
Abstract
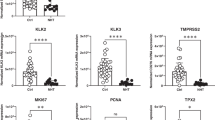
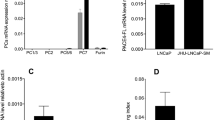
After therapeutic hormone deprivation, most prostate cancer (PrCa) cells develop androgen-independent (AI) growth. PrCa is highly heterogeneous and multifocal, suggesting that several molecular processes or pathways may be contributing to AI. The human LuCaP 23.1 xenograft model retains clinical hallmarks of PrCa, including heterogeneous growth, PSA production, androgen-responsiveness and progression to AI. In this work, we studied the effect of androgen depletion (castration) on the growth of LuCaP 23.1 xenografts. A total of 100 nude mice were implanted and analysed for their growth profiles before and after castration. By 11 and 15 weeks, tumours were harvested and assessed for molecular marker expression specific for PrCa. Prior to castration we found 37 fast growing (FG) tumours (948.9±76.9 mm3) and 63 slow growing (SG) tumours (229.6±18.4 mm3), a previously undescribed result for this PrCa model. Quantitative RT–PCR showed that in comparison to SGs, FGs contained high HER1, uPA and thymidilate synthetase (TS) expression with low levels of 5α-reductase 2 mRNA. All FG tumours progressed rapidly to AI growth 5 weeks after castration (FG-P). In SG castrated tumours, 66% of tumours (SG-P) showed retarded progression (by 12 weeks) to AI, whereas 34% responded to castration (SG-R). Molecular analysis permitted us to define distinct molecular profiles integrating different pathways associated with AI progression. FG-P, and a subgroup of SG-P tumours, presented significantly high levels of peptidylglycine α-amidating monooxygenase (PAM), HER1, HER2, TS, and uPA mRNA, all of which correlated with AR expression. The second subgroup of SG-P tumours showed overexpression of the antiapoptotic gene Bcl-2. A third subgroup of SG-P tumours showed significant expression of hypoxia-related gene (adrenomedullin) after castration. This work permitted to define distinct molecular profiles related to different AI growth in the LuCaP 23.1 xenograft.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Abbreviations
- AI:
-
androgen independent
- AM:
-
adrenomedullin
- FG:
-
fast growing
- FG-P:
-
fast growing/progressing
- 5α-R2:
-
5 alpha-reductase 2
- PAM:
-
peptidyl α-amidating monooxygenase
- PrCa:
-
prostate cancer
- SG:
-
slow growing
- SG-P:
-
slow growing/progressing
- SG-R:
-
slow growing/responding
- TK:
-
thymidine kinase
- TS:
-
thymidilate synthetase
- uPA:
-
urokinase plasminogen activator
References
Abrahamsson PA . (1999). Prostate, 39, 135–148.
Bladou F, Vessella RL, Buhler KR, Ellis WJ, True LD and Lange PH . (1996). Int. J. Cancer, 67, 785–790.
Buttyan R, Ghafar M-A, Shabsigh A, Solit D-B, Zheng F-F, Drobnjak M, Munster P-N, Higgins B, Verbel D, Heller G, Tong W, Cordon-Cardo C, Agus D-B, Scher H-I and Rosen N . (2000). Curr. Opin. Urol., 10, 415–420.
Chomczynski P and Sacchi N . (1987). Anal. Biochem., 162, 156–159.
Craft N, Shostak Y, Carey M and Sawyers CL . (1999). Nat. Med., 5, 280–285.
Culig Z, Hobisch A, Cronauer MV, Radmayr C, Trapman J, Hittmair A, Bartsch G and Klocker H . (1994). Cancer Res., 54, 5474–5478.
Dahiya R, Lee C, Haughney PC, Chui R, Ho R and Deng G . (1996). Urology, 48, 963–970.
Delos S, Carsol JL, Fina F, Raynaud JP and Martin PM . (1998). Int. J. Cancer, 75, 840–846.
Denis L-J and Griffiths K . (2000). Semin. Surg. Oncol., 18, 52–74.
Ellis WJ, Vessella RL, Buhler KR, Bladou F, True LD, Bigler SA, Curtis D and Lange PH . (1996). Clin. Cancer Res., 2, 1039–1048.
Feldman BJ and Feldman D . (2001). Nat. Rev. Cancer, 1, 34–45.
Gaylis FD, Keer HN, Wilson MJ, Kwaan HC, Sinha AA and Kozlowski JM . (1989). J. Urol., 142, 193–198.
Gleave ME, Tolcher A, Miyake H, Beraldi E and Goldie J . (1999). Clin. Cancer Res., 5, 2891–2898.
Helenius MA, Saramäki OR, Linja MJ, Tammel TLJ and Visakorpi T . (2001). Cancer Res., 61, 5340–5344.
Hollas W, Hoosein N, Chung LW, Mazar A, Henkin J, Kariko K, Barnathan ES and Boyd D . (1992). Thromb. Haemost., 68, 662–666.
Huss WJ, Hanrahan CF, Barrios RJ, Simons JW and Greenberg NM . (2001). Cancer Res., 61, 2736–2743.
Janik P, Briand P and Hartman NR . (1975). Cancer Res., 35, 3698–3704.
Jenster G, Denis LJ and Griffiths K . (1999). Semin. Oncol., 26, 407–421.
Joliffe IT and Morgan BJ . (1992). Stat. Methods Med. Res., 1, 69–95.
Koivisto P, Kononen J, Palmberg C, Tammela T, Hyytinen E, Isola J, Trapman J, Cleutjens K, Noordzij A, Visakorpi T and Kallioniemi OP . (1997). Cancer Res., 57, 314–319.
Landström M, Damber JE and Bergh A . (1994). Cancer Res., 54, 4281–4284.
Liu AY, Corey E, Bladou F, Lange PH and Vessella RL . (1996). Int. J. Cancer, 65, 85–89.
Lyon PB, See WA, Xu Y and Cohen MB . (1995). Prostate, 27, 179–186.
McGowen R, Biliran Jr H, Sager R and Sheng S . (2000). Cancer Res., 60, 4771–4778.
McLeod DG . (2003). Urology, 61, 3–7.
Miyake H, Tolcher A and Gleave ME . (1999). Cancer Res., 59, 4030–4034.
Negri-Cesi P, Colciago A, Poletti A and Motta M . (1999). Prostate, 41, 224–232.
Nelson JB and Carducci MA . (2000a). Cancer Invest., 18, 87–96.
Nelson JB and Carducci MA . (2000b). BJU Int., 85 (Suppl 2), 45–48.
Papandreou CN, Usmani B, Geng Y, Bogenrieder T, Freeman R, Wilk S, Finstad CL, Reuter VE, Powell CT, Scheinberg D, Magill C, Scher HI, Albino AP and Nanus DM . (1998). Nat. Med., 4, 50–57.
Parker SL, Tong T, Bolden S and Wingo PA . (1996). Cancer J. Clin., 46, 5–27.
Rocchi P, Boudouresque F, Zamora AJ, Muracciole X, Lechevallier E, Martin PM and Ouafik LH . (2001). Cancer Res., 61, 1196–1206.
Ross RK, Pike MC, Coetzee GA, Reichardt JK, Yu MC, Feigelson H, Stanczyk FZ, Kolonel LN and Henderson BE . (1998). Cancer Res., 58, 4497–4504.
Scher HI . (2000). J. Nalt. Cancer Inst., 92, 1866–1868.
Scher HI, Sarkis A, Reuter V, Cohen D, Netto G, Petrylak D, Lianes P, Fuks Z, Mendelsohn J and Cordon-Cardo C . (1995). Clin. Cancer Res., 1, 545–550.
Solit DB, Zheng FF, Drobnjak M, Munster PN, Higgins B, Verbel D, Heller G, Tong W, Cordon-Cardo C, Agus DB, Scher HI and Rosen N . (2002). Clin Cancer Res., 8, 986–993.
Turkeri LN, Sakr WA, Wykes SM, Grignon DJ, Pontes JE and Macoska JA . (1994). Prostate, 25, 199–205.
Visakorpi T, Hyytinen E, Koivisto P, Tanner M, Keinanen R, Palmberg C., Palotie A, Tammela T, Isola J and Kallioniemi OP . (1995). Nat. Genet., 9, 401–406.
Wen Y, Hu MC, Makino K, Spohn B, Bartholomeusz G, Yan DH and Hung MC . (2000). Cancer Res., 60, 6841–6845.
Xing RH and Rabbani SA . (1999). Endocrinology, 140, 4056–4064.
Yeh S, Lin HK, Kang HY, Thin TH, Lin MF and Chang C . (1999). Proc. Natl. Acad. Sci. USA, 96, 5458–5463.
Zhang L, Johnson M, Le KH, Sato M, Ilagan R, Iyer M, Gambhir SS, Wu L and Carey M . (2003). Cancer Res., 63, 4552–4560.
Zudaire E, Martinez A and Cuttitta F . (2003). Regul. Peptides, 112, 175–183.
Acknowledgements
We thank Dr Robert Vessella's Laboratory, Department of Urology, University of Washington, Seattle for providing with LuCaP 23.1 xenograft. We thank Dr Lisa Valettes, Inserm U419, Nantes, who very kindly carried out Bcl-2 TaqMan analyses.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rocchi, P., Muracciole, X., Fina, F. et al. Molecular analysis integrating different pathways associated with androgen-independent progression in LuCaP 23.1 xenograft. Oncogene 23, 9111–9119 (2004). https://doi.org/10.1038/sj.onc.1208154
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.onc.1208154
Keywords
This article is cited by
-
Targeting TCTP as a New Therapeutic Strategy in Castration-resistant Prostate Cancer
Molecular Therapy (2012)
-
The evolutionary impact of androgen levels on prostate cancer in a multi-scale mathematical model
Biology Direct (2010)
-
Heat shock protein 27 confers resistance to androgen ablation and chemotherapy in prostate cancer cells through eIF4E
Oncogene (2010)
-
Ezrin and alpha-smooth muscle actin are immunohistochemical prognostic markers in conventional osteosarcomas
Virchows Archiv (2007)
-
Identification of genes differentially expressed in glioblastoma versus pilocytic astrocytoma using Suppression Subtractive Hybridization
Oncogene (2006)
