
Abstract
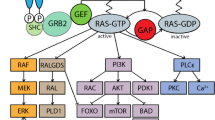
Hypoxia-inducible factor-1alpha (HIF-1α) plays crucial roles in tumor promotion by transactivating approximately 60 kinds of its target genes. Recently, we reported a novel splice variant HIF-1α785, which is regulated primarily by phorbol ester. This variant can be stabilized under normoxic conditions because it loses an acetylation site Lys532. Its expression was found to promote xenografted tumor growth in nude mice. We here found that the Ras oncogene regulates HIF-1α785 expression via the Raf/MEK/ERK pathway, and that both phorbol ester and epidermal growth factor also induced HIF-1α785 via the same pathway. We also identified the nonhypoxic regulatory domain responsible for phorbol ester-induced HIF-1α785 expression. These results imply that HIF-1α785 may play an important role in tumor promotion mediated by the Ras oncogene, phorbol ester or tumor growth factors.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
Alahari SK, Lee JW and Juliano RL . (2000). J. Cell Biol., 151, 1141–1154.
Birner P, Schindl M, Obermair A, Plank C, Breitenecker G and Oberhuber G . (2000). Cancer Res., 60, 4693–4696.
Cho HJ, Jeong HG, Lee JS, Woo ER, Hyun JW, Chung MH and You HJ . (2002). J. Biol. Chem., 277, 19358–19366.
Chun YS, Choi E, Yeo EJ, Lee JH, Kim MS and Park JW . (2001). J. Cell Sci., 114, 4051–4061.
Chun YS, Kim MS and Park JW . (2002). J. Korean Med. Sci., 17, 581–588.
Chun YS, Lee KH, Choi E, Bae SY, Yeo EJ, Huang LE, Kim MS and Park JW . (2003). Cancer Res., 63, 8700–8707.
Feldser D, Agani F, Iyer NV, Pak B, Ferreira G and Semenza GL . (1999). Cancer Res., 59, 3915–3918.
Gothie E, Richard DE, Berra E, Pages G and Pouyssegur J . (2000). J. Biol. Chem., 275, 6922–6927.
Hudson CC, Liu M, Chiang GG, Otterness DM, Loomis DC, Kaper F, Giaccia AJ and Abraham RT . (2002). Mol. Cell. Biol., 22, 7004–7014.
Hur E, Chang KY, Lee E, Lee SK and Park H . (2001). Mol. Pharmacol., 59, 1216–1224.
Jeong JW, Bae MK, Ahn MY, Kim SH, Sohn TK, Bae MH, Yoo MA, Song EJ, Lee KJ and Kim KW . (2002). Cell, 111, 709–720.
Liu XH, Kirschenbaum A, Lu M, Yao S, Dosoretz A, Holland JF and Levine AC . (2002). J. Biol. Chem., 277, 50081–50086.
Rak J, Filmus J, Finkenzeller G, Grugel S, Marme D and Kerbel RS . (1995). Cancer Metastasis Rev., 14, 263–277.
Semenza GL . (1998). Curr. Opin. Genet. Dev., 8, 588–594.
Semenza GL . (2002). Trends Mol. Med., 8, S62–S67.
Shapiro P . (2002). Crit. Rev. Clin. Lab. Sci., 39, 285–330.
Yeo EJ, Chun YS, Cho YS, Kim J, Lee JC, Kim MS and Park JW . (2003). J. Natl. Cancer Inst., 95, 516–525.
Zelzer E, Levy Y, Kahana C, Shilo BZ, Rubinstein M and Cohen B . (1998). EMBO J., 17, 5085–5094.
Zhong H, Chiles K, Feldser D, Laughner E, Hanrahan C, Georgescu MM, Simons JW and Semenza GL . (2000). Cancer Res., 60, 1541–1545.
Zhong H, De Marzo AM, Laughner E, Lim M, Hilton DA, Zagzag D, Buechler P, Isaacs WB, Semenza GL and Simons JW . (1999). Cancer Res., 59, 5830–5835.
Acknowledgements
This work was supported by a grant from the Korea Science and Engineering Foundation (R01-2000-000-00139-0) and by a grant from the Cancer Research Institute Seoul National University (2003).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lim, JH., Lee, ES., You, HJ. et al. Ras-dependent induction of HIF-1α785 via the Raf/MEK/ERK pathway: a novel mechanism of Ras-mediated tumor promotion. Oncogene 23, 9427–9431 (2004). https://doi.org/10.1038/sj.onc.1208003
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.onc.1208003
Keywords
This article is cited by
-
KITENIN promotes aerobic glycolysis through PKM2 induction by upregulating the c-Myc/hnRNPs axis in colorectal cancer
Cell & Bioscience (2023)
-
Molecular convergent and parallel evolution among four high-elevation anuran species from the Tibetan region
BMC Genomics (2020)
-
MEK reduces cancer-specific PpIX accumulation through the RSK-ABCB1 and HIF-1α-FECH axes
Scientific Reports (2020)
-
Involvement of A3 Adenosine Receptor in Neuroblastoma Progression via Modulation of the Hypoxic/Angiogenic Pathway
Journal of Molecular Neuroscience (2019)
-
Klotho negatively regulated aerobic glycolysis in colorectal cancer via ERK/HIF1α axis
Cell Communication and Signaling (2018)
