
Abstract
Early symptoms of food allergy, including diarrhea, are caused by IgE-mediated anaphylactic reactions. To clarify the mechanisms of IgE-mediated anaphylactic reactions in the intestine induced by orally administered antigen, a mouse model was established by s.c. implantation of a murine hybridoma capable of producing monoclonal anti-trinitrophenyl IgE antibody. Morphologic and immunologic changes in the intestine, as well as the effect of the soluble high affinity IgE receptor α chain, were investigated after oral challenge with antigen in this mouse model. Diarrhea, a decrease in s.c. blood flow, an increase in vascular permeability, a substantial increase in serum histamine levels, and noticeable infiltration of mast cells and IgE-bearing cells into the lamina propria were observed around 30 min after antigen challenge. However, these changes were efficiently prevented by pretreatment of the mice with the soluble high affinity IgE receptor α chain. These findings suggested that oral administration of antigen actually induced anaphylactic shock in our mouse model. This reaction was most likely to be mediated by mast cell activation, in response to the IgE-antigen complex, and a soluble form of the high-affinity IgE receptor efficiently prevented this IgE-mediated anaphylactic reaction by trapping free IgE.
Similar content being viewed by others


Main
Diarrhea, as well as eczema and respiratory distress, are often seen in infants and young children as the symptoms of food allergy provoked by ingested foods. These symptoms, observed within minutes to an hour after antigen ingestion, are caused by antigen specific IgE-mediated responses. Intestinal biopsy specimens reveal mucosal edema and mast cell infiltration(1,2); however, the precise mechanisms of these reactions have not been well elucidated. Several animal models of immediate-type allergy were established to investigate allergic responses(3,4). Most of these models were established by active immunization of animals with certain antigen-adjuvant complexes, and as a result not only IgE, but also substantial amounts of IgG1 and IgA were produced in these models(4). Elevation of IgG1 and IgA is also seen in patients suffering from allergic diseases and these models are considered to be suitable for analyzing allergic reactions in vivo. However, they are not appropriate models in which to analyze IgE-dependent reactions in the intestine. This is because IgG1 can directly contribute to anaphylaxis(5) and IgA could protect the host from further antigen invasion into the mucosa; as a result these immunoglobulins could interfere with IgE-dependent reactions.
Multiple steps can be taken in medical intervention against allergic reactions after the immunologic mechanisms. One potentially effective approach is to inhibit IgE-binding to FcεRI on mast cells to abrogate mast cell activation(6,7). It has been demonstrated that the soluble form of human FcεRIα chain (sFcεRIα) inhibits sensitization of mast cells and basophils to IgE in vitro by capturing free IgE(6–8) and, furthermore, suppresses the allergic response in passively sensitized mice as well as rats(7,9,10). However, the effect of sFcεRIα on food allergy has not been investigated.
In this study, to clarify the mechanisms of IgE-dependent reactions in the intestine induced by orally administered allergens, we established a mouse model expressing high levels of antigen-specific IgE by s.c. implantation of an IgE-producing hybridoma into syngeneic mice. We then investigated the morphologic and immunologic changes in the intestinal mucosa after oral antigen challenge. Because repeated allergic responses can be elicited in these mice(9), we also examined the therapeutic effects of sFcεRIα on recurrent allergic changes in the intestine.
METHODS
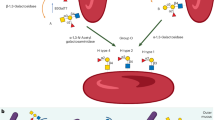
Mice. Female BALB/c mice aged 8-10 wk were purchased from Clea Japan, Inc. (Tokyo). Three groups, consisting of 10 mice each, of hybridoma nonbearing control mice, hybridoma-bearing mice, and sFcεRIα pretreated hybridoma-bearing mice were prepared. Fifty milligrams of TNP37-HSA dissolved in 1 mL of PBS (the volume required to fill up the stomach) was administered into the stomach of each mouse by feeding tube, and the morphologic and immunologic changes were examined in each group (Fig. 1a).

Protocol for experients. (a) Induction of anaphylactic shock in a mouse model (hybridoma-bearing mouse). (b) Prevention of anaphylactic shock in the hybridoma-bearing mouse by treatment with soluble high affinity receptor for IgE (sFcεRIα).
Establishment of the mouse model. Murine hybridoma, IGEL a2 cells (ATCC, TIB 142) secreting anti-TNP IgE(11), were maintained in Dulbecco's minimum essential medium (Nissui Seiyaku Co. Ltd., Tokyo, Japan) supplemented with 10% fetal bovine serum, 10 mM N-2 hydroxyethylpiperazine-N′-2-ethanesulfonic acid, 2 mM glutamine, 50 U/mL penicillin, and 50 µg/mL streptomycin. After washing with Hanks' (Nissui Seiyaku Co. Ltd., Tokyo, Japan), IGEL a2 cells, at a dose of 1 × 107 cells/mouse, were s.c. injected at a site on the back of the syngeneic BALB/c mice. Blood samples were obtained from the orbital sinuses, and the concentration of serum anti-TNP IgE was quantified by ELISA as described previously(10). Serum anti-TNP IgE increased to a level higher than 600 ng/mL around 10 d after inoculation of the mice with these cells.
Production and purification of human sFcεRIα. Chinese hamster ovary cell clones transfected with cDNA encoding human sFcεRIα (constructed by the polymerase chain reaction method) were established(7,9). The sFcεRIα was purified from the culture supernatant of these Chinese hamster ovary cell clones by affinity chromatography using a mouse IgE-AffiGel-10 column, followed by gel filtration through a Sephacryl S-200 (Pharmacia Fine Chemicals, Uppsala, Sweden) column, and extensively dialyzed against PBS. To assess the effect of sFcεRIα on the intestinal anaphylactic reactions, 50 µg of sFcεRIα per mouse was administered i.v. 1 d before the first antigen challenge and periodically between the first and the second antigen challenge, as shown in Figure 1b. The amount of sFcεRIα was apparently in molar excess of the IgE present in the hybridoma-bearing mice.
Histologic evaluation of the intestinal mucosa. More than 5 jejunum samples (each approximately 1 cm long), taken from a location 10 cm toward the anal side from the pylorus, were obtained from each mouse and washed gently with PBS. For evaluation of morphologic changes, each sample was fixed with 10% formalin, and stained with HE solution. To confirm mast cell infiltration, other samples were fixed with Carnoy's solution and stained with Alcian blue and Safranin(12). More than 10 villi were examined in each section and the number of mast cells per 1 mm(2) was determined.
Measurement of s.c. blood flow. S.c. blood flow in the mouse tail was monitored by laser Doppler blood flowmeter (LBF-III, Biomedical Science Co. Ltd., Ishikawa, Japan) that consists of a helium-neon laser, an electronic circuit, and a pair of fiber-optic light guides that conduct laser light to the tissue and carry the backscattered light to a photodetector. Blood flow was measured 30 min after antigen challenge by detecting the mean Doppler frequency of scattered light produced by moving red blood cells(13). Percent blood flow was calculated as: 100 × (blood flow 30 min after antigen challenge)/(blood flow before antigen challenge).
Evaluation of vascular permeability in the intestinal mucosa. Mice were i.v. injected with 0.5% Evans blue (0.5 mL) just before oral antigen challenge and were killed 30 min after the challenge. The jejunum was removed from each animal, cut longitudinally, and dye extravasation from blood vessels of the intestinal mucosa was examined.
Immunohistochemistry of the intestinal mucosa. The freshly obtained jejunal mucosa were rapidly frozen in OCT embedding medium using liquid nitrogen-cooled isopentane and stored at -80°C until processing. Frozen sections (4 µm in thickness) were cut and air dried before further processing. The sections were fixed with cold acetone for 10 min, and after blocking with normal goat serum, were incubated with rat anti-mouse IgE MAb (PharMingen, San Diego, CA). After washing, the sections were incubated with FITC-conjugated goat anti-rat immunoglobulin antibody to detect IgE-bearing cells. Nonspecific staining was evaluated by the sections stained with 0.02% normal rat serum instead of the first antibody.
Determination of plasma histamine levels. Blood samples were obtained 30 min after antigen challenge from the hybridoma-bearing mice with or without sFcεRIα treatment. The blood samples were collected in plastic tubes containing EDTA and immediately placed on ice. Then, the samples were centrifugated at 4°C for 10 min at 2500 × g, and the plasma was carefully pipetted into polypropylene tubes. Plasma histamine concentration was measured by a radioenzymatic assay following the method of Verburg et al.(14) using the kit supplied by Du Pont de Nemours & Co. (NEN Products, Boston, MA). All samples were assayed in duplicate.
Results are expressed as mean ± SD. Statistical significance was evaluated by the Mann-Whitney U test.
RESULTS
TNP37-HSA was orally administered to evaluate the symptoms of anaphylactic reactions in hybridoma-bearing mice displaying serum anti-TNP IgE levels of more than 600 ng/mL. Diarrhea, anal mucosal edema, and hyperemia were observed around 30 min after antigen challenge in the hybridoma-bearing mice, whereas no remarkable changes were observed in the hybridoma nonbearing control mice. Dilatation of the lymph ducts was also observed in the jejunal mucosa of the hybridoma-bearing mice after antigen challenge, as confirmed by HE staining. These responses decayed within 2 h. The inhibitory effect of the sFcεRIα was examined in hybridoma-bearing mice administered 50 µg of sFcεRIα i.v. before antigen challenge. All symptoms, including diarrhea, anal mucosal edema, and hyperemia were prevented after the second antigen challenge in these hybridoma-bearing mice continuously treated with sFcεRIα.
Alcian blue and Safranin staining was carried out to examine mast cell infiltration into the intestinal mucosa. Mucosal-type mast cells, containing three to six granules stained with Alcian-blue dye, were found to have infiltrated into the lamina propria of the hybridoma-bearing mice after antigen challenge. The number of infiltrating mast cells was significantly increased in the hybridoma-bearing mice compared with either that in hybridoma nonbearing control mice 30 min after antigen challenge or that in hybridoma-bearing mice before antigen challenge (p < 0.01 in both). The number of mast cells in sFcεRIα pretreated hybridoma-bearing mice after the second antigen challenge was still significantly elevated compared with that in hybridoma nonbearing control mice (p < 0.05), thus the number was relatively decreased compared with that in hybridoma-bearing mice without sFcεRIα treatment, as shown in Figure 2.

The number of mast cells per mm2 in jejunal mucosa after oral antigen challenge in hybridoma nonbearing control mice, hybridoma-bearing mice, and hybridoma-bearing mice treated with sFcεRIα at a dose of 50 µg/mouse are shown. Data are presented as the mean ± SD of 10 animals for each group (*p < 0.05, ** p < 0.01). Similar results were obtained in three independent experiments.
Subcutaneous blood flow in the mouse tail was monitored with a laser-Doppler blood flowmeter to evaluate systemic changes, as shown in Figure 3. The blood flow was significantly decreased to approximately 30% of the prechallenged level in the hybridoma-bearing mice compared with that in hybridoma nonbearing control mice approximately 30 min after antigen challenge (p < 0.01). The blood flow also decreased after the first antigen challenge, even in the sFcεRIα treated hybridoma-bearing mice, showing a response pattern similar to that observed in the untreated mice. However, the responses of the treated and untreated mice were different after the second antigen challenge. The decrease in blood flow was not significant after the second antigen challenge in the sFcεRIα treated mice, whereas the blood flow dropped in the untreated mice as observed after the first antigen challenge, as shown in Figure 3.

Subcutaneous blood flow in the mouse tail was monitored using a laser-Doppler blood flowmeter and systemic changes in the hybridoma non-bearing control mice, hybridoma-bearing mice, and hybridoma-bearing mice treated with sFcεRIα at a dose of 50 µg/mouse are shown. Blood flow was measured 30 min after oral antigen challenge. Data are presented as the mean ± SD of 10 animals for each group (* p < 0.01). Similar results were obtained in three independent experiments.
An increase in vascular permeability caused by IgE-mediated anaphylaxis in the intestine was confirmed by monitoring dye extravasation. Figure 4a shows the jejunal mucosa from a hybridoma nonbearing control mouse, Figure 4b shows that from a hybridoma-bearing mouse 30 min after antigen challenge, and Figure 4c shows that from a hybridoma-bearing mouse treated with sFcεRIα continuously. Extensive dye extravasation was observed in the hybridoma-bearing mouse but not in the control mouse after antigen challenge, mainly in the upper jejunum rather than the lower jejunum or ileum. Interestingly, the increase in vascular permeability was efficiently prevented after the second antigen challenge in the group treated with sFcεRIα (Fig. 4c).

Vascular permeability in the intestinal mucosa was examined by dye extravasation. (a) Jejunum of a hybridoma nonbearing control mouse 30 min after antigen challenge. (b) That of a hybridoma-bearing mouse. (c) That of a sFcεRIα pretreated hybridoma-bearing mouse. Representative samples from 10 mice each are presented.
To prove that sFcεRIα is effective to prevent binding of antigen-IgE complex to Fc receptors on the surface of mast cells, IgE-bearing cells in the jejunal mucosa of treated and untreated mice were stained with a monoclonal anti-mouse IgE antibody after the second antigen challenge. There were a few cells stained with this antibody in the intestinal mucosa of the hybridoma-bearing mice before antigen challenge (Fig. 5a). Many cells in the lamina propria of the untreated hybridoma-bearing mice were brightly stained with the anti-IgE antibody (Fig. 5b). In contrast, treatment of these mice with sFcεRIα notably decreased the number of IgE-bearing cells in the jejunal mucosa, as shown in Figure 5c.

IgE-bearing cells in the jejunal mucosa of hybridoma-bearing mice were stained with a monoclonal anti-mouse IgE antibody. (a) The jejunal mucosa of an untreated mouse before antigen challenge. (b) That of an untreated mouse 30 min after antigen challenge. (c) The jejunal mucosa of a sFcεRIα treated hybridoma-bearing mouse 30 min after antigen challenge. Representative samples from 10 mice each are presented.
The effect of sFcεRIα on the intestinal allergic response was further investigated by monitoring serum histamine levels (Fig. 6). In the hybridoma-bearing mice, serum histamine levels were significantly elevated 30 min after antigen challenge compared with that of hybridoma nonbearing control mice after antigen challenge (p < 0.01). The increase in serum histamine levels after the second antigen challenge was remarkably prevented by treatment of the hybridoma-bearing mice with sFcεRIα (p < 0.01).

Serum histamine levels after oral antigen challenge in hybridoma nonbearing control mice, hybridoma-bearing mice, and hybridoma-bearing mice treated with sFcεRIα at a dose of 50 µg/mouse are shown. Blood samples were collected 30 min after oral antigen challenge. Data are presented as the mean ± SD of 10 animals for each group (* p < 0.05, ** p < 0.01). Similar results were obtained in three independent experiments.
DISCUSSION
Animals actively immunized with certain antigens and adjuvants have been adopted for investigating anaphylactic reactions(3,4). Consequently, mixed reactions induced by IgE and IgG1 have been observed in these animal models(4), hence the IgE-dependent reactions, without any interference of IgG1 and/or IgA-mediated reactions, could not be identified. To clarify the mechanisms of IgE-dependent reactions in the intestine, we established a mouse model with high serum levels of antigen-specific IgE by s.c. implantation of IGEL a2 hybridoma. This model is limited because there is no increase in antigen-specific IgG1 and IgA; therefore, it does not fully reproduce the intricacies of the naturally occurring IgE-mediated allergic responses. However, the findings of diarrhea, mucosal hyperemia, decreased s.c. blood flow (Fig. 3), and increase in vascular permeability in the intestine (Fig. 4) upon oral antigen challenge suggested that anaphylactic reactions were induced in these mice.
A remarkable increase in number of IgE-bearing cells in the intestinal mucosa and submucosa in response to oral antigen challenge was observed in our mouse model (Fig. 5). Most of these IgE-bearing cells could be mast cells, as reported in the case of human intestine(15), because we actually confirmed the infiltration of mast cells into the intestinal mucosa of these mice (Fig. 2). The specific IgE bound to FcεRI on the surface of mast cells becomes cross-linked upon exposure to allergens, and intracellular signals are transduced to activate the mast cells to release chemical mediators like histamine, leukotriene C4, TNF-α, IL-4, and other cytokines(16,17). To prove that the mast cells contribute to anaphylactic reactions in these mice, histamine was examined and significant elevation of serum histamine levels was detected in the hybridoma-bearing mice after antigen challenge (Fig. 6). It is very likely that these mediators induce a local increase in vascular permeability, lymph duct dilatation, and increased mucus production, and elicit watery diarrhea, resulting in blood hypocirculation in these mice. Taken together, these histologic and immunologic changes suggest that the mast cells that have infiltrated the intestinal mucosa are deeply involved in the anaphylactic reactions induced by oral antigen challenge.
Kinet et al.(18) generated FcεRIα chain-deficient mice and demonstrated that FcεRI is essential for initiation of the IgE-mediated anaphylactic reaction. Therefore, interfering with FcεRI function should be an effective means of treating anaphylactic-type allergy. sFcεRIα has been shown to bind IgE with affinity similar to that of native FcεRI and has been extensively studied as a competitive inhibitor of IgE binding to mast cells in vitro and in vivo(6–8). sFcεRIα completely inhibits the PCA reaction when administered before sensitization, and sFcεRIα partially blocks the PCA reaction(7) and anaphylactic shock(9) when it is injected after the IgE-sensitization. However, these anaphylactic signs could not be prevented with sFcεRIα after the first antigen challenge because some IgE had already attached to the IgE receptor on the surface of the mast cells before sFcεRIα administration, and these captured antigen-stimulated mast cells at the first challenge. After degranulation, the antigen/IgE/FcεRI complexes on the mast cell surface are internalized and degraded, then these cells synthesize and express new FcεRI(19). Hence, repeated administration of sFcεRIα to mice results in the capture of free IgE in body fluids and prevents IgE binding to unoccupied newly introduced IgE receptors on the mast cell surface. Thus, a prolonged administration of sFcεRIα successfully prevented the development of antigen induced anaphylaxis, as it has been demonstrated in the second antigen challenge of the sFcεRIα pretreated hybridoma-bearing mouse model.
The observed decrease in s.c. blood flow, diarrhea, anal mucosal edema, hyperemia, and dye extravasation after the second antigen challenge were prevented in the mice treated with sFcεRIα. Although the number of mast cells was not significantly decreased (Fig. 2), the number of IgE-bearing cells in the intestinal mucosa of the sFcεRIα pretreated mice was substantially decreased in comparison with the untreated mice (Fig. 5). These results indicate that treatment of these mice with sFcεRIα actually blocked the binding of IgE to the IgE receptors on the surface of mast cells in the intestinal mucosa and thus could prevent further reactions. Allergic patients suffer from frequent IgE-mediated responses as a result of repeated antigen exposure, and our findings suggest that these allergic reactions could be completely blocked by continuous administration of sFcεRIα because it can efficiently trap free IgE in the vicinity of mast cells before binding of the IgE to the cell surface FcεRI.
In this study, we demonstrated that oral administration of antigen actually induced anaphylactic shock in mice displaying high serum levels of antigen-specific IgE. We also proved that a soluble form of the high-affinity IgE receptor successfully prevented this repeated anaphylactic reaction. These findings strongly suggested that mast cells, activated by IgE and allergen complexes, play a prominent role in intestinal allergic reactions induced by oral antigen challenge.
Abbreviations
- FcεRI :
-
high affinity IgE receptor
- HSA :
-
human serum albumin
- PCA :
-
passive cutaneous anaphylaxis
- sFcεRIα :
-
soluble high affinity IgE receptor α-chain
- TNP :
-
trinitrophenyl
References
Kosnai I, Kuitunen P, Savilahti E, Sipponen P 1984 Mast cells and eosinophils in the jejunal mucosa of patients with intestinal cow's milk allergy and celiac disease of childhood. J Pediatr Gastroenterol Nutr 3: 368–372.
Perdue MH, Chung M, Gall DG 1984 Effect of intestinal anaphylaxis on gut function in the rat. Gastroenterology 86: 391–397.
Gall DG, Perdue MH 1986 Experimental models of intestinal allergy. Front Gastrointest Res 13: 45–54.
Snider DP, Marshall JS, Perdue MH, Liang H 1994 Production of IgE antibody and allergic sensitization of intestinal and peripheral tissues after oral immunization with protein Ag and cholera toxin. J Immunol 153: 647–657.
Oshiba A, Hamelmann E, Takeda K, Bradley KL, Loader JE, Larsen GL, Gelfand EW 1996 Passive transfer of immediate hypersensitivity and airway hyperresponsiveness by allergen-specific immunoglobulin (Ig)E and IgG1 in mice. J Clin Invest 97: 1398–1408.
Blank U, Ra C, Kinet JP 1991 Characterization of truncated chain products from human, rat, and mouse high affinity receptor for immunoglobulin. Eur J Biol Chem 266: 2639–2646.
Ra C, Kuromitsu S, Hirose T, Yasuda S, Furuichi K, Okumura K 1993 Soluble human high-affinity receptor for IgE abrogates the IgE-mediated allergic reaction. Int Immunol 5: 47–54.
Yagi S, Yanagida M, Tanida I, Hasegawa A, Okumura K, Ra C 1994 High-level expression of the truncated chain of human high-affinity receptor for IgE as a soluble form by baculovirus-infected insect cells. Biochemical characterization of the recombinant product. Eur J Biochem 220: 593–598.
Naito K, Hirama M, Okumura K, Ra C 1996 Recombinant soluble form of the human high affinity receptor for IgE prevents anaphylactic shock in mice. J Aller Clin Immunol 97: 773–780.
Naito K, Hirama M, Okumura K, Ra C 1995 Soluble form of the human high affinity receptor for IgE inhibits recurrent allergic reaction in a novel mouse model of type I allergy. Eur J Immunol 25: 1631–1637.
Rudolph AK, Burrows D, Wabl MR 1981 Thirteen hybridomas secreting hapten-specific immunoglobulin E from mice with Iga or Igb heavy chain haplotype. Eur J Immunol 11: 527–529.
Strobel S, Ferguson A 1981 Human intestinal mucosal mast cells: evaluation of fixation and staining techniques. J Clin Pathol 34: 851–858.
Shepherd AP, Riedel GL 1982 Continuous measurements of intestinal mucosal blood flow by laser-Doppler velocimetry. Am J Physiol 242:G668–G672.
Verburg KM, Bowsher RR, Henry DP 1988 Quantification of urinary histamine by a new radioenzymatic assay: documentation of assay specificity and establishment of normal excretion rates. J Allergy Clin Immunol 82: 339–347.
Rognum TO, Brandtzaeg P 1992 IgE-positive cells in human intestinal mucosa are mainly mast cells. Int Arch Allergy Appl Immunol 89: 256–260.
Metzger H 1992 The receptor with high affinity for IgE. Immunol Rev 125: 37–48.
Gordon JR, Burd PR, Galli SJ 1990 Mast cells as a source of multifunctional cytokines. Immunol Today 11: 458–464.
Dombrowicz D, Flamand V, Brigman KK, Koller BH, Kinet JP 1993 Abolition of anaphylaxis by targeted disruption of the high affinity immunoglobulin E receptor chain gene. Cell 75: 969–976.
Furuichi K, Rivera J, Buonocore LM, Isersky C 1986 Recycling of receptor-bound IgE by rat basophilic leukemia cells. J Immunol 136: 1015–1022.
Author information
Authors and Affiliations
Additional information
Supported by grants from Morinaga Milk Industry Co, Ltd., Kanagawa, Japan and by a Grant-in-Aid for Scientific Research, The Ministry of Education, Science, Sports and Culture, Japan.
Rights and permissions
About this article
Cite this article
Ohtsuka, Y., Naito, K., Yamashiro, Y. et al. Induction of Anaphylaxis in Mouse Intestine by Orally Administered Antigen and its Prevention with Soluble High Affinity Receptor for IgE. Pediatr Res 45, 300–305 (1999). https://doi.org/10.1203/00006450-199903000-00002
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199903000-00002
This article is cited by
-
Abrogation of High-Affinity IgE Receptor-Mediated Mast Cell Activation at the Effector Phase Prevents Contact Hypersensitivity to Oxazolone
Journal of Investigative Dermatology (2010)
