
Abstract
A major challenge in hematopoiesis is to conceive assays that could bring useful insights into experimental and clinical hematology. This means identifying separately the various classes of hematopoietic progenitors that are produced sequentially during the progression from stem cells to differentiated functional cells. Standardized short-term colony assays easily quantify lineage-committed myeloid precursors, but identification of primitive cells, which have both the ability to repopulate durably myeloid and lymphoid lineages and perhaps to self-renew, still depends on in vivo assays. Whatever the assay, two important requisites have to be solved: one is the definition of appropriate read-outs that will depend solely on the function of these cells, and the second is to evaluate precisely their numbers and proliferative potential in quantitative assays. When evaluating hematopoiesis, three parameters have to be taken into account: (1) the lack of reliable correlation between the phenotype of a given cell and its function. This is especially problematic in post-transplantation situations where cells from transplanted animals are analysed; (2) functionally heterogeneous cells are identified in a single assay; and (3) ontogeny-related changes in hematopoietic cell proliferation and self-renewal that, in human beings, hampers the exploration of adult stem cells. Nevertheless, years of progress in the manipulation of hematopoietic stem cells have recently resulted in the purification of a cell subset that repopulates irradiated recipients with absolute efficiency.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
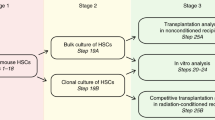

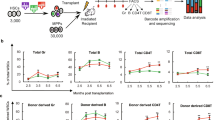
References
Abramson S, Miller RG and Phillips RA . (1977). J. Exp. Med., 145, 1567–1579.
Amsellem S, Pflumio F, Bardinet D, Izac B, Charneau P, Romeo PH, Dubart-Kupperschmitt A and Fichelson S . (2003). Nat. Med., 9, 1423–1427.
Anderson G, Moore NC, Owen JJ and Jenkinson EJ . (1996). Annu. Rev. Immunol., 14, 73–99.
Antonchuk J, Sauvageau G and Humphries RK . (2001). Exp. Hematol., 29, 1125–1134.
Ash RC, Detrick DA and Zanjani ED . (1981). Blood, 58, 309–316.
Barcena A, Galy AH, Punnonen J, Muench MO, Schols D, Roncarolo MG, de Vries JE and Spits H . (1994). J. Exp. Med., 180, 123–132.
Barker J and Verfaillie CM . (2000). Leukemia, 14, 1614–1620.
Baum CM, Weissman IL, Tsukamoto AS, Buckle AM and Peault B . (1992). Proc. Natl. Acad. Sci. USA, 89, 2804–2808.
Bennaceur-Griscelli A, Pondarre C, Schiavon V, Vainchenker W and Coulombel L . (2001). Blood, 97, 435–441.
Benveniste P, Cantin C, Hyam D and Iscove NN . (2003). Nat. Immunol., 4, 708–713.
Berardi AC, Meffre E, Pflumio F, Katz A, Vainchenker W, Schiff C and Coulombel L . (1997). Blood, 89, 3554–3564.
Bhatia M, Wang JC, Kapp U, Bonnet D and Dick JE . (1997). Proc. Natl. Acad. Sci. USA, 94, 5320–5325.
Bonnet D, Bhatia M, Wang JC, Kapp U and Dick JE . (1999). Bone Marrow Transplant., 23, 203–209.
Bonnet D and Dick JE . (1997). Nat. Med., 3, 730–737.
Borge OJ, Ramsfjell V, Cui L and Jacobsen SE . (1997). Blood, 90, 2282–2292.
Breems DA, Blokland EA, Neben S and Ploemacher RE . (1994). Leukemia, 8, 1095–1104.
Breems DA, van Hennik PB, Kusadasi N, Boudewijn A, Cornelissen JJ, Sonneveld P and Ploemacher RE . (1996). Blood, 87, 5370–5378.
Broxmeyer HE . (1984). Crit. Rev. Oncol. Hematol., 1, 227–257.
Brummendorf TH, Dragowska W, Zijlmans J, Thornbury G and Lansdorp PM . (1998). J. Exp. Med., 188, 1117–1124.
Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, Martin RP, Schipani E, Divieti P, Bringhurst FR, Milner LA, Kronenberg HM and Scadden DT . (2003). Nature, 425, 841–846.
Cashman J, Bockhold K, Hogge DE, Eaves AC and Eaves CJ . (1997). Br. J. Haematol., 98, 1026–1036.
Cashman JD and Eaves CJ . (2000). Blood, 96, 3979–3981.
Chang J, Coutinho L, Morgenstern G, Scarffe JH, Deakin D, Harrison C, Testa NG and Dexter TM . (1986). Lancet, 1, 294–295.
Chen J, Shinkai Y, Young F and Alt FW . (1994). Curr. Opin. Immunol., 6, 313–319.
Conneally E, Cashman J, Petzer A and Eaves C . (1997). Proc. Natl. Acad. Sci. USA, 94, 9836–9841.
Coulombel L, Eaves AC and Eaves CJ . (1983a). Blood, 62, 291–297.
Coulombel L, Kalousek DK, Eaves CJ, Gupta CM and Eaves AC . (1983b). N. Engl. J. Med., 308, 1493–1498.
Croisille L, Auffray I, Katz A, Izac B, Vainchenker W and Coulombel L . (1994). Blood, 84, 4116–4124.
Cumano A, Dorshkind K, Gillis S and Paige CJ . (1990). Eur. J. Immunol., 20, 2183–2189.
Danet GH, Lee HW, Luongo JL, Simon MC and Bonnet DA . (2001). Exp. Hematol., 29, 1465–1473.
Denning-Kendall P, Singha S, Bradley B and Hows J . (2003). Stem Cells, 21, 694–701.
Dexter TM, Allen TD and Lajtha LG . (1977). J. Cell. Physiol., 91, 335–344.
Eaves AC, Barnett MJ, Ponchio L, Cashman JD, Petzer AL and Eaves CJ . (1998). Stem Cells, 16 (Suppl 1), 77–83 (discussion 89).
Eaves CJ and Eaves AC . (1978). Blood, 52, 1196–1210.
Fluckiger AC, Sanz E, Garcia-Lloret M, Su T, Hao QL, Kato R, Quan S, de la Hera A, Crooks GM, Witte ON and Rawlings DJ . (1998). Blood, 92, 4509–4520.
Galy A, Travis M, Cen D and Chen B . (1995). Immunity, 3, 459–473.
Gammaitoni L, Bruno S, Sanavio F, Gunetti M, Kollet O, Cavalloni G, Falda M, Fagioli F, Lapidot T, Aglietta M and Piacibello W . (2003). Exp. Hematol., 31, 261–270.
Glimm H, Eisterer W, Lee K, Cashman J, Holyoake TL, Nicolini F, Shultz LD, von Kalle C and Eaves CJ . (2001). J. Clin. Invest., 107, 199–206.
Godin I, Dieterlen-Lievre F and Cumano A . (1995). Proc. Natl. Acad. Sci. USA, 92, 773–777.
Hao QL, Smogorzewska EM, Barsky LW and Crooks GM . (1998). Blood, 91, 4145–4151.
Hao QL, Thiemann FT, Petersen D, Smogorzewska EM and Crooks GM . (1996). Blood, 88, 3306–3313.
Hao QL, Zhu J, Price MA, Payne KJ, Barsky LW and Crooks GM . (2001). Blood, 97, 3683–3690.
Hiramatsu H, Nishikomori R, Heike T, Ito M, Kobayashi K, Katamura K and Nakahata T . (2003). Blood, 102, 873–880.
Hodgson GS and Bradley TR . (1979). Nature, 281, 381–382.
Hogan CJ, Shpall EJ and Keller G . (2002). Proc. Natl. Acad. Sci. USA, 99, 413–418.
Hogan CJ, Shpall EJ, McNulty O, McNiece I, Dick JE, Shultz LD and Keller G . (1997). Blood, 90, 85–96.
Holyoake TL, Nicolini FE and Eaves CJ . (1999). Exp. Hematol., 27, 1418–1427.
Horn PA, Thomasson BM, Wood BL, Andrews RG, Morris JC and Kiem HP . (2003). Blood, 102, 4329–4335.
Iscove NN, Senn JS, Till JE and McCulloch EA . (1971). Blood, 37, 1–5.
Ishihara S, Tachibana N, Okayama A, Murai K, Tsuda K and Mueller N . (1992). Jpn. J. Cancer Res., 83, 320–323.
Issaad C, Croisille L, Katz A, Vainchenker W and Coulombel L . (1993). Blood, 81, 2916–2924.
Ivanova NB, Dimos JT, Schaniel C, Hackney JA, Moore KA and Lemischka IR . (2002). Science, 298, 601–604.
Jaleco AC, Neves H, Hooijberg E, Gameiro P, Clode N, Haury M, Henrique D and Parreira L . (2001). J. Exp. Med., 194, 991–1002.
Jetmore A, Plett PA, Tong X, Wolber FM, Breese R, Abonour R, Orschell-Traycoff CM and Srour EF . (2002). Blood, 99, 1585–1593.
Jones RJ, Wagner JE, Celano P, Zicha MS and Sharkis SJ . (1990). Nature, 347, 188–189.
Jordan CT and Lemischka IR . (1990). Genes Dev., 4, 220–232.
Jordan CT, McKearn JP and Lemischka IR . (1990). Cell, 61, 953–963.
Kamel-Reid S and Dick JE . (1988). Science, 242, 1706–1709.
Kawamoto H, Ohmura K and Katsura Y . (1997). Int. Immunol., 9, 1011–1019.
Keller G and Snodgrass R . (1990). J. Exp. Med., 171, 1407–1418.
Kerre TC, De Smet G, De Smedt M, Zippelius A, Pittet MJ, Langerak AW, De Bosscher J, Offner F, Vandekerckhove B and Plum J . (2002). Blood, 99, 1620–1626.
Kincade PW, Payne KJ, Tudor KS, Yamashita Y, Medina KL, Rossi MI and Kouro T . (2000). Curr. Top. Microbiol. Immunol., 251, 67–72.
Kollet O, Peled A, Byk T, Ben-Hur H, Greiner D, Shultz L and Lapidot T . (2000). Blood, 95, 3102–3105.
Kondo M, Weissman IL and Akashi K . (1997). Cell, 91, 661–672.
Kondo T, Setoguchi T and Taga T . (2004). Proc. Natl. Acad. Sci. USA, 101, 781–786.
Krosl J, Austin P, Beslu N, Kroon E, Humphries RK and Sauvageau G . (2003). Nat. Med., 9, 1428–1432.
Lacaud G, Carlsson L and Keller G . (1998). Immunity, 9, 827–838.
Lansdorp PM, Dragowska W and Mayani H . (1993). J. Exp. Med., 178, 787–791.
Lapidot T, Pflumio F, Doedens M, Murdoch B, Williams DE and Dick JE . (1992). Science, 255, 1137–1141.
Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, Minden M, Paterson B, Caligiuri MA and Dick JE . (1994). Nature, 367, 645–648.
Larochelle A, Vormoor J, Hanenberg H, Wang JC, Bhatia M, Lapidot T, Moritz T, Murdoch B, Xiao XL, Kato I, Williams DA and Dick JE . (1996). Nat. Med., 2, 1329–1337.
Leary AG and Ogawa M . (1987). Blood, 69, 953–956.
LeBien TW . (1989). Immunol. Today, 10, 296–298.
Lemieux ME and Eaves CJ . (1996). Blood, 88, 1639–1648.
Lemieux ME, Rebel VI, Lansdorp PM and Eaves CJ . (1995). Blood, 86, 1339–1347.
Lewis ID, McDiarmid LA, Samels LM, To LB and Hughes TP . (1998). Blood, 91, 630–640.
Lin Q, Dong C and Cooper MD . (1998). J. Exp. Med., 187, 79–87.
Lord BI and Woolford LB . (1993). Stem Cells, 11, 212–217.
Magli MC, Iscove NN and Odartchenko N . (1982). Nature, 295, 527–529.
Matsuzaki Y, Kinjo K, Mulligan RC and Okano H . (2004). Immunity, 20, 87–93.
Mayani H, Dragowska W and Lansdorp PM . (1993). J. Cell. Physiol., 157, 579–586.
McCulloch EA and Till JE . (1964). Radiat. Res., 22, 383–397.
McNiece I and Briddell R . (2001). Exp. Hematol., 29, 3–11.
Metcalf D and MacDonald HR . (1975). J. Cell. Physiol., 85, 643–654.
Miller CL and Eaves CJ . (1997). Proc. Natl. Acad. Sci. USA, 94, 13648–13653.
Morris JC, Conerly M, Thomasson B, Storek J, Riddell SR and Kiem HP . (2004). Blood, 103, 492–499.
Murdoch B, Chadwick K, Martin M, Shojaei F, Shah KV, Gallacher L, Moon RT and Bhatia M . (2003). Proc. Natl. Acad. Sci. USA, 100, 3422–3427.
Nakamura K, Kouro T, Kincade PW, Malykhin A, Maeda K and Coggeshall KM . (2004). J. Exp. Med., 199, 243–254.
Nakorn NT, Traver D, Weissman IL and Akashi K . (2002). J. Clin. Invest., 109, 1579–1585.
Norol F, Drouet M, Pflumio F, Leonardi M, Mourcin F, Debili N, Job A, Vainchenker W, Kuentz M and Herodin F . (2002). Br. J. Haematol., 117, 924–934.
Ohishi K, Varnum-Finney B and Bernstein ID . (2002). J. Clin. Invest., 110, 1165–1174.
Osawa M, Hanada K, Hamada H and Nakauchi H . (1996). Science, 273, 242–245.
Paige CJ, Gisler RH, McKearn JP and Iscove NN . (1984). Eur. J. Immunol., 14, 979–987.
Pardal R, Clarke MF and Morrison SJ . (2003). Nat. Rev. Cancer, 3, 895–902.
Pettengell R, Luft T, Henschler R, Hows JM, Dexter TM, Ryder D and Testa NG . (1994). Blood, 84, 3653–3659.
Petzer AL, Zandstra PW, Piret JM and Eaves CJ . (1996). J. Exp. Med., 183, 2551–2558.
Pflumio F, Izac B, Katz A, Shultz LD, Vainchenker W and Coulombel L . (1996). Blood, 88, 3731–3740.
Phillips RA . (1980). J. Supramol. Struct., 14, 77–83.
Ploemacher RE, van der Sluijs JP, van Beurden CA, Baert MR and Chan PL . (1991). Blood, 78, 2527–2533.
Ploemacher RE, van der Sluijs JP, Voerman JS and Brons NH . (1989). Blood, 74, 2755–2763.
Plum J, De Smedt M, Defresne MP, Leclercq G and Vandekerckhove B . (1994). Blood, 84, 1587–1593.
Plum J, De Smedt M, Verhasselt B, Kerre T, Vanhecke D, Vandekerckhove B and Leclercq G . (2000). Ann. NY Acad. Sci., 917, 724–731.
Punzel M, Moore KA, Lemischka IR and Verfaillie CM . (1999a). Leukemia, 13, 92–97.
Punzel M, Wissink SD, Miller JS, Moore KA, Lemischka IR and Verfaillie CM . (1999b). Blood, 93, 3750–3756.
Radtke F, Wilson A, Mancini SJ and MacDonald HR . (2004). Nat. Immunol., 5, 247–253.
Rawlings DJ, Quan S, Hao QL, Thiemann FT, Smogorzewska M, Witte ON and Crooks GM . (1997). Exp. Hematol., 25, 66–72.
Reya T, Duncan AW, Ailles L, Domen J, Scherer DC, Willert K, Hintz L, Nusse R and Weissman IL . (2003). Nature, 423, 409–414.
Reynaud D, Lefort N, Manie E, Coulombel L and Levy Y . (2003). Blood, 101, 4313–4321.
Robin C, Bennaceur-Griscelli A, Louache F, Vainchenker W and Coulombel L . (1999a). Br. J. Haematol., 104, 809–819.
Robin C, Pflumio F, Vainchenker W and Coulombel L . (1999b). J. Exp. Med., 189, 1601–1610.
Rollini P, Kaiser S, Faes-van’t Hull E, Kapp U and Leyvraz S . (2004). Blood, 103, 1166–1170.
Rosenzweig M, Marks DF, Zhu H, Hempel D, Mansfield KG, Sehgal PK, Kalams S, Scadden DT and Johnson RP . (1996). Blood, 87, 4040–4048.
Sato N, Meijer L, Skaltsounis L, Greengard P and Brivanlou AH . (2004). Nat. Med., 10, 55–63.
Sato T, Laver JH and Ogawa M . (1999). Blood, 94, 2548–2554.
Sauvageau G, Thorsteinsdottir U, Eaves CJ, Lawrence HJ, Largman C, Lansdorp PM and Humphries RK . (1995). Genes Dev., 9, 1753–1765.
Schmitt TM, de Pooter RF, Gronski MA, Cho SK, Ohashi PS and Zuniga-Pflucker JC . (2004). Nat. Immunol., 5, 410–417.
Schmitt TM and Zuniga-Pflucker JC . (2002). Immunity, 17, 749–756.
Shultz LD, Banuelos SJ, Leif J, Appel MC, Cunningham M, Ballen K, Burzenski L and Greiner DL . (2003). Exp. Hematol., 31, 551–558.
Spanopoulou E . (1996). Int. Rev. Immunol., 13, 257–288.
Srour EF, Jetmore A, Wolber FM, Plett PA, Abonour R, Yoder MC and Orschell-Traycoff CM . (2001). Leukemia, 15, 1681–1684.
Srour EF, Zanjani ED, Brandt JE, Leemhuis T, Briddell RA, Heerema NA and Hoffman R . (1992). Blood, 79, 1404–1412.
Sutherland HJ, Blair A and Zapf RW . (1996). Blood, 87, 4754–4761.
Sutherland HJ, Eaves CJ, Eaves AC, Dragowska W and Lansdorp PM . (1989). Blood, 74, 1563–1570.
Sutherland HJ, Eaves CJ, Lansdorp PM, Thacker JD and Hogge DE . (1991). Blood, 78, 666–672.
Sutherland HJ, Lansdorp PM, Henkelman DH, Eaves AC and Eaves CJ . (1990). Proc. Natl. Acad. Sci. USA, 87, 3584–3588.
Szilvassy SJ and Cory S . (1993). Blood, 81, 2310–2320.
Szilvassy SJ, Humphries RK, Lansdorp PM, Eaves AC and Eaves CJ . (1990). Proc. Natl. Acad. Sci. USA, 87, 8736–8740.
Szilvassy SJ, Ragland PL, Miller CL and Eaves CJ . (2003). Exp. Hematol., 31, 331–338.
Takano H, Ema H, Sudo K and Nakauchi H . (2004). J. Exp. Med., 199, 295–302.
Terstappen LW, Huang S, Safford M, Lansdorp PM and Loken MR . (1991). Blood, 77, 1218–1227.
Thiemann FT, Moore KA, Smogorzewska EM, Lemischka IR and Crooks GM . (1998). Exp. Hematol., 26, 612–619.
Traggiai E, Chicha L, Mazzucchelli L, Bronz L, Piffaretti JC, Lanzavecchia A and Manz MG . (2004). Science, 304, 104–107.
Trevisan M and Iscove NN . (1995). J. Exp. Med., 181, 93–103.
Uchida N, Dykstra B, Lyons KJ, Leung FY and Eaves CJ . (2003). Exp. Hematol., 31, 1338–1347.
Vormoor J, Lapidot T, Pflumio F, Risdon G, Patterson B, Broxmeyer HE and Dick JE . (1994). Blood, 83, 2489–2497.
Wagers AJ, Sherwood RI, Christensen JL and Weissman IL . (2002). Science, 297, 2256–2259.
Wang JC, Doedens M and Dick JE . (1997). Blood, 89, 3919–3924.
Wang JC, Lapidot T, Cashman JD, Doedens M, Addy L, Sutherland DR, Nayar R, Laraya P, Minden M, Keating A, Eaves AC, Eaves CJ and Dick JE . (1998). Blood, 91, 2406–2414.
Weekx SF, Snoeck HW, Offner F, De Smedt M, Van Bockstaele DR, Nijs G, Lenjou M, Moulijn A, Rodrigus I, Berneman ZN and Plum J . (2000). Blood, 95, 2806–2812.
Whitlock CA, Robertson D and Witte ON . (1984). J. Immunol. Methods, 67, 353–369.
Wu AM, Till JE, Siminovitch L and McCulloch EA . (1967). J. Cell. Physiol., 69, 177–184.
Yokota T, Kouro T, Hirose J, Igarashi H, Garrett KP, Gregory SC, Sakaguchi N, Owen JJ and Kincade PW . (2003). Immunity, 19, 365–375.
Zandstra PW, Conneally E, Petzer AL, Piret JM and Eaves CJ . (1997). Proc. Natl. Acad. Sci. USA, 94, 4698–4703.
Zanjani ED, Flake AW, Rice H, Hedrick M and Tavassoli M . (1994). J. Clin. Invest., 93, 1051–1055.
Zhang J, Niu C, Ye L, Huang H, He X, Tong WG, Ross J, Haug J, Johnson T, Feng JQ, Harris S, Wiedemann LM, Mishina Y and Li L . (2003). Nature, 425, 836–841.
Acknowledgements
I thank INSERM and Association pour la recherche contre le cancer (ARC) for their continuous support. I also thank all my collaborators and students who enriched my thoughts on stem cells through their discussions during all these years, and to the invaluable initial teaching of Connie Eaves who started it all for me.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Coulombel, L. Identification of hematopoietic stem/progenitor cells: strength and drawbacks of functional assays. Oncogene 23, 7210–7222 (2004). https://doi.org/10.1038/sj.onc.1207941
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.onc.1207941
