
Abstract
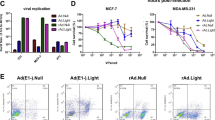
Telomerase is a therapeutic target for cancer. Human telomerase reverse transcriptase (hTERT), the catalytic subunit of the telomerase, is transcriptionaly upregulated exclusively in about 90% of cancer cells. Previous studies have demonstrated that hTERT promoter can control the expression of exogenous genes to the telomerase-positive cancer cells, thus hTERT promoter is an excellent candidate for generating cancer-specific oncolytic adenovirus. In this study, we devised a novel oncolytic adenovirus (Ad.TERT) by replacing the normal E1A regulatory elements with hTERT promoter. Ad.TERT displays cancer-specific E1A expression, virus replication and cytolysis in in vitro experiments. In animal experiments, intratumoral administration of Ad.TERT demonstrates potent antitumoral efficacy at least in two xenograft models (Bcap37 and BEL7404). Ad.TERT was targeted by the telomerase activity in cancer cells and has potent antitumoral efficacy in vivo, and since telomerase activity is a wide-ranged tumor marker, Ad.TERT could be a powerful therapeutic agent for a variety of cancers.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

References
Alemany R, Balague C and Curiel D . (2000). Nat. Biotechnol., 18, 723–727.
Bilsland AE, Anderson CJ, Fletcher-Monaghan AJ, McGregor F, Evans TR, Ganly I, Knox RJ, Plumb JA and Keith WN . (2003). Oncogene, 22, 370–380.
Blackburn E . (1991). Nature, 350, 569–573.
Bodnar A, Kim N, Effros R and Chiu C . (1996). Exp. Cell Res., 228, 58–64.
Chiu C-P, Dragowska W, Kim NW, Vaziri H, Yui J, Thomas TE, Harley CB and Lansdorp PM . (1996). Stem Cells, 14, 239–248.
Cong Y, Wen J and Bacchetti S . (1999). Hum. Mol. Genet., 8, 137–142.
Curiel DT . (2000). Clin. Cancer Res., 6, 3395–3399.
Feng J, Funk WD, Wang SS, Weinrich SL, Avilion AA, Chiu CP, Adams RR, Chang E, Allsopp RC and Yu J (1995). Science, 269, 1236–1241.
Freytag S, Rogulski K, Paielli D, Gilbert J and Kim J . (1998). Hum. Gene Ther., 9, 1323–1333.
Gu J, Kagawa S, Takakura M, Kyo S, Inoue M, Roth JA and Fang B . (2000). Cancer Res., 60, 5359–5364.
Gu J, Zhang L, Huang X, Lin T, Yin M, Xu K, Ji L, Roth JA and Fang B . (2002). Oncogene, 21, 4757–4764.
Hallenbeck P, Chang Y, Hay C, Golightly D, Stewart D, Lin J, Phipps S and Chiang Y . (1999). Hum. Gene Ther., 10, 1721–1733.
Hernandez-Alcoceb R, Pihalja M, Qian D and Clarke M . (2002). Hum. Gene Ther., 13, 1737–1750.
Horwitz MS . (1996). Fields, BN, Knipe DM and Howley PM (ed.) Fields Virology. Lippincott-Raven Publishers: Philadelphia, pp 2149–2171.
Ichikawa T and Chiocca EA . (2001). Cancer Res., 61, 5336–5339.
Jakubczak JL, Ryan P, Gorziglia M, Clarke L, Hawkins LK, Hay C, Huang Y, Kaloss M, Marinov A, Phipps S, Pinkstaff A, Shirley P, Skripchenko Y, Stewart D, Forry-Schaudies S and Hallenbeck PL . (2003). Cancer Res., 63, 1490–1499.
Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, Coviello GM, Wright WE, Weinrich SL and Shay JW . (1994). Science, 266, 2011–2015.
Kirch HC, Ruschen S, Brockmann D, Esche H, Horikawa I, Barrett JC, Opalka B and Hengge UR . (2002). Oncogene, 21, 7991–8000.
Kirn D, Martuza R and Zwiebel J . (2001). Nat. Med., 7, 781–787.
Komata T, Kanzawa T, Kondo Y and Kondo S . (2002). Oncogene, 21, 656–663.
Kruyt F and Curiel D . (2002). Hum. Gene Ther., 13, 485–495.
Kurihara T, Brough DE, Kovesdi I and Kufe DW (2000). J. Clin. Invest., 106, 763–771.
Li Y, Yu D-C, Chen Y, Amin P, Zhang H, Nguyen N and Henderson DR . (2001). Cancer Res., 61, 6428–6436.
Lin T, Huang X, Gu J, Zhang L, Roth JA, Xiong M, Curley SA, Yu Y, Hunt KK and Fang B . (2002). Oncogene, 21, 8020–8028.
Liu J, Zou W, Lang M, Luo J, Sun L, Wang X, Qian Q and Liu X . (2002). Int. J. Oncol., 21, 661–666.
Matsubara S, Wada Y, Gardner TA, Egawa M, Park M-S, Hsieh C-L, Zhau HE, Kao C, Kamidono S, Gillenwater JY and Chung LWK . (2001). Cancer Res., 61, 6012–6019.
McKinnon R, Bacchetti S and Graham F . (1982). Gene, 19, 33–42.
Meyerson M, Counter C, Eaton E, Ellisen L, Steiner P, Caddle S, Ziaugra L, Beijersbergen R, Davidoff M, Liu Q, Bacchetti S, Haber D and Weinberg R . (1997). Cell, 90, 785–795.
Motoi F, Sunamura M, Ding L, Duda D, Yoshida Y, Zhang W, Matsuno S and Hamada H . (2000). Hum. Gene Ther., 11, 223–235.
Nakamura Y, Tahara E, Tahara H, Yasui W and Ide T . (1999). Mol. Carcinog., 26, 312–320.
Nakayama J, Saito M, Nakamura H, Matsuura A and Ishikawa F . (1997). Cell, 88, 875–884.
Nettelbeck D, Jerome V and Muller R . (2000). Trends Genet., 16, 174–181.
Plumb JA, Bilsland A, Kakani R, Zhao J, Glasspool RM, Knox RJ, Evans TR and Keith WN . (2001). Oncogene, 20, 7797–7803.
Post DE and Van Meir EG . (2003). Oncogene, 22, 2065–2072.
Ramirez R, Wright W, Shay J and Taylor R . (1997). J. Invest. Dermatol., 108, 113–117.
Rodriguez R, Schuur E, Lim H, Henderson G, Simons J and Henderson D . (1997). Cancer Res., 57, 2559–2563.
Shay J . (1997). J. Cell Physiol., 173, 266–270.
Takakura M, Kyo S, Kanaya T, Hirano H, Takeda J, Yutsudo M and Inoue M . (1999). Cancer Res., 59, 551–557.
Tsukuda K, Wiewrodt R, Molnar-Kimber K, Jovanovic VP and Amin KM . (2002). Cancer Res., 62, 3438–3447.
Wang J, Yao M, Gu J, Sun L, Shen Y and Liu X . (2002). Biochem. Biophys. Res. Commun., 290, 1454–1461.
Watanabe T, Kuszynski C, Ino K, Heimann D, Shepard H, Yasui Y, Maneval D and Talmadge J . (1996). Blood, 87, 5032–5039.
Whyte P, Ruley HE and Harlow E . (1988). J. Virol., 62, 257–265.
Wildner O, Morris J, Vahanian N, Ford H, Ramsey W and Blaese R . (1999). Gene Ther., 6, 57–62.
Yasumoto S, Kunimura C, Kikuchi K, Tahara H, Ohji H, Yamamoto H, Ide T and Utakoji T . (1996). Oncogene, 13, 433–439.
Yu D-C, Chen Y, Dilley J, Li Y, Embry M, Zhang H, Nguyen N, Amin P, Oh J and Henderson DR . (2001). Cancer Res., 61, 517–525.
Yu D-C, Chen Y, Seng M, Dilley J and Henderson DR . (1999). Cancer Res., 59, 4200–4203.
Acknowledgements
We thank Bingliang Fang for providing us with the hTERT promoter. We also thank Lanyin Sun for help in the cell culture, Weijing Xu and Binghua Li for critical reading and discussion of this paper. This work was supported by the Key Project of the Chinese Academy of Sciences (No. KSCX2-3-06), the National Natural Science Foundation of China (No.30120160823), and the State 863 High Technology R&D Project of China (No. 2001AA217031).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zou, W., Luo, C., Zhang, Z. et al. A novel oncolytic adenovirus targeting to telomerase activity in tumor cells with potent. Oncogene 23, 457–464 (2004). https://doi.org/10.1038/sj.onc.1207033
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.onc.1207033
Keywords
This article is cited by
-
Mechanism of action and treatment of type I interferon in hepatocellular carcinoma
Clinical and Translational Oncology (2023)
-
Synergistic suppression effect on tumor growth of hepatocellular carcinoma by combining oncolytic adenovirus carrying XAF1 with cisplatin
Journal of Cancer Research and Clinical Oncology (2015)
-
Cancer targeting Gene-Viro-Therapy of liver carcinoma by dual-regulated oncolytic adenovirus armed with TRAIL gene
Gene Therapy (2011)
-
Targeting strategies for adeno-associated viral vector
Chinese Science Bulletin (2007)
-
Targeting gene-virotherapy of cancer
Cell Research (2006)
