
Abstract
Arising from: S. J. Lolle, J. L. Victor, J. M. Young & R. E. Pruitt Nature 434, 505–509 (2005); see also communication from Ray; Lolle et al. reply.
Lolle et al. suggest that non-mendelian inheritance in Arabidopsis thaliana might be attributable to an ancestral RNA-sequence cache1, whereby the RNA genome of previous generations causes a high rate of reversion of the plant's mutant hothead (hth) and erecta (er) genes. Here I describe a ‘distributed genome’ model that also explains their results, in which mutant hth DNA is restored by homologous sequences present in the genome itself. This model has implications for the generation of diversity without mating.
Similar content being viewed by others

Main
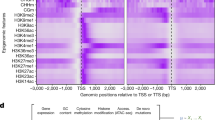
DNA-homology searches of the Arabidopsis genome based on the 21 nucleotides surrounding hth-4, hth-8, hth-10 and er reveal the presence of short, perfectly homologous DNA stretches (known as ‘reverting sequences’) that include nucleotides needed to correct these mutations (Fig. 1). There are also many examples of short homology in genes tested for polymorphism (including GL1, UFO and GAPC).

Sequences of the mutants are shown in blue, with the mutated nucleotide in lower-case; the corresponding wild-type sequences are in black and the nucleotide at the site of mutation is highlighted in red. Homologous sequences that might cause the mutations to revert (RS sequences), obtained by BLAST-searching the Landsberg erecta database from www.arabidopsis.org, are shown in green, with the wild-type nucleotide in red. Reversion frequency is lower for the hth-4 allele (with 6 RS sequences of 13–15 nucleotides), than for hth-8 (with 20 RS of 13–15 nucleotides) and hth-10 (with 24 RS and 13–18 nucleotides).
These sequences might be transcribed into short RNA molecules directed against other chromosomal loci by the cellular machinery, perhaps with the involvement of DRD1 (ref. 2), producing short RNA–DNA hybrids with potential mismatches that can be corrected by mismatch repair3. Consistent with this model, the hth-4 allele — with 6 reverting sequences of 13–15 nucleotides each — has a lower reversion frequency than hth-10, which has 24 reverting sequences of 13–18 nucleotides. As a result of such differences in number, as well as differences in the production of short RNA, some sequences might be changed more than others.
These short sequences should also produce forward mutations, so it is important to measure forward-mutation frequency for several loci. Also, of the several short sequences available for reversion, some might express more short RNA in the male gamete, explaining the preferential transmission of reverted alleles through pollen. The messenger RNA of the corresponding gene may competitively hybridize with a small length of RNA and prevent its interaction with DNA but, owing to the shortness of the base-paired sequence, the mRNA would not be totally inactivated and so would not produce a mutant phenotype. As both nonsense and missense alleles can cause a reduction in transcription compared with the wild-type allele4, there would be more opportunity for reversion to wild type than for forward mutation, which would account for the shielding of the genome against forward mutations.
Conversion to neutral alleles could also occur by this mechanism, but again some sequence stretches might be more effective than others. Lolle et al. did not find any neutral mutation in nine reverted HTH genes1. Even if the activity of all sequence stretches were comparable, the active reversion at any site should be independent of events at other sites: therefore, the frequency of a neutral mutation among revertants is expected to be around 1%, lower than the level of detection in Lolle et al.1.
The proposed sequence-mediated reversion frequency could be boosted by another mechanism not described by Lolle et al.1. The hth mutant cuticles have increased cellular permeability compared with the wild type5. I suggest that the hth embryo sac is also more porous than the HTH embryo sac, causing DNA in the HTH/hth heterozygote to enter the hth embryo sac from the two degraded HTH spores and become ‘archived’, as in the P22 phage6,7. Although the increased cellular permeability would allow the hth gametophyte to obtain HTH molecules in the F1 generation, in subsequent generations these DNAs need not replicate; the endogenous reverting sequence might provide a basal level of reversion.
Lolle et al. propose that metabolic stress in hth increases its reversion frequency1, which might increase information transfer between selected short sequences to alter DNA and create genetic diversity.
References
Lolle, S. J., Victor, J. L., Young, J. M. & Pruitt, R. E. Nature 434, 505–509 (2005).
Matzke, M. A. & Birchler, J. A. Nature 455, 24–34 (2005).
Watson, J. D., Losick, R., Bell, S. P., Gann, A. & Levine, M. Molecular Biology of the Gene (Cummings, San Francisco, 2003).
Mustajoki, S., Kauppinen, R., Mustajoki, P., Suomalainen, A. & Peltonen, L. Genome Res. 7, 1054–1060 (1997).
Lolle, S. J., Hsu, W. & Pruitt, R. E. Genetics 149, 607–619 (1998).
Downs, D. M. & Roth, J. R. Genetics 117, 367–380 (1987).
Ray, A. Nature doi:10.1038/nature04063 (2005).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Chaudhury, A. Hothead healer and extragenomic information. Nature 437, E1 (2005). https://doi.org/10.1038/nature04062
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature04062
This article is cited by
-
RNA cache or genome trash?
Nature (2005)
-
Hothead healer and extragenomic information (reply)
Nature (2005)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
