
Abstract
Reverse transcriptases, used in prime editing systems, exhibit lower fidelity, processivity and dNTP affinity than many DNA-dependent DNA polymerases. We report that a DNA-dependent DNA polymerase (phi29), untethered from Cas9, enables editing from a synthetic, end-stabilized DNA-containing template at up to 60% efficiency in human cells. Compared to prime editing, DNA polymerase editing avoids autoinhibitory intramolecular base pairing of the template, facilitates template synthesis and supports larger insertions (>100 nucleotides).
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others


Data availability
A Reporting Summary for this article is available as a supplementary information file. Plasmids for mRNA in vitro transcription, including MCP–RT, MCP–phi29 and nCas9 (H840A), have been deposited to Addgene for distribution. Illumina Sequencing data have been submitted to the Sequence Read Archive, and datasets are available under BioProject accession number PRJNA1004245 (ref. 37). Source data are provided with this paper.
References
Anzalone, A. V. et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 576, 149–157 (2019).
Lin, Q. et al. High-efficiency prime editing with optimized, paired pegRNAs in plants. Nat. Biotechnol. 39, 923–927 (2021).
Zhuang, Y. et al. Increasing the efficiency and precision of prime editing with guide RNA pairs. Nat. Chem. Biol. 18, 29–37 (2022).
Wang, J. et al. Efficient targeted insertion of large DNA fragments without DNA donors. Nat. Methods 19, 331–340 (2022).
Choi, J. et al. Precise genomic deletions using paired prime editing. Nat. Biotechnol. 40, 218–226 (2022).
Jiang, T., Zhang, X.-O., Weng, Z. & Xue, W. Deletion and replacement of long genomic sequences using prime editing. Nat. Biotechnol. 40, 227–234 (2022).
Chen, P. J. & Liu, D. R. Prime editing for precise and highly versatile genome manipulation. Nat. Rev. Genet. 24, 161–177 (2023).
Liu, B. et al. A split prime editor with untethered reverse transcriptase and circular RNA template. Nat. Biotechnol. 40, 1388–1393 (2022).
Nelson, J. W. et al. Engineered pegRNAs improve prime editing efficiency. Nat. Biotechnol. 40, 402–410 (2022).
Liu, Y. et al. Enhancing prime editing by Csy4-mediated processing of pegRNA. Cell Res. 31, 1134–1136 (2021).
Zhang, G. et al. Enhancement of prime editing via xrRNA motif-joined pegRNA. Nat. Commun. 13, 1856 (2022).
Li, X. et al. Enhancing prime editing efficiency by modified pegRNA with RNA G-quadruplexes. J. Mol. Cell. Biol. 14, mjac022 (2022).
Feng, Y. et al. Enhancing prime editing efficiency and flexibility with tethered and split pegRNAs. Protein Cell 14, 304–308 (2022).
Ponnienselvan, K. et al. Reducing the inherent auto-inhibitory interaction within the pegRNA enhances prime editing efficiency. Nucleic Acids Res. 51, 6966–6980 (2023).
Ellefson, J. W. et al. Synthetic evolutionary origin of a proofreading reverse transcriptase. Science 352, 1590–1593 (2016).
Skasko, M. et al. Mechanistic differences in RNA-dependent DNA polymerization and fidelity between murine leukemia virus and HIV-1 reverse transcriptases. J. Biol. Chem. 280, 12190–12200 (2005).
Mathews, C. K. Deoxyribonucleotide metabolism, mutagenesis and cancer. Nat. Rev. Cancer 15, 528–539 (2015).
Wu, W.-J., Yang, W. & Tsai, M.-D. How DNA polymerases catalyse replication and repair with contrasting fidelity. Nat. Rev. Chem. 1, 0068 (2017).
Truniger, V., Lázaro, J. M. & Salas, M. Two positively charged residues of phi29 DNA polymerase, conserved in protein-primed DNA polymerases, are involved in stabilisation of the incoming nucleotide. J. Mol. Biol. 335, 481–494 (2004).
Halperin, S. O. et al. CRISPR-guided DNA polymerases enable diversification of all nucleotides in a tunable window. Nature 560, 248–252 (2018).
Tou, C. J., Schaffer, D. V. & Dueber, J. E. Targeted diversification in the S. cerevisiae genome with CRISPR-guided DNA polymerase I. ACS Synth. Biol. 9, 1911–1916 (2020).
Grünewald, J. et al. Engineered CRISPR prime editors with compact, untethered reverse transcriptases. Nat. Biotechnol. 41, 337–343 (2022).
Ohtsubo, Y., Sasaki, H., Nagata, Y. & Tsuda, M. Optimization of single strand DNA incorporation reaction by Moloney murine leukaemia virus reverse transcriptase. DNA Res. 25, 477–487 (2018).
Petri, K. et al. CRISPR prime editing with ribonucleoprotein complexes in zebrafish and primary human cells. Nat. Biotechnol. 40, 189–193 (2022).
Wang, Q. et al. Broadening the reach and investigating the potential of prime editors through fully viral gene-deleted adenoviral vector delivery. Nucleic Acids Res. 49, 11986–12001 (2021).
Liu, P. et al. Improved prime editors enable pathogenic allele correction and cancer modelling in adult mice. Nat. Commun. 12, 2121 (2021).
Esteban, J. A., Salas, M. & Blanco, L. Fidelity of phi 29 DNA polymerase. Comparison between protein-primed initiation and DNA polymerization. J. Biol. Chem. 268, 2719–2726 (1993).
Lieberman, K. R. et al. Processive replication of single DNA molecules in a nanopore catalyzed by phi29 DNA polymerase. J. Am. Chem. Soc. 132, 17961–17972 (2010).
Anzalone, A. V. et al. Programmable deletion, replacement, integration and inversion of large DNA sequences with twin prime editing. Nat. Biotechnol. 40, 731–740 (2021).
Blanco, L. et al. Highly efficient DNA synthesis by the phage phi 29 DNA polymerase. Symmetrical mode of DNA replication. J. Biol. Chem. 264, 8935–8940 (1989).
Gillmore, J. D. et al. CRISPR–Cas9 in vivo gene editing for transthyretin amyloidosis. N. Engl. J. Med. 385, 493–502 (2021).
Lee, R. G. et al. Efficacy and safety of an investigational single-course CRISPR base-editing therapy targeting PCSK9 in nonhuman primate and mouse models. Circulation 147, 242–253 (2023).
Hopfner, K.-P. & Hornung, V. Molecular mechanisms and cellular functions of cGAS–STING signalling. Nat. Rev. Mol. Cell Biol. 21, 501–521 (2020).
Jiang, T. et al. Chemical modifications of adenine base editor mRNA and guide RNA expand its application scope. Nat. Commun. 11, 1979 (2020).
Zheng, C. et al. A flexible split prime editor using truncated reverse transcriptase improves dual-AAV delivery in mouse liver. Mol. Ther. 30, 1343–1351 (2022).
Mir, A. et al. Heavily and fully modified RNAs guide efficient SpyCas9-mediated genome editing. Nat. Commun. 9, 2641 (2018).
Liu, B. et al. Targeted genome editing with a DNA-dependent DNA polymerase and exogenous DNA-containing templates. NCBI Bioproject https://www.ncbi.nlm.nih.gov/bioproject/1004245 (2023).
Acknowledgements
We thank S. Wolfe, B. Kelch, S. Liang, P. Liu and members of the laboratories of W.X. and E.J.S. for helpful discussions as well as T. Yu and Z. Weng for help with data analysis. We also thank Ben Kleinstiver for communicating results before publication. W.X. was supported by grants from the National Institutes of Health (DP2HL137167, P01HL158506 and UH3HL147367) and the Cystic Fibrosis Foundation. X.D. and E.J.S. acknowledge support from the Leducq Foundation Transatlantic Network of Excellence Program.
Author information
Authors and Affiliations
Contributions
B.L., X.D., W.X. and E.J.S. conceptualized the project and designed experiments. B.L., X.D., C.Z. and Z.C. conducted molecular biological experiments. D.K. synthesized and purified LPETs and assisted in their design. B.L., X.D., C.Z. and H.C. conducted high-throughput sequencing and bioinformatic analyses. B.L., X.D., J.K.W., W.X. and E.J.S. interpreted the data and wrote the paper, and all authors edited the paper.
Corresponding authors
Ethics declarations
Competing interests
E.J.S. is a co-founder and Scientific Advisory Board member of Intellia Therapeutics and a Scientific Advisory Board member at Tessera Therapeutics. The University of Massachusetts Chan Medical School has filed patent applications related to this work. All other authors have no competing interests.
Peer review
Peer review information
Nature Biotechnology thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 LPET-mediated precision editing.
a, The levels of LPET and in vitro transcribed epegRNA were measured by qPCR after electroporation in 293T cells (n = 3). b, Diagram of LPET, MS2-less LPET, ssDNA, modified petRNA (LPET, −17), and unmodified petRNA. c, LPET and no-RT controls. HEK293T cells were electroporated with indicated mRNA (1 μg), sgRNA (100 pmol), nicking sgRNA (100 pmol), and FANCF LPET(+2), PRNP LPET (+0), IDS LPET (+0), RUNX1 LPET (+2), HBB LPET (+2). Editing was measured by deep sequencing (n = 3). Data and error bars indicate mean and s.d. of three independent biological replicates. d, Analysis of precise editing, imprecise editing, and scaffold incorporation events (FANCF) by CRISPResso2-prime editing mode (n = 3). 'MODIFIED’ represents reads containing unexpected insertions, deletions, or substitutions. Data and error bars indicate mean and s.d. of three independent biological replicates. e, Representative reads for FANCF and PRNP. Some deletions at FANCF are likely caused by short homology (underlined) near the nicking sites. Top five reads are shown.
Extended Data Fig. 2 MMLV RT-mediated precision editing with different LPET modifications.
a, Various modifications in the LPET. Precision editing by LPET with additional chemically modified residues [including Locked Nucleic Acid (LNA), 2′-F, 2′-O-methyl (OMe), 2′-fluoro (F) and 3′ phosphorothioate (PS)] as measured by deep sequencing in 293T cells (n = 3). * = 3′ PS linker. All sequences are written from 5′ to 3′. Data and error bars indicate the mean and s.d. of three independent biological replicates. b, RT mediates precise editing by LPETs with an all-DNA RTT and PBS with no modifications, or with the indicated 3′-terminal modifications, as measured by deep sequencing (n = 3). Blue and purple letters denote DNA, RNA and 2′-O-methyl RNA, respectively. Data and error bars indicate the mean and s.d. of three independent biological replicates.
Extended Data Fig. 3 Phi29 with unmodified and modified DPETs and additional controls.
a, Precise editing mediated by Phi29. HEK293T cells were electroporated with indicated mRNA (1 μg), sgRNA (100 pmol), nicking sgRNA (100 pmol), and indicated DPET(+0), MS2-less DPET, MS2-less ssDNA (RTT+PBS), modified linear DPET(−17)), or un-modified (all-RNA) DPET. b, Precise editing mediated by Phi29 and pegRNA. Editing efficiency was measured by deep sequencing (n = 3). Data and error bars indicate the mean and s.d. of three independent biological replicates; two-tailed unpaired Student’s t-test; **P < 0.01.
Extended Data Fig. 4 Phi29 DPE with different DPET modifications.
a, Phi29-mediated precision editing with various modifications in the DPET (including locked nucleic acid, 2′-fluoro, 2′-O-methyl, and 3′ phosphorothioate) as measured by deep sequencing (n = 3). * = 3′ PS linker. All sequences are written from 5′ to 3′. Data and error bars indicate the mean and s.d. of three independent biological replicates. b, Phi29 mediates precise editing by DPETs with an all-DNA RTT and PBS with no modifications, or with the indicated 3′-terminal modifications, as measured by deep sequencing (n = 3). Blue and purple letters denote DNA, RNA and 2′-O-methyl RNA residues, respectively. Data and error bars indicate the mean and s.d. of three independent biological replicates.
Extended Data Fig. 5 Incremental DNA replacement in the PBS/RTT at the PRNP site.
Precision editing with DNA replacement in the PBS/RTT with LPET/RT or DPET/Phi29. Data and error bars indicate the mean and s.d. of three independent biological replicates.
Extended Data Fig. 6 LPETs and DPETs enable precise genome editing without an additional nicking sgRNA in multiple endogenous sites.
a–c, HEK293T cells were electroporated with indicated mRNAs (1 μg), pegRNA (100 pmol, PE2) and nicking sgRNA (100 pmol, PE3). LPET and DPET groups include mRNAs, sgRNA and nicking sgRNA (100 pmol), and LPET/DPET (FANCF +2 (a), PRNP +0 (b), RUNX1 +2 (c)). Data and error bars indicate the mean and s.d. of three independent biological replicates.
Extended Data Fig. 7 LPETs and DPETs enable precise genome editing.
a–c, LPET/RT and DPET/Phi29 editing in multiple cell lines, as measured by deep sequencing (n = 3). ND, not determined. ‘SNP’ indicates the existence of a SNP in the HBB target region in the U2OS cell line, precluding comparative testing. d,e, Representative FACS plots in Fig. 2f. f, Diagram depicting mSA-RT and Phi29. The 5′-biotinylated LPET or DPET is recruited by mSA and anneals to the nicked DNA. g, Tests of biotinylated-LPETs and biotinylated-DPETs in an mCherry reporter line and at endogenous sites (n = 3). Data and error bars indicate the mean and s.d. of three independent biological replicates.
Extended Data Fig. 8 PoII5M DNA polymerase, MarathonRT and TGIRT editing with synthetic templates.
a, Diagram depicting PoII5M/DPET editing (left). MCP-tethered PoII5M binds to the DPET via MS2. The DPET anneals to the nicked DNA strand and serves as the template for PoII5M DNA polymerase. Data indicate precision editing (3-nt substitution) at the FANCF locus with DPET (+2) and fully 2′-O-methyl (OMe)-substituted PBS (right). b, Precise editing at the FANCF locus with MarathonRT and TGIRT. Editing efficiency was measured by deep sequencing on day 3 (n = 3). Data and error bars indicate the mean and s.d. of three independent biological replicates.
Extended Data Fig. 9 Off-target and cytotoxicity evaluation.
a, On-target and off-target editing efficiencies for FANCF. Data indicate the mean and standard deviation of three independent biological replicates. b, HEK293T cells were electroporated with indicated mRNA reagents or buffers. Cell viability was measured by the CellTiter-Glo assay (n = 3) on the indicated time points. Data and error bars indicate mean and s.d. of three independent biological replicates. Two-tailed unpaired Student’s t-test: *, P < 0.05.
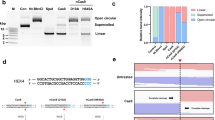
Extended Data Fig. 10 Precise editing for a 40-bp replacement and a 132-bp insertion.
a, Precise editing for a 40-bp insertion and 90-bp deletion at two AAVS sites (1615 and 1705) using LPET/DPET through mRNA nucleofection, followed by PCR amplification and agarose gel electrophoresis. b, Quantification of 40-bp insertion in a by deep sequencing (n = 3). Data and error bars indicate the mean and s.d. of three independent biological replicates, unless otherwise indicated. c, Diagram of the TwinPE by LPETs/DPETs for long insertion. d, Precise editing for a 132-bp insertion at the HEK3 site. Editing efficiency was measured by ddPCR (n = 2).
Supplementary information
Supplementary Information
Supplementary Table 1. Synthetic sgRNAs, nicking sgRNAs and pegRNAs. Supplementary Table 2. Synthetic LPETs and DPETs. Supplementary Table 3. Primers used for high-throughput sequencing. Supplementary Table 4. Primers used for ddPCR. Supplementary Table 5. Primers used for qPCR. Supplementary sequences.
Source Data Extended Data Fig. 10
Unmodified gel
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Liu, B., Dong, X., Zheng, C. et al. Targeted genome editing with a DNA-dependent DNA polymerase and exogenous DNA-containing templates. Nat Biotechnol (2023). https://doi.org/10.1038/s41587-023-01947-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41587-023-01947-w
This article is cited by
-
Targeted genome-modification tools and their advanced applications in crop breeding
Nature Reviews Genetics (2024)
-
Targeted genome editing with a DNA-dependent DNA polymerase and exogenous DNA-containing templates
Nature Biotechnology (2023)

{kind=link}