
Abstract
In this overview we have summarized some aspects of our published work related to the effects of the endocannabinoid system on appetite and suckling. As noted also by several other groups we have found that anandamide, a major endocannabinoid, enhances appetite in mice. On partial or full food deprivation over 24 h the levels of 2-arachidonoyl glycerol (2-AG), a second major cannabinoid, are initially elevated in mouse brain; however, partial food deprivation over a longer period causes reduction of 2-AG levels. Blocking the endocannabinoid system with a CB1 antagonist on the 1st day after birth leads to inhibition of suckling; later administration also affects suckling, but does not fully block it.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

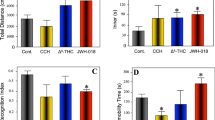
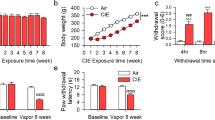
References
Mechoulam R . The Pharmacohistory of Cannabis sativa. In: Mechoulam R (ed), The Cannabinoids as Therapeutic Agents. CRC Press Inc.: Boca Raton, FL, 1986, pp 1–19.
Gaoni Y, Mechoulam R . Isolation, structure and partial synthesis of an active constituent of hashish. J Am Chem Soc 1964; 86: 1646–1647.
Di Tomaso E, Beltramo M, Piomelli D . Brain cannabinoids in chocolate. Nature 1996; 382: 677–678.
Di Marzo V, Sepe N, De Petrocellis L, Berger A, Crozier G, Fride E et al. Trick or treat from food endocannabinoids? Nature 1998; 396: 636.
Fride E, Ginzburg Y, Breuer A, Bisogno T, Di Marzo V, Mechoulam R . Critical role of the endogenous cannabinoid system in mouse pup suckling and growth. Eur J Pharmacol 2001; 419: 207–214.
Zimmer A, Zimmer AM, Hohmann AG, Herkenhanm M, Bonner TI . Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc Natl Acad Sci USA 1999; 96: 5780–5785.
Petrov ES, Varlinskaya EI, Smotherman WP . Endogenous opioids and the first suckling episode in the rat. Dev Psychobiol 1998; 33: 175–183.
Contos JJ, Fukushima N, Weiner JA, Kaushal D, Chun J . Requirement for the lpA1 lysophosphatidic acid receptor gene in normal suckling behavior. Proc Natl Acad Sci USA 2000; 97: 13384–13389.
Moolenaar WH . Development of our current understanding of bioactive lysophospholipids. Ann NY Acad Sci 2000; 905: 1–10.
Fride E, Foox A, Rosenberg E, Faigenboim M, Cohen V, Barda L et al. Milk intake and survival in newborn cannabinoid CB1 receptor knockout mice: evidence for a ‘CB3’ receptor. Eur J Pharmacol 2003; 461: 27–34.
Breivogel CS, Griffin G, Di Marzo V, Martin BR . Evidence for a new G protein-coupled cannabinoids receptor in mouse brain. Mol Pharmacol 2001; 60: 155–163.
Berry EM, Mechoulam R . Tetrahydrocannabinol and endocannabinoids in feeding and appetite. Pharmacol Ther 2002; 95: 185–190.
Kirkham TC, Williams CM . Endogenous cannabinoids and appetite. Nutr Res Rev 2001; 14: 65–86.
Williams CM, Kirkham TC . Anandamide induces overeating: mediation by central cannabinoid (CB1) receptors. Psychopharmacol 1999; 143: 315–317.
Hao S, Avraham Y, Mechoulam R, Berry EM . Low dose anandamide affects food intake, cognitive function, neurotransmitter and corticosterone levels in diet-restricted mice. Eur J Pharmacol 2002; 392: 147–156.
Šulcová E, Mechoulam R, Fride E . Biphasic effects of anandamide. Pharmacol Biochem Behav 1998; 59: 347–352.
Paton WDM, Pertwee RG . The pharmacology of cannabis in animals. In: Mechoulam R (ed). Marijuana, Chemistry, Pharmacology, Metabolism and Clinical Effects. Academic Press: New York, 1973, pp 191–285.
McLaughlin CL, Baile CA, Bender PE . Cannabinols and feeding in sheep. Psychopharmacol 1979; 64: 321–323.
Berrendero F, Sepe N, Ramos JA, Di Marzo V, Fernandez-Ruiz JJ . Analysis of cannabinoid receptor binding and mRNA expression and endogenous cannabinoid contents in the developing rat brain during late gestation and early postnatal period. Synapse 1999; 33: 181–191.
Mechoulam R, Ben-Shabat S, Hanuš L, Kaminski NE, Schatz AR, Gopher A et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol 1995; 50: 83–90.
Mechoulam R, Fride E, Ben-Shabat S, Meiri U, Horowitz M . Carbachol, an acetylcholine receptor agonist, enhances production in rat aorta of 2-arachidonoyl glycerol, a hypotensive endocannabinoid. Eur J Pharmacol 1998; 362: R1–R3.
Panikashvili D, Simeonidou C, Ben-Shabat S, Hanuš L, Breuer A, Mechoulam R et al. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury. Nature 2001; 413: 527–531.
Kirkham TC, Williams CM, Fezza F, Di Marzo V . Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: stimulation of eating by 2-arachidonoyl glycerol. Br J Pharmacol 2002; 136: 550–557.
Hanuš L, Avraham Y, Ben-Shushan D, Zolotarev O, Berry EM, Mechoulam R . Short term fasting and prolonged semistarvation have opposite effect on 2-AG levels in mouse brain. Brain Res 2003; 983: 144–151.
Berger A, Crozier G, Bisogno T, Cavaliere P, Innis S, Di Marzo V . Anandamide and diet: inclusion of dietary arachidonate and docosahexaenoate leads to increased brain levels of the corresponding N-acylethanolamines in piglets. Proc Natl Acad Sci USA 2001; 98: 6402–6406.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mechoulam, R., Berry, E., Avraham, Y. et al. Endocannabinoids, feeding and suckling – from our perspective. Int J Obes 30 (Suppl 1), S24–S28 (2006). https://doi.org/10.1038/sj.ijo.0803274
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.ijo.0803274
Keywords
This article is cited by
-
Medical Marijuana: Current Concepts, Pharmacological Actions of Cannabinoid Receptor Mediated Activation, and Societal Implications
Current Pain and Headache Reports (2018)
