
Abstract
Tumor necrosis factor (TNF) receptor-associated factors (TRAFs) are cytoplasmic adapter proteins that link a wide variety of cell surface receptors to the apoptotic signaling cascade. The purpose of this study was to delineate the signaling pathways and TRAF1 promoter elements responsible for phorbol ester-mediated TRAF1 induction in human colon cancers. Here, we found that the PKC activators, phorbol 12-myristate 13-acetate (PMA) and bryostatin I, induced TRAF1 mRNA expression; pretreatment with actinomycin D blocked PMA-mediated TRAF1 expression suggesting induction at the transcriptional level. In contrast, expression of other TRAFs (TRAF2, 3 and 4) was minimally altered by PMA. Various PKC isoform-selective inhibitors blocked PMA-mediated TRAF1 mRNA and promoter stimulation; rottlerin, a selective PKCδ inhibitor, had no effect suggesting that Ca2+-dependent PKC isoforms (e.g., PKCα and βI) play a role in TRAF1 regulation. In addition, the MEK/ERK inhibitors, PD98059 and UO126, suppressed PMA-stimulated TRAF1 promoter activity indicating a role for ERK in TRAF1 induction. Moreover, cotransfection of a dominant-negative Raf-1 (Raf-C4) significantly reduced PMA-stimulated TRAF1 promoter activity whereas transfection of dominant-negative Ras or treatment with Ras inhibitors had minimal to no effect on TRAF1 induction suggesting dependence on Raf, but not Ras, activation. Finally, site-specific mutagenesis of functional NF-κB sites (particularly the most proximal site) in the TRAF1 promoter significantly decreased PMA-mediated promoter activity. In conclusion, our results demonstrate selective induction of TRAF1 in human colon cancer cells through a Ca2+-dependent PKC/Raf-1/ERK/NF-κB-dependent pathway.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
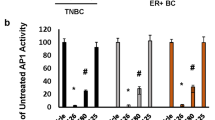
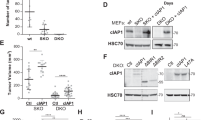
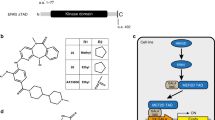
References
Adjei AA . (2001). J. Natl. Cancer Inst., 93, 1062–1074.
Arch RH, Gedrich RW and Thompson CB . (1998). Genes Dev., 12, 2821–2830.
Betzel C, Rachev R, Dolashka P and Genov N . (1993). Biochim. Biophys. Acta, 1161, 47–51.
Billman GE . (1993). Am. J. Physiol., 265, H1529–35.
Bradford MM . (1976). Anal. Biochem., 72, 248–254.
Bradley JR and Pober JS . (2001). Oncogene, 20, 6482–6491.
Brasier AR, Lu M, Hai T, Lu Y and Boldogh I . (2001). J. Biol. Chem., 276, 32080–32093.
Cao Z, Xiong J, Takeuchi M, Kurama T and Goeddel DV . (1996). Nature, 383, 443–446.
Carpentier I and Beyaert R . (1999). FEBS Lett., 460, 246–250.
Chen F, Castranova V, Shi X and Demers LM . (1999). Clin. Chem., 45, 7–17.
Chiloeches A, Paterson HF, Marais R, Clerk A, Marshall CJ and Sugden PH . (1999). J. Biol. Chem., 274, 19762–19770.
Deng J, Miller SA, Wang HY, Xia W, Wen Y, Zhou BP, Li Y, Lin SY and Hung MC . (2002). Cancer Cell, 2, 323–334.
Deng L, Wang C, Spencer E, Yang L, Braun A, You J, Slaughter C, Pickart C and Chen ZJ . (2000). Cell, 103, 351–361.
Dhawan P and Richmond A . (2002). J. Biol. Chem., 277, 7920–7928.
Dhillon AS, Meikle S, Peyssonnaux C, Grindlay J, Kaiser C, Steen H, Shaw PE, Mischak H, Eychene A and Kolch W . (2003). Mol. Cell Biol., 23, 1983–1993.
Dunn IF, Geha RS and Tsitsikov EN . (1999). Mol. Immunol., 36, 611–617.
Evers BM, Zhou Z, Celano P and Li J . (1995). J. Clin. Invest., 95, 2822–2830.
Fang X, Yu S, Tanyi JL, Lu Y, Woodgett JR and Mills GB . (2002). Mol. Cell. Biol., 22, 2099–2110.
Finder JD, Litz JL, Blaskovich MA, McGuire TF, Qian Y, Hamilton AD, Davies P and Sebti SM . (1997). J. Biol. Chem., 272, 13484–13488.
Gekeler V, Boer R, Uberall F, Ise W, Schubert C, Utz I, Hofmann J, Sanders KH, Schachtele C, Klemm K and Grunicke H . (1996). Br. J. Cancer, 74, 897–905.
Gutierrez L, Magee AI, Marshall CJ and Hancock JF . (1989). EMBO J., 8, 1093–1098.
Han SS, Keum YS, Seo HJ and Surh YJ . (2002). J. Biochem. Mol. Biol., 35, 337–342.
Hennings H, Blumberg PM, Pettit GR, Herald CL, Shores R and Yuspa SH . (1987). Carcinogenesis, 8, 1343–1346.
Hernandez A, Wang QD, Schwartz SA and Evers BM . (2001). J. Gastrointest. Surg., 5, 56–65.
Hipp MS, Urbich C, Mayer P, Wischhusen J, Weller M, Kracht M and Spyridopoulos I . (2002). Eur. J. Immunol., 32, 2208–2217.
Inoue J, Ishida T, Tsukamoto N, Kobayashi N, Naito A, Azuma S and Yamamoto T . (2000). Exp. Cell Res., 254, 14–24.
Irmler M, Steiner V, Ruegg C, Wajant H and Tschopp J . (2000). FEBS Lett., 468, 129–133.
Ishida TK, Tojo T, Aoki T, Kobayashi N, Ohishi T, Watanabe T, Yamamoto T and Inoue J . (1996). Proc. Natl. Acad. Sci. USA, 93, 9437–9442.
Jaken S . (1996). Curr. Opin. Cell Biol., 8, 168–173.
James G, Goldstein JL and Brown MS . (1996). Proc. Natl. Acad. Sci. USA, 93, 4454–4458.
James GL, Goldstein JL and Brown MS . (1995). J. Biol. Chem., 270, 6221–6226.
Jiang B, Brecher P and Cohen RA . (2001). Arterioscler. Thromb. Vasc. Biol., 21, 1915–1920.
Jiang B, Xu S, Brecher P and Cohen RA . (2002). Arterioscler. Thromb. Vasc. Biol., 22, 1811–1816.
Jiang S, Chow SC, Nicotera P and Orrenius S . (1994). Exp. Cell Res., 212, 84–92.
Karin M and Ben-Neriah Y . (2000). Annu. Rev. Immunol., 18, 621–663.
Karin M and Lin A . (2002). Nat. Immunol., 3, 221–227.
Kolch W, Heidecker G, Kochs G, Hummel R, Vahidi H, Mischak H, Finkenzeller G, Marme D and Rapp UR . (1993). Nature, 364, 249–252.
Ku WC, Cheng AJ and Wang TC . (1997). Biochem. Biophys. Res. Commun., 241, 730–736.
Kurland JF, Voehringer DW and Meyn RE . (2003). J. Biol. Chem., 278, 32465–32470.
Lee HC . (1993). J. Biol. Chem., 268, 293–299.
Lee HW, Ahn DH, Crawley SC, Li JD, Gum Jr JR, Basbaum CB, Fan NQ, Szymkowski DE, Han SY, Lee BH, Sleisenger MH and Kim YS . (2002). J. Biol. Chem., 277, 32624–32631.
Lee SY, Park CG and Choi Y . (1996). J. Exp. Med., 183, 669–674.
Lerner EC, Qian Y, Hamilton AD and Sebti SM . (1995). J. Biol. Chem., 270, 26770–26773.
Lerner EC, Zhang TT, Knowles DB, Qian Y, Hamilton AD and Sebti SM . (1997). Oncogene, 15, 1283–1288.
Mercurio F and Manning AM . (1999). Curr. Opin. Cell Biol., 11, 226–232.
Moores SL, Schaber MD, Mosser SD, Rands E, O'Hara MB, Garsky VM, Marshall MS, Pompliano DL and Gibbs JB . (1991). J. Biol. Chem., 266, 14603–14610.
Mosialos G, Birkenbach M, Yalamanchili R, VanArsdale T, Ware C and Kieff E . (1995). Cell, 80, 389–399.
Mulvaney JM and Roberson MS . (2000). J. Biol. Chem., 275, 14182–14189.
Nakano H, Shindo M, Sakon S, Nishinaka S, Mihara M, Yagita H and Okumura K . (1998). Proc. Natl. Acad. Sci. USA, 95, 3537–3542.
Ni CZ, Welsh K, Leo E, Chiou CK, Wu H, Reed JC and Ely KR . (2000). Proc. Natl. Acad. Sci. USA, 97, 10395–10399.
Nishizuka Y . (1995). FASEB J., 9, 484–496.
Okumura K, Shirasawa S, Nishioka M and Sasazuki T . (1999). Cancer Res., 59, 2445–2450.
Pahl HL, Krauss B, Schulze-Osthoff K, Decker T, Traenckner EB, Vogt M, Myers C, Parks T, Warring P, Muhlbacher A, Czernilofsky AP and Baeuerle PA . (1996). J. Exp. Med., 183, 1829–1840.
Parekh DB, Ziegler W and Parker PJ . (2000). EMBO J., 19, 496–503.
Park YC, Burkitt V, Villa AR, Tong L and Wu H . (1999). Nature, 398, 533–538.
Pearson G, English JM, White MA and Cobb MH . (2001). J. Biol. Chem., 276, 7927–7931.
Regnier CH, Tomasetto C, Moog-Lutz C, Chenard MP, Wendling C, Basset P and Rio MC . (1995). J. Biol. Chem., 270, 25715–25721.
Rothe M, Wong SC, Henzel WJ and Goeddel DV . (1994). Cell, 78, 681–692.
Schaber MD, O'Hara MB, Garsky VM, Mosser SC, Bergstrom JD, Moores SL, Marshall MS, Friedman PA, Dixon RA and Gibbs JB . (1990). J. Biol. Chem., 265, 14701–14704.
Schwenzer R, Siemienski K, Liptay S, Schubert G, Peters N, Scheurich P, Schmid RM and Wajant H . (1999). J. Biol. Chem., 274, 19368–19374.
Seternes OM, Johansen B and Moens U . (1999). Mol. Endocrinol., 13, 1071–1083.
Song HY and Donner DB . (1995). Biochem. J., 309 (Part 3), 825–829.
Takaesu G, Kishida S, Hiyama A, Yamaguchi K, Shibuya H, Irie K, Ninomiya-Tsuji J and Matsumoto K . (2000). Mol. Cell, 5, 649–658.
Taylor MT, Lawson KR, Ignatenko NA, Marek SE, Stringer DE, Skovan BA and Gerner EW . (2000). Cancer Res., 60, 6607–6610.
Tsao DH, McDonagh T, Telliez JB, Hsu S, Malakian K, Xu GY and Lin LL . (2000). Mol. Cell, 5, 1051–1057.
Ueda Y, Hirai S, Osada S, Suzuki A, Mizuno K and Ohno S . (1996). J. Biol. Chem., 271, 23512–23519.
Verin AD, Liu F, Bogatcheva N, Borbiev T, Hershenson MB, Wang P and Garcia JG . (2000). Am. J. Physiol. Lung Cell Mol. Physiol., 279, L360–70.
Vuong H, Patterson T, Shapiro P, Kalvakolanu DV, Wu R, Ma WY, Dong Z, Kleeberger SR and Reddy SP . (2000). J. Biol. Chem., 275, 32250–32259.
Wajant H, Haas E, Schwenzer R, Muhlenbeck F, Kreuz S, Schubert G, Grell M, Smith C and Scheurich P . (2000). J. Biol. Chem., 275, 24357–24366.
Wajant H, Henkler F and Scheurich P . (2001). Cell Signal, 13, 389–400.
Wang D, Yu X and Brecher P . (1998b). J. Biol. Chem., 273, 33027–33034.
Wang CY, Mayo MW, Korneluk RG, Goeddel DV and Baldwin Jr AS . (1998a). Science, 281, 1680–1683.
Wang Q, Wang X, Hernandez A, Hellmich MR, Gatalica Z and Evers BM . (2002). J. Biol. Chem., 277, 36602–36610.
Wang Q, Wang X, Hernandez A, Kim S and Evers BM . (2001). Gastroenterology, 120, 1381–1392.
Wu MH and Yung BY . (1994). Eur. J. Pharmacol., 270, 203–212.
Zapata JM, Krajewska M, Krajewski S, Kitada S, Welsh K, Monks A, McCloskey N, Gordon J, Kipps TJ, Gascoyne RD, Shabaik A and Reed JC . (2000). J. Immunol., 165, 5084–5096.
Zhang FL and Casey PJ . (1996). Annu. Rev. Biochem., 65, 241–269.
Zhao D, Keates AC, Kuhnt-Moore S, Moyer MP, Kelly CP and Pothoulakis C . (2001). J. Biol. Chem., 276, 44464–44471.
Zou Y, Komuro I, Yamazaki T, Aikawa R, Kudoh S, Shiojima I, Hiroi Y, Mizuno T and Yazaki Y . (1996). J. Biol. Chem., 271, 33592–33597.
Acknowledgements
We thank Eileen Figueroa, Mary Lou Mraz and Karen Martin for manuscript preparation. We also thank Drs Mark Hellmich and Kathleen O'Connor and members of the Evers laboratory for helpful comments and discussion. This work was supported by Grants RO1 DK48498 and PO1 DK35408 from the National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wang, X., Wang, Q., Hu, W. et al. Regulation of phorbol ester-mediated TRAF1 induction in human colon cancer cells through a PKC/RAF/ERK/NF-κB-dependent pathway. Oncogene 23, 1885–1895 (2004). https://doi.org/10.1038/sj.onc.1207312
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.onc.1207312
Keywords
This article is cited by
-
miR-373 promotes invasion and metastasis of colorectal cancer cells via activating ERK/MAPK pathway
Scientific Reports (2024)
-
The renin-angiotensin-aldosterone system (RAAS) signaling pathways and cancer: foes versus allies
Cancer Cell International (2023)
-
Immunohistochemical investigation of prognostic biomarkers in resected colorectal liver metastases: a systematic review and meta-analysis
Cancer Cell International (2018)
-
Altered expression of Tumor Necrosis Factor Alpha -Induced Protein 3 correlates with disease severity in Ulcerative Colitis
Scientific Reports (2017)
-
The DEAD box protein p68: a crucial regulator of AKT/FOXO3a signaling axis in oncogenesis
Oncogene (2015)
