
Abstract
Multiple growth factors (e.g., BMP2, TGF-β1, FGF2) and isolated genes have been shown to improve osteoblastic proliferation and mineralization, advancing bone tissue engineering. Among these factors, both polydopamine (PDA) and dopamine (DA) monomer have recently been reported to increase osteoblast proliferation and mineralization in vitro. Although a well-characterized neurotransmitter, DA’s role in the bone is unknown. We hypothesize that DA can directly act on osteoblasts, and examined whether osteoblasts express DA receptors that respond to exogenous DA. mRNAs and protein cell lysates were obtained from MC3T3-E1 cells during osteogenic differentiation phase. Reverse transcription polymerase chain reaction and western blot analysis were used to examine the expression of DA receptors, D1–D5. Dose-response effect and time course of DA treatment on cell proliferation, mineralization, and osteogenic differentiation were investigated at pre-determined days. Real-time PCR was performed to investigate whether DA affects osteogenic gene expression (ALP, BSP, OC, OSX, RUNX2, and Collagen1a2) with or without receptor antagonists (SCH233390 and GR103691). Two-way ANOVA was used for statistical analysis. All five DA receptors (D1, D2, D3, D4, and D5) mRNAs and proteins were expressed in MC3T3-E1 cells. DA treatment increased cell proliferation for up to 7 days (P < 0.05). Osteogenic mineralization was significantly greater in the DA-treated group than control group (P < 0.05). Finally, expression of all the osteogenic genes was inhibited by DA receptor antagonists for D1, D3, and D5. Our findings suggest that MC3T3-E1 osteoblasts express functional DA receptors that enhance proliferation and mineralization. PDA is not biologically inert and has important implications in orthopedic applications. Furthermore, osteoblast differentiation might be regulated by the nervous system, presumably during bone development, remodeling, or repair.
Similar content being viewed by others
Introduction
Three-dimensional (3D) scaffolds for tissue regeneration require mechanical strength and the desired environment for seeded cells to gain functionality in the scaffold materials. Scaffolds that release soluble bioactive factors achieved higher success rates of tissue regeneration compared to direct delivery of signal molecules into the regeneration site or on a porous scaffold.1 Oxidative polymerization of dopamine (DA) was recently used in our laboratory to increase the mechanical property of 3D bone scaffolds made of Hydroxyapatite-Gelatin Calcium Silicate (HGCS) by providing an interpenetrating polydopamine (PDA) network (HGCS-PDA).2
Surface modification with PDA, inspired by marine bivalve mollusks and mussels, has been used to promote cell adhesion in wet environments and on surfaces resistant to cell adhesion.3 Under alkaline condition, the hydroxyl or C=O groups in 1,2-dihydroxybenzene oxidize to Quinone and induce polymerization of the DA. This reaction has been applied to form a thin layer coating the substrates by covalent bond, hydrogen bond, and metal chelation.4
The adhesion of mouse pre-osteoblasts MC3T3-E1 cells were significantly improved on the surface of PDA modified materials such as polyethylene, polytetrafluoroethylene, silicone, and polydimethylsiloxane.5–7 Additionally, PDA coating on substrates such as titanium and electrospun polymers has been shown to promote osteogenic differentiation.8,9 While these exciting results have improved cell adhesion, they are restricted to two-dimensional surface coating.
We were the first to report the amalgamation of PDA in a 3D composite structure instead of a surface coating. The use of PDA in our HGCS-PDA substrate increased the mechanical strength by 30% compared to scaffolds with no DA. Surprisingly, the HGCS-PDA scaffold was found to release DA into the surrounding liquid environment, which was detected by using high-performance liquid chromatography (HPLC).2 In general, DA is known to function as a neurotransmitter in neurons by binding to D1 and D2 types of DA receptors. While signals from D1-type receptors transduce through G proteins to activate adenylyl cyclase, forming cyclic adenosine monophosphate (cAMP) and activating protein kinase A (PKA), D2-type receptors block this signaling by inhibiting adenylyl cyclase.10
Recent data suggest that osteoblasts might respond to neurotransmitters. For instance, sensory and sympathetic nerve fibers directly transduce chemical messenger to the bone and periosteum.11 Also, the finding of nerve endings directly contacted with bone cells may have possible effect on the bone remodeling. Axons containing catecholamine were found near osteoblasts in vivo.12 The presence of beta adrenergic receptors in both osteoblasts and osteoclasts further support the existence of receptors for neurotransmitters and neuropeptide from sensory and sympathetic nerve fibers.13 Finally, many studies showed that damaged or missing sympathetic nerves result in abnormal bone formation and bone mass.14 Therefore, these findings implicate that osteoblasts may respond to neurotransmitters such as DA.
In addition to the DA effects on osteoblasts, there were relevant evidences of DA receptor and DA active transporter (DAT) can also influence osteogenic activity. Hanami et al. have demonstrated that D2-like DA receptor signaling suppressed human osteoclastogenesis.15 Furthermore, Bliziotes et al. have shown mice deleted for DAT gene exhibit reduced bone mass.16 These findings indicate that dopaminergic signaling plays an important role in bone homeostasis via direct effects upon osteoclast differentiation and the deletion of the DAT gene results in deficiencies in skeletal structure and integrity.
To determine whether DA can influence osteoblast proliferation and differentiation, we hypothesize osteoblasts may express DA receptors and respond to the DA. To test the hypothesis, MC3T3-E1 osteoblast cells were examined for DA receptor expression using RT-PCR and western blot analysis. Potential effects of the DA on osteogenic gene expression, proliferation, and mineralization were also investigated.
Materials and methods
Osteoblasts cultures
MC3T3-E1 pre-osteoblasts were obtained from ATCC (Subclone 14, CRL-2594).17 The cells were cultured and expanded in growth media (alpha minimal essential medium (α-MEM) containing 10% fetal bovine serum (FBS) and 1% penicillin and streptomycin) and were differentiated with growth media supplemented with 10 mmol·L−1 beta glycerophosphate and 0.2 mmol·L−1 ascorbic acid at 37°C under 5% CO2. The media was changed every 3 days.
RT-PCR for DA receptor expression
Total RNA was isolated from 5 × 106 cells by following the instructions from QiagenRNeasy Mini kits (Qiagen, Valencia, CA, USA), and then the RNA was reverse-transcribed into cDNA using an QuantiTect Reverse Transcription Kit (Qiagen, Valencia, CA, USA). Oligonucleotide primers for the PCR were designed for mouse DA receptors as described in Table 1. For the first step of the PCR, the conditions for the DA receptors and GAPDH were 29 cycles of denaturation (at 94°C for 40 seconds), annealing (at 55°C for 45 seconds), and extension (at 72°C for 40 seconds), followed by a final 5-minute extension at 72°C. RNAs extracted from fresh mouse brain were used as a control to identify the correct size of DA mRNAs from MC3T3-E1 cells. To reduce non-specific binding in PCR products, nested PCR was performed. A 1 µL of amplified PCR products (D1–D5 and GAPDH) from the first step was used as a template to initiate another PCR reaction, which was performed under the same conditions as first step except that different primers were used for the nested PCR (Table 1). The PCR products from both first and second steps were separated by electrophoresis through a 1% agarose gel containing GelRed Nucleic Acid Stain (Biotium, Inc., Hayward, CA, USA), and the image was captured by ImageQuant LAS 4000 (GE, Piscataway, NJ, USA).
Western blot analysis for DA receptors
MC3T3-E1 cells under osteogenic differentiation for 0, 4, 7, and 14 days were lysed with RIPA lysis buffer (50 mmol·L−1 Tris-HCl: pH 8.0, 5 mmol·L−1 EDTA, 150 mmol·L−1 NaCl, 1% Triton X-100, 1 mmol·L−1 phenylmethylsulfonyl fluoride, and phosphatase & protease inhibitor cocktail) for 10 minutes. After centrifugation at 14 000 rpm·min−1, supernatant was collected as a whole-cell lysate.
Mouse brain tissue was used as positive control. All animal works were performed with Institutional Animal Care and Use Committee (IACUC) approval at Duke University. Total protein was measured by using Pierce BCA Protein Assay Kit (Thermo Fisher Scientific Inc., Rockford, IL, USA). Cell lysate from each group containing 15 μg of protein was loaded to and separated through 12% NuPAGE SDS-PAGE Gel (Invitrogen, Carlsbad, CA, USA), and then transferred onto a nitrocellulose membrane (Millipore, Billerica, MA, USA) by using Trans-Blot SD Semi-Dry Transfer Cell (Bio-Rad, Hercules, CA, USA). After blocking for 30 minutes, the membrane was incubated overnight at 4°C with the anti-DA receptors 1 (Abcam, Cambridge, MA, USA), 2 (Alomone Labs, Jerusalem, Israel), 3 (Alomone Labs, Jerusalem, Israel), 4 and 5 (Abcam®, Cambridge, MA, USA) antibodies (1:500 ratio). After subsequent washing, the membrane was incubated for 1 hour with secondary antibodies conjugated to HRP (Milipore, Billerica, MA, USA), rewashed, and developed using an enhanced chemiluminescence solution (Thermo Fisher Scientific Inc., Rockford, IL, USA). The band images were captured by ImageQuant LAS 4000 (GE, Piscataway, NJ, USA). Beta actin (Abcam®, Cambridge, MA, USA) was used as a loading control and for normalization.
Optimization of DA concentration
MC3T3-E1 cells (2 × 105 per 35 mm dish) were maintained in the growth media for 24 hour. DA was dissolved in molecular grade ultrapure water (Mediatech, Inc., Manassas, VA, USA) and filter through 0.2 µmol·L−1 pore syringe filters. The cells were then exposed to DA in different final concentrations (0, 50, 100, and 200 µmol·L−1) for 24, 36, 48, and 72 hours, respectively.
The dose and time effect of DA on MC3T3-E1 cells were determined with 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfophenyl)-2H-tetrazolium salt (MTS) assay according to the manufacturer’s instructions (CellTiter96; Promega, Madison, WI, USA). Briefly, 40 µL of MTS reagent in 500 µL of basal media was added to the cells in each dish at the end of the designed cultivation period and incubated for 1 hour at 37°C. Aliquots of 100 µL were removed for analysis. Colored formazan products were quantified by measuring absorbance at 490 nm using a microplate reader (Bio-Rad Model 550, Life Science Research, Hercules, CA, USA). Three samples were tested with triplicate at each time point for each group.
Proliferation assay
MC3T3-E1 cells (5 × 104 cells per 35 mm dish) were seeded and attached overnight, and then stimulated with DA. The cells were supplied with fresh culture media containing 50 µmol·L−1 of DA every 48 hours during the culture period. Cell proliferation also was carried out under low FBS (3%) containing growth media. After 1, 3, 5, and 7 days in culture, the absorbance of MTS solution from each well was measured at 490 nm using a microplate reader. For those proliferating cells, 5-bromo-2′-deoxyuridine (BrdU: Life Technologies, Grand Island, NY, USA) was incorporated with cells for 2 hours and detected using anti-BrdU antibody as instructed by company protocol.
Mineralization assay with cetylpyridiniumchloride quantitation
MC3T3-E1 cells were seeded (2 × 105 cells per 35 mm dish) and supplied with fresh osteogenic media every 3 days. A total of four groups were examined for mineral formation: the cells cultured with osteogenic media, osteogenic media containing 50 µmol·L−1 DA, growth media containing 50 µmol·L−1 DA, and growth media only. A control group without cells was also examined. After 7, 14, 18, and 21 days the cells were fixed with 75% cold ethanol for 30 minutes and then stained with 1% Alizarin Red (Acros Organics, Geel, Belgium) solution (pH 4.2) for 10 minutes at RT. The cells were rinsed with dH2O six times and air dried. Color images of the mineral deposition were acquired using an Olympus DP12 camera (Olympus Optical Co. Ltd, Japan) attached to a Nikon Eclipse Ti-U microscope (Nikon Instruments, Melville, NY, USA). To quantify Alizarin Red S-retained minerals, 10% (v/w) cetylpyridinium chloride (CPC; Sigma-Aldrich, St. Louis, MO, USA) in sodium phosphate was used and measured at 570 nm.
Real-time PCR for osteogenic gene expression with DA receptor blockers
MC3T3-E1 cells were culture with both growth and osteogenic media for 0, 4, and 7 days. DA receptor antagonists SCH233390 (Tocris Bioscience, Bristol, UK) for D1 and D5 and GR103691 (Tocris Bioscience, Bristol, UK) for D3 was treated in each 35 mm dish with final concentration of 1 mmol·L−1 and 0.5 mmol·L−1, respectively. After 30 minutes of antagonists treatment, DA solution was added directly to the experimental group for a final concentration of 50 µmol·L−1. cDNA was synthesized as previously described in RT-PCR. Real-time PCR was performed using the 7200 Fast Real-Time PCR System (Applied Biosystems, Bedford, MA, USA) to determine mRNA expression of each osteogenic specific genes (Table 2) after DA treatment. The control was a non-DA-treated culture.
Statistics
All data were presented in terms of mean ± SD and P value less than 0.05 was considered significant. While comparison of two groups was made by paired t-test, comparison within groups was made by ANOVA.
Results
Expression of DA receptor genes in MC3T3-E1 cells
Nested polymerase chain reaction (Nested PCR; primers in Table 1), showed clear nested PCR bands of all five DA receptors without any nonspecific bands (Figure 1). The size of PCR products from both mouse brain and MC3T3-E1 cells were found to be identical.18

Expression of dopamine (DA) receptor genes in cultured MC3T3‐E1 cells. Total RNAs extracted from MC3T3‐E1 cells were analyzed by RT‐PCR using specific primers for each DA receptor (Table 1). The size of the PCR products was analyzed by electrophoresis compared with mRNAs extracted from mouse brain (Br) tissue. D1, D2, D3, D4, and D5 receptors were consistently expressed in both MC3T3‐E1 cells and Br tissue. bp, base pair.
Western blot analysis of DA receptors in MC3T3-E1 cells
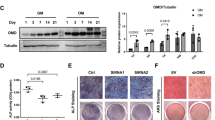
The band size of DA receptors of mouse brain and the MC3T3-E1 cells was almost identical, and the band specificity against antibodies was also high in both groups (Figure 2). Antibodies against D1, D2, D3, D4, and D5 receptor proteins recognized band size around 70, 50, 40, 70, and 50 kD respectively in both mouse brain and MC3T3-E cells. Although all five DA receptor expressions were clearly detected on day 0, day 4, day 7, and day 14, there is not much changes in the level of expression over time.

Western blot analysis of DA receptor expression on the MC3T3‐E1 osteoblasts during osteogenic differentiation on days 0, 4, 7, and 14. Br was used as control to identify the correct size of each DA receptor protein and binding specificity against each DA receptor antibody. Each loading was normalized by beta actin and quantified using Image J software. The relative molecular mass of significant band identified on the blots was labelled on the right side of blot image.
Optimization of DA treatment
DA concentration was optimized not only to prevent cell death due to overdose but also for a stimulatory effect on MC3T3-E1 cells. This was accomplished by investigating the effect of various DA concentrations as well as length of DA treatment. As illustrated in Figure 3, the highest OD values for the cells that were exposed at each time and dose are 0.32 ± 0.04 in 24 hours at the concentration of 100 µmol·L−1, 0.46 ± 0.02 in 36 hours with 50 µmol·L−1, 0.51 ± 0.02 in 48 hours with 50 µmol·L−1, and 0.66 ± 0.02 with 0 µmol·L−1 in 72 hours (data not shown here). Collectively, the highest OD value (0.51 ± 0.02) was acquired for the cells treated with 50 µmol·L−1 of DA for 48 hours, which is considered as the most effective condition for studies less than 72 hours. After the treatment of cells with 100 and 200 µmol·L−1 of DA for 36 and 48 hours, we could observe color darkening in culture media. Most of the cells that were treated with higher than 100 µmol·L−1 of DA died. Thus, the 50 µmol·L−1 of DA was used for experiments on cell proliferation up to 7 days of culture described below.

Optimization of DA concentration and treatment duration on MC3T3‐E1 cells using MTS assay. Total of 50 000 cells per 35 mm dish were seeded overnight and exposed to various concentrations (0, 50, 100, and 200 µmol·L−1) of dopamine for 24, 36, and 48 hours of culture respectively. Cell proliferation was most effective with 50 µmol·L−1 of DA treatment after 48 hours (n = 3 measurements from each three independent samples per group, *P < 0.05 vs. 0 µmol·L−1 dopamine group).
Effect of DA on MC3T3-E1 cell proliferation
Based on the DA optimization data, MC3T3-E1 cells were supplied with fresh media (10% FBS) containing 50 µmol·L−1 DA every 48 hours for 7 days. A significant difference in growth was observed up to day 4 between the DA-treated and non-treated cells (Figure 4a). The cell number in both groups increased insignificantly at almost the same rate up to day 7. DA treatment provided higher growth in the treated group than in the non-treated group only from day 1 to day 4. The value of OD measurement in the DA-treated group (0.76 ± 0.05) on day 3 was significantly higher than that of the non-treated group (0.65 ± 0.02, P < 0.05). However, MC3T3-E1 cells in both DA-treated and non-treated media represented almost the same growth rate from day 4 to day 7 without significant difference in the proliferation rate (P > 0.05). Beginning with cell density of 50 000 the confluent after 7 days of culture reached the plateau of the growth curve in both groups. Compared with the control cells lacking DA treatment, the MC3T3-E1 cells displayed that the enhanced proliferation by the DA (50 µmol·L−1) treatment up to 7 days. In addition, BrdU assay indicated higher number of BrdU+ cells in DA-treated group on day 1 but will have similar number of BrdU+ cells on days 3, 5, and 7 (Figure 4c).

Effect of DA on cell proliferation. MTS proliferation assays were performed on days 1, 3, 5, and 7 in growth media containing 10% FBS (a) and 3% FBS (b), respectively. Proliferating cells were detected by BrdU assay (c), scale: 100 μm. Cells were treated with 50 µmol·L−1 DA on day 0. The medium was changed every 48 hours with freshly prepared DA solution. The assay was performed in 96‐well plates with n = 3 measurements from each three independent samples per group.
MC3T3-E1 cells were supplied with media (3% FBS) containing 50 µmol·L−1 DA every 48 hours for 7 days. Significant differences in DA-treated cells on both days 3 and 5 were observed compared to non-treated cells. Although the overall cell growth in 3% serum was slower than in 10% serum, DA treatment on the cells clearly showed more effective proliferation than the cells without DA in 3% serum condition (Figure 4b).
Effect of DA on mineralization
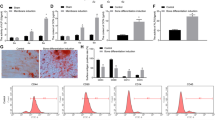
Bone mineralization resulting from calcium deposition is known as a late marker in osteogenic differentiation. Most mineralization was initiated after 7 days, the results on days 10, 14, 18, and 21 (Figure 5). From the scanned image of mineralization (Figure 5a), the DA-treated group (50 µmol·L−1) showed higher levels of mineralization as shown by a darker color, rougher particles and widely branched deposits in comparison with that in the non-treated group. The difference in mineralization was most significant after 18 days between the two groups. After 21 days, difference in mineralization could not be assessed due to over-deposition of minerals. The negative control group without DA showed no mineral deposits even after 21 days. The mineralization was induced using DA treated on the cells under non-osteogenic media. Although no distinct mineral deposits were observed until 14 days, only small, weakly stained mineral particles were observed 21 days, which was different than the mineral deposits in the other two groups under osteogenic media. The shape of the mineral nodules after DA treatment indicated the larger average size of nodules (>1 mm in diameter) and more branched with neighboring nodules than those in the group without DA treatment.

Mineral nodule formation after Alizarin Red S staining and quantification. (a) DA treatment enhanced mineralization by MC3T3‐E1 cells cultured with osteogenic media. The level of mineralization was equalized in both DA‐treated (50 µmol·L−1) and non‐treated group after 21 days. DA effect was also observed in the cells cultured with growth media for 21 days forming small minerals. Scale bar represents 200 μm. (b) Semi‐quantitative analysis of Alizarin Red S‐stained particles by CPC extraction method. Extracted solution was measured by the absorbance at 570 nm (n = 6, *P < 0.05 vs. in osteogenic media without DA, and †P < 0.05 vs. in growth media without DA). (c) Image J analysis of the mineral coverage of the culture dish. Both CPC and Image J analysis yielded similar results.
Both CPC (Figure 5b) and Image J analysis (Figure 5c) extraction indicated an increased area of calcium nodules in the DA-treated group compared with the non-treated group on days 10, 14, 18, and 21. Mineralization was significantly higher at the 10th, 14th, and 18th days for the DA-treated group, compared to the non-treated group. However, the level of mineralization was just little higher on the 21st day of differentiation with DA.
Effect of DA on osteogenic gene expression with receptor antagonists
To compare the osteogenic gene expression, the MC3T3-E1 cell culture was categorized as (i) cells cultured in growth media with or without DA, (ii) the cells cultured in osteogenic media with or without DA, and (iii) the cells blocked by antagonists and then cultured in both growth and osteogenic media with DA. Any elevated mRNA expression of osteogenic genes was considered to have higher osteogenic potential by the cells.
To confirm that DA treatment enhanced mineralization, we performed real-time PCR analysis to determine whether osteogenic gene expression is also increased in MC3T3-E1 cells. Our data showed that the expression of the osteogenic genes is different in cells with DA treatment than those without DA in osteogenic and growth media at either day 4 or day 7 (Figure 6).

The osteogenic gene expression of MC3T3‐E1 cells that were cultured with/without DA (50 µmol·L−1), as evaluated with real‐time PCR analysis to detect ALP, BSP, OC, OSX, RUNX2, and Collagen1a2 genes. Each gene data was presented as fold changes in the control, dopamine treated, and dopamine treated with antagonists (SCH233390 and GR103691) in growth media (a) and osteogenic media (b). The real‐time PCR data were normalized with GAPDH expression (n = 3 per group; *P < 0.05 compared with control and antagonists treated group at same time point).
Osteogenic media-stimulated osteogenic gene expression was higher than the osteogenic gene expression in growth media on both day 4 and day 7 (P < 0.05). However, the osteogenic gene expression in osteogenic media showed significant decrease on both day 4 and day 7 after DA treatment. Particularly, Runx2 gene was suppressed after blocking with combined antagonists for DA receptors, SCH233390 and GR103691, while the rest of the genes were elevated level of induction regardless of the antagonists treatment on day 4. But all the osteogenic genes in osteogenic media were decreased after antagonists treatment on day 7 except no obvious difference on OSX gene (Figure 6a).
DA alone induced osteogenic gene expression level. The level of each gene expression of MC3T3-E1 cells in growth media increased with DA treatment on either day 4 or day 7 except the collagen 1 gene expression on day 7, which did not show much effect on its gene expression even after DA treatment. The gene expression by DA treatment after antagonists treatment was significantly inhibited by the stimulatory effect of the DA except the collagen gene at 7 days. In contrast, the DA treatment on the cells cultured in the osteogenic media significantly reduced all the gene expression at both day 4 and day 7 (Figure 6b).
Discussion
Although the expression of serotonin and gamma-aminobutyric acid (GABA) receptors in osteoblasts was previously reported,17,18 the presence of DA receptors in osteoblasts has not been discussed. In this study, we found that DA receptors are expressed in MC3T3-E1 cells and are functional to induced mineralization. To our knowledge, this study is the first to report that osteoblasts express DA receptors and the cellular effect of DA treatment. DA receptors are usually known to have broad expression patterns not only in the central nervous system (CNS) such as brain and spinal cord but also in the peripheral nervous system (PNS).19,20 The roles and functions of DA and DA receptors in osteoblasts are unknown. In addition, there is no report of osteoblast releasing DA to date. We performed western blotting for tyrosine hydroxylase (TH) expression, which is responsible for catalyzing the conversion of the amino acid L-tyrosine to L-3,4-dihydroxyphenylalanine (L-DOPA) for cells that release DA. However, we did not detect any TH expression in osteoblasts even though it was detected in brain lysates used as a positive control (data not shown). The source of DA is likely extracelluar and not from osteoblasts themselves.
In this study, RT-PCR and western blot analysis were assessed to examine DA receptor mRNA and protein expression in MC3T3-E1 cells. Surprisingly, all five DA receptors genes and proteins were expressed as identified by the RT-PCR and western blot, respectively.
Although we did not measure direct affinity between DA and DA receptors, DA receptors (D1–D5) are known to be activated by DA with various affinities ranging from the nanomolar to micromolar range. Different subtypes of DA receptors vary significantly in their sensitivity to DA agonists and antagonists. The dissociation constants (Ki values) of the DA agonists were determined with selective radioligands. According to the reference by the National Institute of Mental Health Psychoactive Drug Screening Program database (http://pdsp.med.unc.edu), the Ki values for the five DA receptors were 130 nmol·L−1 for D1, 598 nmol·L−1 for D2, 32.5 nmol·L−1 for D3, 182.6 nmol·L−1 for D4, and 228 nmol·L−1 for D5.
Plasma concentration of DA in healthy human population is reported to be 21.8 ± 9.5 ng·L−1.21 Also, Mayo Clinic Lab reference value shows normal plasma concentration of DA should be less than 30 pg·mL−1 (=0.196 nmol·L−1). Although physiological DA concentration is much lower than what we used to stimulate MC3T3-E1 cells, there is no previous report on DA concentration in bone or DA effects on osteoblastic cells. Our study shows there was no clear cellular toxicity from DA concentration with 50 µmol·L−1 on osteoblasts. Although we stimulate with a higher concentration of DA than the physiological condition to osteoblasts, our study is limited on in vitro culture condition, not in vivo physiological conditions. If the cells stimulated by high concentration of DA can improve osteogenic differentiation in vitro without toxic effect, we can consider them as good candidate for bone regeneration in tissue engineering.
There is strong evidence in neuronal cells that DA can cause cytotoxic effects by both oxidative stress of reactive oxygen species (free radicals) and formation of hydrogen peroxide.22 Previously Clement et al. proved that DA-induced cell death is not specific only for neuronal cells but also for the rat pheochromocytoma cell line (PC12) and the M14 human melanoma cell line.23 Future dopaminergic toxicity test and apoptosis studies for MC3T3-E1 are warranted. One of the most reliable methods to detect the apoptotic cells affected by the DA is caspase activity assay, which detects a relatively early event in apoptosis. The detection method for caspase is reliable and does not require a washing step. Although the TUNEL assay is known as the gold standard to detect apoptosis, TUNEL detects necrotic cells as well. Moreover, it is a late event in apoptosis so there is a high chance to wash out dead cells during the staining procedures, which may result in inaccurate quantitation. It’s also a good way to apply DA antagonists to observe whether it can prevent MC3T3-E1 cell death at high dosage and this will show the apoptotic effect is from DA not from something else.
We further characterized the effect of DA on osteogenic activities by assessing proliferation, osteogenic differentiation and formation of mineralized bone-like nodules. While seeding with low number of cells (<20 000) can cause both inaccurate and inconsistent cell seeding, these cells divide rapidly and become confluent in only a few days. Thus, the initial cell number was optimized as 50 000 cells to allow the cells to be cultured for 7 days to avoid contact inhibition. Using a low serum medium for proliferation, we clarified the role that DA plays in promoting proliferation during 7 days of culture. Since FBS activates cellular mitogenic activity, it is important to test proliferation in low serum (3%) condition to observe better effect by DA, not by serum. Controls (no DA) were used and compared the proliferation taking into account the background response due to the presence of serum proteins in the culture medium. Proliferating cells were observed using BrdU assay. About 30% of cells were BrdU positive in each group except in the DA-treated group on day 1. BrdU positive cell number was higher than that in untreated group. Initially, DA could have an effect on the cell attachment.
While the nodules formed with DA treatment were larger and more branched, and connected with neighboring particles, the group without DA treatment showed smaller nodules that were not connected. Sometimes, discrepancies among the degree of mineral deposition may be due to differences in the differentiation stage, cell passage and condition culture period before differentiation triggered and the time that cells are grown in osteogenic media. Therefore, we also used growth media, in addition to osteogenic media to confirm the effect of DA on osteoblasts. Future studies can clarify the molecular mechanism of how DA stimulates mineralization through DA receptors. In contrast to other research studies that focused on the effect of cells on the coated substrates, this study provided clearer insight of the effect of DA on bone cells by adding a known concentration of DA directly to the culture media.
Real-time PCR was performed to quantify the expression of osteoblast-related genes (Table 2) in MC3T3-E1 cells after DA treatment in both growth and osteogenic media. Interestingly, all the genes investigated were significantly upregulated by culturing in growth media for either day 4 or day 7. To investigate whether DA affects osteogenic differentiation is specific via DA receptors, we used DA receptor antagonists SCH233390 (D1 and D5 blocker) and GR103691 (D3 blocker) to block DA receptors. The results demonstrated that gene expression, which was stimulated by DA treatment (50 µmol·L−1), was completely blocked by administration of the antagonists. However, all the gene expression of MC3T3-E1 cells cultured in osteogenic media decreased after DA treatment possibly due to a chemical reaction between the osteogenic media and DA. The antagonists also did not block the effect of the DA treatment.
Although gene expression in osteogenic media did not show any DA effects, the mineralization under osteogenic media was increased by DA. From these results, our assumption is that DA can directly interact with either β-glycerophosphate or ascorbic acid during osteogenic differentiation to decrease gene expression and further studies on the interaction should be investigated in future experiments. At the same time, DA antagonists also did not have any effect on most of the osteogenic gene expression in osteogenic media. One possible assumption is that the binding of the DA antagonists to the DA receptors might be inhibited by β-glycerophosphate or ascorbic acid. The other possibility is the reaction of antagonists with β-glycerophosphate or ascorbic acid changes their chemical conformation and prevents binding to the receptors. Further chemical analysis will be required before the osteogenic gene expression study by DA is attempted in osteogenic media. One limitation of our study is the use of multiple inhibitors at one time, which does not delineate the role of individual DA receptor. It is important to investigate the role of each DA receptor antagonist for confirmation of each DA receptor-mediated biological effects in the future.
In this study, DA receptors were expressed in MC3T3-E1 osteoblasts cells and are functional. Direct administration of DA-promoted cellular proliferation and mineralization and was confirmed by using specific DA receptor antagonists. Our findings suggest that DA can enhance bone regeneration, possibly in association with novel bone scaffold, HGCS-PDA scaffold in bone tissue engineering. Duel effect of DA on both osteoblasts and HGCS-PDA scaffold may lead to a novel strategy of bone regeneration for future orthopedic application.
References
Ungaro F, Biondi M, Indolfi L et al. Bioactivated Polymer Scaffolds for Tissue Engineering. In: Ashammakhi N, Reis RL . Topics in Tissue Engineering. 2006: 1–38.
Ko CC, Wang Z, Tseng HC, Lee DJ, Guez C . Design, synthesis, and evaluation of polydopamine-laced gelatinous hydroxyapatite nanocomposites for orthopedic applications. In: McKittrick JM, Narayan R, Lin H . Advances in Bioceramics and Biotechnologies II: Ceramic Transactions. 2014: 135–148.
Jiang J, Zhu L, Zhu L, Zhu B, Xu Y . Surface characteristics of a self-polymerized dopamine coating deposited on hydrophobic polymer films. Langmuir 2011; 27: 14180–14187.
Lee H, Scherer NF, Messersmith PB . Single-molecule mechanics of mussel adhesion. Proc Natl Acad Sci U S A 2006; 103: 12999–13003.
Lee H, Dellatore SM, Miller WM, Messersmith PB . Mussel-inspired surface chemistry for multifunctional coatings. Science 2007; 318: 426–430.
Tsai WB, Chen WT, Chien HW, Kuo WH, Wang MJ . Poly(dopamine) coating of scaffolds for articular cartilage tissue engineering. Acta Biomater 2011; 7: 4187–4194.
Ku SH, Ryu J, Hong SK, Lee H, Park CB . General functionalization route for cell adhesion on non-wetting surfaces. Biomaterials 2010; 31: 2535–2541.
Sun Y, Deng Y, Ye Z, Liang S, Tang Z, Wei S . Peptide decorated nano-hydroxyapatite with enhanced bioactivity and osteogenic differentiation via polydopamine coating. Colloids Surf B Biointerfaces 2013; 111: 107–116.
Rim NG, Kim SJ, Shin YM et al. Mussel-inspired surface modification of poly(L-lactide) electrospun fibers for modulation of osteogenic differentiation of human mesenchymal stem cells. Colloids Surf B Biointerfaces 2012; 91: 189–197.
Missale C, Nash SR, Robinson SW, Jaber M, Caron MG . Dopamine receptors: from structure to function. Physiol Rev 1998; 78: 189–225.
Serre CM, Farlay D, Delmas PD, Chenu C . Evidence for a dense and intimate innervation of the bone tissue, including glutamate-containing fibers. Bone 1999; 25: 623–629.
Lerner UH, Lundberg P . Kinins and neuro-osteogenic factors. Principles Bone Biol 2008; 1: 1025–1057.
Togari A . Adrenergic regulation of bone metabolism: possible involvement of sympathetic innervation of osteoblastic and osteoclastic cells. Microsc Res Tech 2002; 58: 77–84.
Chenu C, Marenzana M . Sympathetic nervous system and bone remodeling. Joint Bone Spine 2005; 72: 481–483.
Hanami K, Nakano K, Saito K et al. Dopamine D2-like receptor signaling suppresses human osteoclastogenesis. Bone 2013; 56: 1–8.
Bliziotes M, McLoughlin S, Gunness M, Fumagalli F, Jones SR, Caron MG . Bone histomorphometric and biomechanical abnormalities in mice homozygous for deletion of the dopamine transporter gene. Bone 2000; 26:15–19.
Wang D, Christensen K, Chawla K, Xiao G, Krebsbach PH, Franceschi RT . Isolation and characterization of MC3T3-E1 preosteoblast subclones with distinct in vitro and in vivo differentiation/mineralization potential. J Bone Miner Res 1999; 14: 893–903.
Maison SF, Liu XP, Eatock RA, Sibley DR, Grandy DK, Liberman MC . Dopaminergic signaling in the cochlea: receptor expression patterns and deletion phenotypes. J Neurosci 2012; 32: 344–355.
Fujimori S, Hinoi E, Yoneda Y . Functional GABA(B) receptors expressed in cultured calvarial osteoblasts. Biochem Biophys Res Commun 2002; 293: 1445–1452.
Westbroek I, van der Plas A, de Rooij KE, Klein-Nulend J, Nijweide PJ . Expression of serotonin receptors in bone. J Biol Chem 2001; 276: 28961–28968.
Ambade V, Arora BM, Singh CP, Somani BL, Basannar D . Adrenaline, noradrenaline and dopamine level estimation in depression: does it help? Med J Armed Forces India 2009; 65: 216–220.
Clement MV, Long LH, Ramalingam J, Halliwell B . The cytotoxicity of dopamine may be an artefact of cell culture. J Neurochem 2002; 81: 414–421.
Lai CT, Yu PH . Dopamine- and L-beta-3,4-dihydroxyphenylalanine hydrochloride (L-Dopa)-induced cytotoxicity towards catecholaminergic neuroblastoma SH-SY5Y cells. Effects of oxidative stress and antioxidative factors. Biochem Pharmacol 1997; 53: 363–372.
Acknowledgements
The authors would like to thank Mr. John Whitley for his assistance in manuscript preparation and Dr. He Zhang for her technical assistance. This work was supported in part by NIH/NIDCR K08DE018695, K12-EY016333/K08-EY021520 (HCT), and R01DE022816.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article

Cite this article
Lee, D., Tseng, H., Wong, S. et al. Dopaminergic effects on in vitro osteogenesis. Bone Res 3, 15020 (2015). https://doi.org/10.1038/boneres.2015.20
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/boneres.2015.20
This article is cited by
-
Polydopamine‐based surface functionalization of watermelon rind as a 3D nanofibrous cellulose scaffold for osteogenesis
Cellulose (2024)
-
Small-molecule amines: a big role in the regulation of bone homeostasis
Bone Research (2023)
-
Dopamine is involved in reparative dentin formation through odontoblastic differentiation of dental pulp stem cells
Scientific Reports (2023)
-
Activation of dopamine receptor D1 promotes osteogenic differentiation and reduces glucocorticoid-induced bone loss by upregulating the ERK1/2 signaling pathway
Molecular Medicine (2022)
-
A tailored bioactive 3D porous poly(lactic-acid)-exosome scaffold with osteo-immunomodulatory and osteogenic differentiation properties
Journal of Biological Engineering (2022)
