
Abstract
Affected and unaffected members of a Caucasian family with Werner syndrome were analyzed for mutations in the recently described Werner syndrome (WRN) gene and for their relevance to phenotypic expression of chromosomal instability and x-ray hypersensitivity. Two distinct molecular alterations were documented in the family. Analysis of the genomic DNA revealed a single-base exchange from A to T at an intron-exon boundary in the otherwise strongly conserved 5′ donor splice site. Consequently, exon 30 is spliced together with the intron. The ensuing structure could be confirmed by the presence and calculated size of the resulting RNA fragments. The patients, all compound heterozygotes, had a 1-bp deletion in the first third of the coding sequence in the other allele. The genotypes of the family members for these mutations were determined and consequences for the cellular phenotype of the otherwise unaffected heterozygotes are documented.
Similar content being viewed by others

Introduction
Werner syndrome is an inherited disease demonstrating features of premature aging as early loss and graying of hair, an ‘old face’, scleroderma-like skin with chronic ulceration, bilateral cataracts and other prematurely occurring age-related diseases, such as arteriosclerosis, osteoporosis, diabetes mellitus and cancer to such a degree that Werner syndrome is often considered to represent a partial phenocopy of human aging [1–3]. The relatively early death of these patients is primarily a consequence of cardiovascular breakdown or neoplasias [4]. In addition to the clinical appearance, cells from patients with Werner syndrome exhibit increased chromosomal instability, a reduced life span and a prolonged population-doubling time compared to age-matched controls [5–9]. These observations correlate well with parameters indicative of normal fibroblast senescence in culture [9].
Analysis of such cellular parameters in primary fibroblasts of 8 members of a family with Werner syndrome revealed a ‘Werner-like’ phenotype in otherwise unaffected siblings [10]. These findings gain importance in light of the Werner gene’s protein expression in heterozygotes and the reported relatively high incidence of cancer deaths in Werner syndrome families in Japan [2].
The gene for Werner syndrome (WRN) has recently been identified by positional cloning [11]. From the coding sequence, a protein of 1,432 amino acids has been deduced. Homology search revealed similarities with proteins of the helicase family. While the actual function of the protein is unknown, the presence of helicase motifs suggests a possible role in DNA metabolism. Mutations in the WRN gene’s coding region have been described in Japanese and Caucasian patients [11–14]. Patients have been found carrying homozygous mutations in the C-terminal, the N-terminal and also in the central helicase regions of the protein. Some patients also are compound heterozygotes. In order to determine whether the pheno-typic expression of Werner syndrome features in homozygotes and heterozygotes is influenced by the position of mutations in the protein, we characterized the mutations in the WRN gene in a Caucasian family with Werner syndrome.
Clinical Data and Family History
The family described originated from a village of Austria. The parents claimed to be nonconsanguineous. The mother died at age 49 from cancer. The father was 65 years and healthy at the time of skin biopsy. Two brothers, 35 and 26 years at the time of biopsy, were diagnosed clinically as typical Werner syndrome patients by their appearance (short stature: 165 cm; 157 cm, bird-like face, hypermelanosis) along with early cataracts, atrophic skin, diabetes mellitus, and osteoporosis. In both, hyaluronate excretion in the urine was markedly elevated. Subsequently, we confirmed the diagnosis by cellular parameters such as chromosomal instability, population doubling time, and life span of fibroblasts. The older brother had been married and had two children. He died at age 37 from myocardial infarction, acute leukemia and pneumonia.
A third brother, the oldest child, seems to be similarly affected, but refused further examination. One brother and one sister, 40 and 29 years of age at the time of biopsy, were phenotypically unaffected. Their cellular parameters of chromosomal instability and x-ray sensitivity, however, differed from unrelated age-matched controls.
Material and Methods
Cell Lines and Culture Conditions
All primary fibroblast cell strains (table 1) were established in our laboratory from skin biopsies and grown in minimum essential medium (MEM-Eagle, Life Technologies) supplemented with 10 or 15% fetal bovine serum (Life Technologies) without antibiotics at 37 °C with 5% CO2 in the atmosphere and 90% humidity.
Mycoplasma infection was excluded.
Lymphocytes (table 1) were cultivated from heparinized whole blood in RPMI-1640 medium supplemented with 25% fetal bovine serum and phytohemagglutinin according to conventional methods [15].
Micronucleus Assay
1 × 105 cells were seeded on slides and cultivated in MEM supplemented with 10% fetal bovine serum. After 60 h, the cells were washed twice with phosphate-buffered saline (PBS) fixed with ice-cold methanol and stained with bis-benzimide (Sigma, working solution 1 µg/ml in PBS) for 30 min. Slides where washed with PBS and coverslips were mounted with PBS/glycerol/distilled water (3:1:1). Cells (1,000–2,000 per slide) were scored for micronuclei using an epifluorescence microscope with phase contrast (Zeiss), according to Countryman and Heddle [16].
X-Ray Irradiation and Scoring of Chromosomes
1.5 × 105 fibroblasts were cultivated on slides for 24 h before irradiation. Lymphocyte cultures were irradiated 3 h before harvest. Cells were irradiated with a modified Müller RT 250 Mundastat generator (220 kV, 15 mA, 1 mm copper filter, dose rate 1 Gy/min, irradiation distance 20 cm) at room temperature. Fibroblasts were irradiated with 0.25 and 0.5 Gy, lymphocytes with 0.25, 0.5 and 1.0 Gy. Immediately after irradiation, colcemid was added to irradiated and nonirradiated cultures. Preparation of chromosomes was performed according to standard methods [15]. 50 well-spread Giemsa-stained integral metaphases per slide were scored for chromosomal aberrations.
Nomenclature and criteria for chromosomal breaks were based on the ISCN (1978). Structural rearrangements, dicentrics and rings were considered as two breakage events. Chromatid and isochromatid breaks and deletions were scored as single events. Achromatic gaps were neglected.
Genomic DNA Isolation
Genomic DNA from fibroblasts was isolated as described [17] with the following modification: after phenol:chloroform extraction, the pooled aqueous phases were dialysed against 50 mM Tris/HCl pH 8.0, 10 mM EDTA pH 8.0 over night. The dialysed DNA probe was precipitated by adding 0.2 vol of 10 M ammonium acetate and 2 vol of ethanol.
Polymerase Chain Reaction
The polymerase chain reaction (PCR) was performed with DNA-Thermal Cycler 480 (Perkin Elmer) and Thermocycler 60 (Bio-med). Different Taq DNA polymerases were used (Taq DNA Pol, Gibco BRL; Combi Pol™ DNA Pol Mix, Invitek; InviTaq™ DNA Pol, Invitek and UlTma™ DNA Pol, Perkin Elmer). For the splice mutation, genomic DNA was annealed at 56 °C with Mut 1 h primer: 5′gAT gTg ACA gTg gAA gCT ATg g and Mut 1 v primer: 5′ggA AAA ATg Tgg TAT CTg AAg CTC. The PCR product was cloned into the pCR′ 2.1 Vector (Invitrogen) and transfected into Escherichia coli NM522. Werner syndrome cDNA was annealed at 56 °C with Werner syndrome vneu primer: 5′CTg Agg gAT TCT Tgg TAg AAg and Werner syndrome m primer: 5′TTC TTg Agg TTT TgT ACT TgA A. The 1-bp deletion was found in a PCR product created with the 1267v primer: 5′CAT ggg ACC CAA CAC TTg ATC and 1445h primer: 5′CCT gAg ACT gCT gTT CC at 62 °C annealing temperature.
DNA Sequencing
DNA sequencing was done with Sequenase Version 2.0 DNA Sequencing Kit (USB) and T7 Sequencing™ Kit (Pharmacia) with 35S ATP and M13 primer (5′AAC AgC TAT gAC CAT g) and T7 primer (5′TAA TAC gAC TCA CTA TA).
Additional DNA sequencing was performed with Thermo Sequenase fluorescence-labeled primer cycle sequencing Kit with 7-deaza-dGTP (Amersham Life Science) and the DNA Sequencer model 4000L LI-COR (MWG-Biotech).
RNA Isolation and cDNA Synthesis
Isolation of RNA from fibroblasts was performed as described [17].
Aliquots of 1.5 µg RNA were used for cDNA synthesis [17] with the following modification: after incubation for 60 min at 37 °C for first strand synthesis, 30 µl distilled water was added and the reverse transcriptase was heat inactivated (80 °C for 10 min). RNA was digested with 1 µl RNAse H (1 U/µl) for 1 h at 37 °C and after inactivating the enzyme by heating, the single-strand cDNA was stored at −80°C.
Results
Skin biopsies and lymphocytes were obtained from 5 members of a nonconsanguineous family with two typical Werner syndrome probands. Fibroblasts were established and were screened periodically during their entire life span for spontaneous chromosomal instability. Micronuclei which are suitable indicators for chromosomal breakage events were increased in Werner syndrome patients and to some extent in unaffected siblings, when compared to age-matched controls (fig. 1). Fibroblasts and lymphocytes were analyzed for their reaction to DNA damaging agents. Werner syndrome patients as well as siblings and their father exhibited a hypersensitivity towards x-ray irradiation in both cell types (fig. 2). To correlate these observations to possible mutations in the recently reported WRN gene [11], we prepared genomic DNA from all family members and unrelated controls and screened the WRN gene for mutations.

Spontaneous formation of micronuclei in fibroblasts of WS patients, family members and unrelated controls. Micronuclei per 1,000 cells are scored in 4 unrelated controls (IC1–4, n = 55), in 2 patients with Werner syndrome (LW3, n = 15; LW4, n = 18) in 2 heterozygous siblings (LwI, n = 18; LwK, n = 6) and the heterozygous father (LwH, n = 16). The standard deviations are given.

Chromosomal breaks induced by x-ray irradiation in fibroblasts (a) and lymphocytes (b). Werner syndrome family members are compared to age-matched controls. Several controls were analysed and averaged; the error bars are shown as well. The numbers of chromosomal breaks per 100 metaphase cells are shown on the ordinate. a1, b1 Werner syndrome patients. a2, b2 Heterozygous siblings. a3, b3 Heterozygous father.
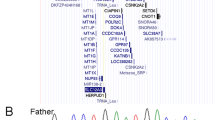
PCR fragments of the 3′-end region of the WRN gene were amplified and sequenced. In one of the fragments a single base was changed from A to T in some family members. This base was positioned at an exon-intron boundary at the highly conserved 5′ CpA sequence of a splice donor site (fig. 3). Such a mutation would result in a deletion of the 113-bp long exon heading the intron 5′ and cause a frameshift resulting in a termination signal (TGA) at nucleotide (nt) 3816. In agarose gel electrophoresis, RT-PCR products of this region showed two bands in the Werner syndrome patients, the father and the unaffected brother, but only one band in the healthy sister and in unrelated controls. The length of the shorter band reflected the length of the missing exon well (fig. 4).

Consensus sequence of the 5’ donor splice site. One exon-intron boundary is given in the WRN genomic DNA and in the deduced pre-mRNA. Conserved splice-site sequence is given in bulky letters and is changed from ca to ct in the mutated allele.

RT-PCR products obtained from mRNA fragments amplified by the oligonucleotides described in Material and Methods. A fragment of 650 bp (bp markers: lane 4) was expected and found in fibroblasts of a healthy, unrelated control (lane 1). In the Werner syndrome patients LW3 (lane 2) and LW4 (lane 3), this band and an additional one of 537 bp in length is shown. The smaller band corresponds to the RNA in which one exon is missing. The two bands from the father and the brother and the single band from the healthy sister are not shown.
Subsequent cloning and sequencing of the two bands proved that they represented the normal and the altered fragments. Both patients, the father and the brother, were shown to be heterozygous for the splice mutation.
The exon loss causes a frameshift which results in a stop codon and a protein that is 275 amino acids shorter than the normal. Furthermore, at position nt 3453, a T was changed to a G in the father’s normal-length fragment. This exchange, resulting in a codon for Leu instead of Phe, has to be considered a polymorphism since the father carries this allele but is phenotypically unaffected. This polymorphism helped to trace back the alleles in the offspring.
Taking into account that the patients in this family expressed Werner syndrome symptoms rather early, hypermelanosis and growth retardation around 13, cataract operations at 23 and 25 years of age, we expected a mutation in the second allele to be found in a more 5′ position. Mutations in the N-terminal region or in other regions eliminating the helicase domains may result in a protein with complete loss of function. Indeed, we were able to localize the second mutation as a 1-bp deletion at nt 1396. This deletion led to a frameshift and created a stop codon at nt 1406–1408 resulting in a protein of 391 amino acids. In addition, close to that deletion, a polymorphism was found at nt 1392 changing G to A (Met to Isoleu).
Both patients and the unaffected sister carried this allele, that they inherited from their prematurely deceased mother. In addition, this allele carried the same polymorphism at nt 3453 as the father’s normal allele. This coincidence and the fact that we did not find this alteration in unrelated controls may give rise to the suspicion that the two alleles may be of identical descent. One of the ancestors may have acquired the 1-bp deletion as a new mutation (fig. 5).

Pedigree of a Caucasian Werner syndrome family. The allele distribution of the WRN gene (A, B, C, D) is given for the parents and 4 siblings of the first generation. The ages at time of biopsy and of death (2 family members) are indicated. WRN cDNA and WRN protein are given schematically for all 4 alleles. Mutations and polymorphisms are recorded on the cDNA, the latter with stars and the nucleotide in question in brackets. The length of the bars drawn underneath of the cDNAs gives the length of the predicted protein.
While the specific splice mutation at a donor splice site downstream from exon 30 has not been described as yet in any of the analyzed Werner syndrome patients, the 1-bp deletion probably exists as a heterozygous mutation [12, 14].
Discussion
The function of the WRN gene is as yet unknown. The characterization of mutations in the gene remains the sole method to correlate the mutated genotype with the clinical phenotypic Werner syndrome manifestations. This was the purpose of our Werner syndrome family study. In our affected family, 2 brothers, both compound heterozygotes, were afflicted at an early age. One had died at age 37. They had inherited one allele from their father which was mutated in the C-terminal of the protein. The father, a heterozygote, was still alive. The mother, however, having been heterozygous for the allele with the N-terminal mutation, had died prematurely at age 49 from cancer, one of the symptoms in Werner syndrome. One of the mutated alleles could be found in each of her 2 unaffected children. We have recently reported low but significant expression of cellular features typical for Werner syndrome in primary fibroblasts of otherwise unaffected members of another family with Werner syndrome [10]. These features included chromosomal instability, a shortened life span and prolongation of population doubling time that normally would be found in older individuals, indicating the close correlation between senescence and DNA repair [9]. We also found pathological reactions towards exogenous DNA damage not only in the patients but also in their siblings [unpubl. results]. In our recent study, the young age of the phenotypically normal family members, especially the sister, was noteworthy. The increased chromosomal instability in her fibroblasts was evidence that she was heterozygous for the defective gene before her actual genotype had been identified. The same was true, but to a lesser extent, for the 10-year older brother and the father, who both bore the splice mutation. From these observations we assume that the Werner syndrome protein functions in a dose-dependent manner in which both the amount and the integrity of the protein are important. As in other metabolic diseases of autosomal recessive inheritance, the reduced enzymatic efficiency of the WRN protein in heterozygotes results in an elevated intolerance towards endogenous and exogenous DNA damage. Even though involvement of the protein in DNA repair was generally negated [15–18], the coincidence of WRN gene mutations with spontaneous as well as DNA-damage-induced alterations of the cellular phenotype in heterozygotes indicates a role for the enzyme in DNA repair metabolism and encourages further research in this direction.
References
Werner CWO: Katarakte in Verbindung mit Hautveränderungen, Diss. Kiel, 1904.
Goto M, Tanimoto K, Horiuchi Y, Sasazuki T: Family analysis of Werner syndrome: A survey of 42 Japanese families with a review of literature. Clin Genet 1981;19:8–15.
Epstein CJ, Martin GM, Schultz AL, Motulsky AG: Werner’s syndrome. A review of its symptomatology, natural history, pathologic features, genetics, and relationship to the natural aging process. Medicine 1966;45:177–221.
Goto M, Takeuchi F, Miyamoto K: Clinical, demographic, and genetic aspects of the Werner syndrome in Japan. Adv Exp Med Biol 1985;190:245–261.
Salk D: Werner syndrome: A review of recent research with an analysis of connective tissue metabolism, growth control of cultured cells, and chromosomal aberrations. Hum Genet 1982;62:1–15.
Martin GM, Sprague CA, Epstein CJ: Replicative life-span of cultivated human cells. Effect of donor age, tissue, genotype. Lab Invest 1970; 23:86–92.
Salk D, Bryant K, Hoehn H, Martin GM: Systematic growth studies, cocultivation, and cell hybridisation studies of Werner syndrome cultured skin fibroblasts. Hum Genet 1981;58: 310–316.
Poot M, Hoehn H, Rünger T, Martin GM: Impaired S-phase transit of Werner syndrome cells expressed in lymphoblastoid cell lines. Exp Cell Res 1992;202:267–273.
Weirich-Schwaiger H, Weirich HG, Gruber B, Schweiger M, Hirsch-Kauffmann M: Correlation between senescence and DNA repair in cells from young and old individuals and in premature aging syndromes. Mutat Res 1994; 316:37–48.
Weirich HG, Weirich-Schwaiger H, Kofler K, Sidoroff A, Fritsch P, Schachtschabel DO, Schweiger M, Hirsch-Kauffmann M: Werner syndrome: Studies in an affected family reveal a cellular phenotype of unaffected siblings. Mech Ageing Dev 1996;88:1–15.
Yu CE, Oshima J, Fu YW, Hisama F, Wijsman EM, Alisch R, Matthews S, Nakura J, Miki T, Ouais S, Martin GM, Mulligan J, Schellenberg GD: Positional cloning of the Werner’s syndrome gene. Science 1996;272:258–262.
Oshima J, Yu CE, Piussan C, Klein G, Jabkowski J, Balci S, Miki T, Nakura J, Ogihara T, Ells J, Smith M de AC, Melaragno MI, Fraccaro M, Scappaticci S, Matthews J, Ouais S, Jarzebowicz A, Schellenberg GD, Martin GM: Homozygous and compound heterozygous mutations at the Werner syndrome locus. Hum Mol Genet 1996;5:1909–1913.
Goto M, Imamura O, Kuromitsu J, Matsumoto T, Yamabe Y, Tokutake Y, Suzuki N, Mason B, Drayna D, Sugawara M, Sugimoto M, Furuichi Y: Analysis of helicase gene mutations in Japanese Werner’s syndrome patients. Hum Genet 1997;99:191–193.
Yu CE, Oshima J, Wijsman EM, Nakura J, Miki T, Piussan C, Matthews S, Fu YH, Martin GM, Schellenberg GD and the Werner’s Syndrome Collaborative Group: Mutations in the consensus helicase domains of the Werner syndrome gene. Am J Hum Genet 1997;60:330–341.
Verma RS, Babu A: Human Chromosomes. Manual of Basic Techniques, New York, Pergamon Press, 1989.
Countryman PL, Heddle JA: The production of micronuclei from chromosome aberrations on irradiated cultures of human lymphocytes. Mutat Res 1976;41:321–332.
Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning, A Laboratory Manual, ed 2, 1989.
Stefanini M, Scappaticci S, Lagomarsini P, Borroni G, Berardesca E, Nuzzo F: Chromosome instability in lymphocytes from patients with Werner’s syndrome is not associated with DNA repair defects. Mutat Res 1979;219:179–185.
Saito H, Moses RE: Immortalization of Werner syndrome and progeria fibroblasts. Exp Cell Res 1991;192:373–379.
Fujiwara Y, Higashikawa T, Tatsumi M: A retarded rate of DNA replication and normal level of DNA repair in Werner’s syndrome fibroblast cultures. J Cell Physiol 1977;92:365–374.
Higashikawa T, Fujiwara Y: Normal level of unscheduled DNA synthesis in Werner’s syndrome fibroblasts in culture. Exp Cell Res 1978;113:438–441.
Acknowledgements
We are indebted to the Thyssen-Stiftung and to the ‘Fonds der Chemischen Industrie’ for financial support. The work was further supported by the ‘Deutsche Forschungsgemeinschaft’ SCHW 532/2-1 and by the ‘Österreichische Nationalbank, Jubiläumsfonds 4842’. Determination of hyaluronate in urine was performed by Dr. H. Greiling, Aachen, Germany. We are also grateful to Dr. D.H. Kauffmann Jockl, New York, N.Y., for critical reading and improving the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Meißlitzer, C., Ruppitsch, W., Weirich-Schwaiger, H. et al. Werner Syndrome: Characterization of Mutations in the WRN Gene in an Affected Family. Eur J Hum Genet 5, 364–370 (1997). https://doi.org/10.1007/BF03405944
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF03405944
