washed, and fixed with 4% PFA followed by permeabilization with 0.5% Triton-X 100 in PBS, blocked with 10% FCS in PBS and incubated overnight at 4 °C with first antibody. Next, cells were washed, incubated for 1 h with secondary fluorescence-conjugated antibodies. Finally, cells were washed again and mounted with DAKO fluorescence mounting medium (DAKO, Hamburg, Germany) containing DAPI (0.5 μg/ml, Invitrogen). Following antibodies were used: mouse anti-PPARγ (Santa Cruz), rabbit anit-p65 (Santa Cruz), rabbit anti-active caspase-3 (Cell Signaling), goat anti-mouse IgG1-PE (BD Pharmingen, Heidelberg, Germany), and goat anti-rabbit IgG-FITC (Santa Cruz). Slides were viewed using an Olympus AX70 fluorescence microscope equipped with appropriate filters (Olympus Optical Co., Hamburg, Germany). Images were captured with a CC12 digital camera (Soft Imaging System, Münster, Germany) using cell^P software (Olympus, Hamburg, Germany).
Lentiviral-mediated knockdown of caspase-3 and caspase-8
Knock down of caspase-3 and caspase-8 was performed by using the BLOCK-iT Inducible H1 Lentiviral RNAi System (Invitrogen, Darmstadt, Germany) according to the manufacturer’s protocol. Specific shRNA constructs against caspase-3 #1 (start 834, 5′-GCCGACTTCTTGTATGCATAC-3′), caspase-3 #2 (start 917, 5′-GCTTTGTGCCATGCTGAAACA-3′), caspase-8 #1 (start 411, 5′-GGAACAACTGGACAGTGAAGA-3′), caspase-8 #2 (start 693, 5′-GGGTCATGCTCTATCAGATTT-3′), and a sequence with no corresponding counterpart in the human genome (hyper random sequence) (5′-GATCATGTAGATACGCTCA-3′) were cloned into the pENTR/H1/TO vector using the BLOCK-iT Inducible H1 RNAi Entry vector kit. After propagation of the vector, the shRNA expression cassettes were transferred into the pLenti4/BLOCK-iT-DEST vector by targeted recombination. Viral particles were produced after transfection of 293FT cells in the presence of Lipofectamine2000 using the ViraPower Lentiviral Expression Systems (Invitrogen). Three days after transfection, viral supernatants were harvested, filtered through a 0.45-μm PVDF filter and stored at −80 °C upon usage. SGBS adipocytes were transduced at day 10 and used for experiments on day 14 of adipogenic differentiation. The knock down was controlled by western blot analysis. For all experiments, cells from at least three independent lentiviral transductions were used.
Experimental subjects
Human fat tissue samples were obtained from 10 Caucasian obese and lean women undergoing plastic surgery. Mean age of patients was 49 years (range 40–64 years) and their mean BMI was 39 kg/m2 (range 20–67 kg/m2). All procedures were performed in accordance with the Declaration of Helsinki guidelines and approved by the ethics committee of the University of Ulm. Patients gave a written informed consent in advance.
Experimental animals
Data for obese mice were obtained from Keller et al.17 Male FVB/NJ mice at the age of 12 weeks were killed in the fed state at 1900 hours (n=5; standard diet, no. 5058; LabDiet, Richmont, IN, USA), 24 h fasted at 1900 hours (n=5) or 2 h after re-feeding at 2100 hours following a 24-h fast. The animals were housed in a controlled environment (20±2 °C, 12 h/12 h of a light/dark cycle) and had free access to water. All animal experiments were approved by the Institutional Animal Care and Use Committee of the University of Texas Southwestern Medical Center (Dallas, TX, USA). Body weights were taken before and after fasting. Inguinal as well as epididymal adipose tissue were dissected and stored in RNAlater (Invitrogen) at −80 °C before RNA extraction. mRNA extraction and qRT-PCR analysis was performed as described previously.36
Statistical analysis
If not otherwise stated, data from three independent triplicate experiments were expressed as mean and S.E.M. For statistical comparison, one-way ANOVA (Holm–Sidak–Method treatment versus control) was performed. P<0.05 was considered statistically significant. For correlation analysis, Spearman’s correlation was performed. All statistical tests were performed using SigmaStat version 11.0 (Systat Software GmbH, Inc., San Jose, CA, USA).
Abbreviations
- ACC:
-
acetyl CoA-carboxylase
- BMI:
-
body mass index
- DISC:
-
death-inducing signaling complex
- FASN:
-
fatty acid synthase
- Glut-4:
-
glucose transporter-4
- JNK:
-
c-Jun N-terminal kinase
- MAPK:
-
mitogen-activated protein kinase
- NF-κB:
-
nuclear factor kappa B
- PPARγ:
-
peroxisome proliferator-activated receptor gamma
- SGBS:
-
Simpsom Golabi Behmel syndrome
- TNF:
-
tumor necrosis factor
- TRAIL:
-
TNF-related apoptosis-inducing ligand
- TRAIL-R:
-
TNF-related apoptosis-inducing ligand receptor
- WAT:
-
white adipose tissue
References
Fischer-Posovszky P, Wabitsch M, Hochberg Z . Endocrinology of adipose tissue-an update. Horm Metab Res 2007; 39: 314–321.
Scherer PE . Adipose tissue: from lipid storage compartment to endocrine organ. Diabetes 2006; 55: 1537–1545.
Trayhurn P . Endocrine and signalling role of adipose tissue: new perspectives on fat. Acta Physiol Scand 2005; 184: 285–293.
Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW . Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest 2003; 112: 1796–1808.
Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest 2003; 112: 1821–1830.
Wiley SR, Schooley K, Smolak PJ, Din WS, Huang CP, Nicholl JK et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity 1995; 3: 673–682.
Gonzalvez F, Ashkenazi A . New insights into apoptosis signaling by Apo2L/TRAIL. Oncogene 2010; 29: 4752–4765.
Koschny R, Walczak H, Ganten TM . The promise of TRAIL—potential and risks of a novel anticancer therapy. J Mol Med (Berl) 2007; 85: 923–935.
Herbst RS, Eckhardt SG, Kurzrock R, Ebbinghaus S, O’Dwyer PJ, Gordon MS et al. Phase I dose-escalation study of recombinant human Apo2L/TRAIL, a dual proapoptotic receptor agonist, in patients with advanced cancer. J Clin Oncol 2010; 28: 2839–2846.
Zhang XD, Nguyen T, Thomas WD, Sanders JE, Hersey P . Mechanisms of resistance of normal cells to TRAIL induced apoptosis vary between different cell types. FEBS Lett 2000; 482: 193–199.
Hotamisligil GS . The role of TNFalpha and TNF receptors in obesity and insulin resistance. J Intern Med 1999; 245: 621–625.
Hotamisligil GS, Shargill NS, Spiegelman BM . Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science 1993; 259: 87–91.
Wueest S, Rapold RA, Schumann DM, Rytka JM, Schildknecht A, Nov O et al. Deletion of Fas in adipocytes relieves adipose tissue inflammation and hepatic manifestations of obesity in mice. J Clin Invest 2010; 120: 191–202.
Choi JW, Song JS, Pai SH . Associations of serum TRAIL concentrations, anthropometric variables, and serum lipid parameters in healthy adults. Ann Clin Lab Sci 2004 Autumn 34: 400–404.
Fischer-Posovszky P, Tornqvist H, Debatin KM, Wabitsch M . Inhibition of death-receptor mediated apoptosis in human adipocytes by the insulin-like growth factor I (IGF-I)/IGF-I receptor autocrine circuit. Endocrinology 2004; 145: 1849–1859.
Mader I, Wabitsch M, Debatin KM, Fischer-Posovszky P, Fulda S . Identification of a novel proapoptotic function of resveratrol in fat cells: SIRT1-independent sensitization to TRAIL-induced apoptosis. FASEB J 2010; 24: 1997–2009.
Keller MP, Choi Y, Wang P, Davis DB, Rabaglia ME, Oler AT et al. A gene expression network model of type 2 diabetes links cell cycle regulation in islets with diabetes susceptibility. Genome Res 2008; 18: 706–716.
Wakelee HA, Patnaik A, Sikic BI, Mita M, Fox NL, Miceli R et al. Phase I and pharmacokinetic study of lexatumumab (HGS-ETR2) given every 2 weeks in patients with advanced solid tumors. Ann Oncol 2010; 21: 376–381.
Wajant H, Scheurich P . TNFR1-induced activation of the classical NF-kappaB pathway. Febs J 2011; 278: 862–876.
Ruan H, Hacohen N, Golub TR, Van Parijs L, Lodish HF . Tumor necrosis factor-alpha suppresses adipocyte-specific genes and activates expression of preadipocyte genes in 3T3-L1 adipocytes: nuclear factor-kappaB activation by TNF-alpha is obligatory. Diabetes 2002; 51: 1319–1336.
Liu LS, Spelleken M, Rohrig K, Hauner H, Eckel J . Tumor necrosis factor-alpha acutely inhibits insulin signaling in human adipocytes: implication of the p80 tumor necrosis factor receptor. Diabetes 1998; 47: 515–522.
Medina EA, Afsari RR, Ravid T, Castillo SS, Erickson KL, Goldkorn T . Tumor necrosis factor-{alpha} decreases Akt protein levels in 3T3-L1 adipocytes via the caspase-dependent ubiquitination of Akt. Endocrinology 2005; 146: 2726–2735.
Wueest S, Rapold RA, Schoenle EJ, Konrad D . Fas activation in adipocytes impairs insulin-stimulated glucose uptake by reducing Akt. FEBS Lett 2010; 584: 4187–4192.
Fernandez-Veledo S, Vila-Bedmar R, Nieto-Vazquez I, Lorenzo M . c-Jun N-terminal kinase 1/2 activation by tumor necrosis factor-alpha induces insulin resistance in human visceral but not subcutaneous adipocytes: reversal by liver X receptor agonists. J Clin Endocrinol Metab 2009; 94: 3583–3593.
He F, Doucet JA, Stephens JM . Caspase-mediated degradation of PPARgamma proteins in adipocytes. Obesity (Silver Spring) 2008; 16: 1735–1741.
Guilherme A, Tesz GJ, Guntur KV, Czech MP . Tumor necrosis factor-alpha induces caspase-mediated cleavage of peroxisome proliferator-activated receptor gamma in adipocytes. J Biol Chem 2009; 284: 17082–17091.
Stienstra R, Joosten LA, Koenen T, van Tits B, van Diepen JA, van den Berg SA et al. The inflammasome-mediated caspase-1 activation controls adipocyte differentiation and insulin sensitivity. Cell Metab 2010; 12: 593–605.
Falschlehner C, Schaefer U, Walczak H . Following TRAIL’s path in the immune system. Immunology 2009; 127: 145–154.
Wang S . TRAIL: a sword for killing tumors. Curr Med Chem 2010; 17: 3309–3317.
Secchiero P, Gonelli A, Mirandola P, Melloni E, Zamai L, Celeghini C et al. Tumor necrosis factor-related apoptosis-inducing ligand induces monocytic maturation of leukemic and normal myeloid precursors through a caspase-dependent pathway. Blood 2002; 100: 2421–2429.
Secchiero P, Zerbinati C, Rimondi E, Corallini F, Milani D, Grill V et al. TRAIL promotes the survival, migration and proliferation of vascular smooth muscle cells. Cell Mol Life Sci 2004; 61: 1965–1974.
Rimondi E, Secchiero P, Quaroni A, Zerbinati C, Capitani S, Zauli G . Involvement of TRAIL/TRAIL-receptors in human intestinal cell differentiation. J Cell Physiol 2006; 206: 647–654.
Lamhamedi-Cherradi SE, Zheng S, Tisch RM, Chen YH . Critical roles of tumor necrosis factor-related apoptosis-inducing ligand in type 1 diabetes. Diabetes 2003; 52: 2274–2278.
Mi QS, Ly D, Lamhamedi-Cherradi SE, Salojin KV, Zhou L, Grattan M et al. Blockade of tumor necrosis factor-related apoptosis-inducing ligand exacerbates type 1 diabetes in NOD mice. Diabetes 2003; 52: 1967–1975.
Biolo G, Secchiero P, De Giorgi S, Tisato V, Zauli G . The energy balance positively regulates the levels of circulating TNF-related apoptosis inducing ligand in humans. Clin Nutr 2012; 31: 1018–1021.
Fischer-Posovszky P, Keuper M, Nagel S, Hesse D, Schurmann A, Debatin KM et al. Downregulation of FLIP by cycloheximide sensitizes human fat cells to CD95-induced apoptosis. Exp Cell Res 2011; 317: 2200–2209.
Araujo EP, De Souza CT, Ueno M, Cintra DE, Bertolo MB, Carvalheira JB et al. Infliximab restores glucose homeostasis in an animal model of diet-induced obesity and diabetes. Endocrinology 2007; 148: 5991–5997.
Yazdani-Biuki B, Stelzl H, Brezinschek HP, Hermann J, Mueller T, Krippl P et al. Improvement of insulin sensitivity in insulin resistant subjects during prolonged treatment with the anti-TNF-alpha antibody infliximab. Eur J Clin Invest 2004; 34: 641–642.
Zheng SJ, Wang P, Tsabary G, Chen YH . Critical roles of TRAIL in hepatic cell death and hepatic inflammation. J Clin Invest 2004; 113: 58–64.
Ashkenazi A, Holland P, Eckhardt SG . Ligand-based targeting of apoptosis in cancer: the potential of recombinant human apoptosis ligand 2/Tumor necrosis factor-related apoptosis-inducing ligand (rhApo2L/TRAIL). J Clin Oncol 2008; 26: 3621–3630.
Schmidt MV, Brune B, von Knethen A . The nuclear hormone receptor PPARgamma as a therapeutic target in major diseases. ScientificWorldJournal 2010; 10: 2181–2197.
Humphreys RC, Halpern W . Trail receptors: targets for cancer therapy. Adv Exp Med Biol 2008; 615: 127–158.
Fischer-Posovszky P, Newell FS, Wabitsch M, Tornqvist HE . Human SGBS Cells—a Unique Tool for Studies of Human Fat Cell Biology. Obes Facts 2008; 1: 184–189.
Wabitsch M, Brenner RE, Melzner I, Braun M, Moller P, Heinze E et al. Characterization of a human preadipocyte cell strain with high capacity for adipose differentiation. Int J Obes Relat Metab Disord 2001; 25: 8–15.
Acknowledgements
We thank Alexandra Killian and Susann Baumgart for excellent technical assistance. PFP is funded by a Margarete von Wrangell scholarship financed by the Baden-Wuerttemberg Ministry of Science, Research and Arts, the European Social Fund, and Ulm University. This study was partly supported by the German Research Association (FI 1700/1-1, WA 1096/3–2).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by M Federici
Supplementary Information accompanies the paper on Cell Death and Disease website
Supplementary information
Rights and permissions
This work is licensed under the Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Keuper, M., Wernstedt Asterholm, I., Scherer, P. et al. TRAIL (TNF-related apoptosis-inducing ligand) regulates adipocyte metabolism by caspase-mediated cleavage of PPARgamma. Cell Death Dis 4, e474 (2013). https://doi.org/10.1038/cddis.2012.212
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cddis.2012.212
Keywords
This article is cited by
-
20 Years with SGBS cells - a versatile in vitro model of human adipocyte biology
International Journal of Obesity (2022)
-
TRAIL/DR5 pathway promotes AKT phosphorylation, skeletal muscle differentiation, and glucose uptake
Cell Death & Disease (2021)
-
The role of caspase-8 in the tumor microenvironment of ovarian cancer
Cancer and Metastasis Reviews (2021)
-
Developing TRAIL/TRAIL death receptor-based cancer therapies
Cancer and Metastasis Reviews (2018)
-
Trail (TNF-related apoptosis-inducing ligand) induces an inflammatory response in human adipocytes
Scientific Reports (2017)
