
Abstract
Central nervous system nutrient sensing and afferent endocrine signaling have been established as parallel systems communicating metabolic status and energy availability in vertebrates. The only afferent endocrine signal known to require modification with a fatty acid side chain is the orexigenic hormone ghrelin. We find that the ghrelin O-acyl transferase (GOAT), which is essential for ghrelin acylation, is regulated by nutrient availability, depends on specific dietary lipids as acylation substrates and links ingested lipids to energy expenditure and body fat mass. These data implicate the ghrelin-GOAT system as a signaling pathway that alerts the central nervous system to the presence of dietary calories, rather than to their absence as is commonly accepted.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others
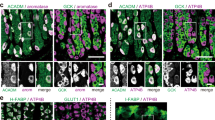
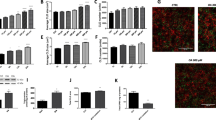

Change history
04 September 2009
In the version of this article initially published, the fourth condition from the top in the key to the bar graphs in Figure 2c was mislabeled as ‘mC8’. The correct label is ‘hC8’. The error has been corrected in the HTML and PDF versions of the article.
References
Obici, S. et al. J. Clin. Invest. 108, 1079–1085 (2001).
Schwartz, M.W., Woods, S.C., Porte, D. Jr., Seeley, R.J. & Baskin, D.G. Nature 404, 661–671 (2000).
Cao, H. et al. Cell 134, 933–944 (2008).
Gillum, M.P. et al. Cell 135, 813–824 (2008).
Kojima, M. et al. Nature 402, 656–660 (1999).
López, M. et al. Cell Metab. 7, 389–399 (2008).
Tschöp, M., Smiley, D.L. & Heiman, M.L. Nature 407, 908–913 (2000).
Kamegai, J. et al. Diabetes 50, 2438–2443 (2001).
Tschöp, M. et al. J. Endocrinol. Invest. 24, RC19–RC21 (2001).
Gutierrez, J.A. et al. Proc. Natl. Acad. Sci. USA 105, 6320–6325 (2008).
Yang, J., Brown, M.S., Liang, G., Grishin, N.V. & Goldstein, J.L. Cell 132, 387–396 (2008).
Nishi, Y. et al. Endocrinology 146, 2255–2264 (2005).
Drazen, D.L., Vahl, T.P., D'Alessio, D.A., Seeley, R.J. & Woods, S.C. Endocrinology 147, 23–30 (2006).
Liu, J. et al. J. Clin. Endocrinol. Metab. 93, 1980–1987 (2008).
Dole, V.P. & Rizack, M.A. J. Lipid Res. 2, 90–91 (1961).
Schwartz, G.J. et al. Cell Metab. 8, 281–288 (2008).
Zurier, R.B., Campbell, R.G., Hashim, S.A. & Van Itallie, T.B. N. Engl. J. Med. 274, 490–493 (1966).
Prior, I.A., Davidson, F., Salmond, C.E. & Czochanska, Z. Am. J. Clin. Nutr. 34, 1552–1561 (1981).
Pfluger, P.T. et al. Am. J. Physiol. Gastrointest. Liver Physiol. 294, G610–G618 (2008).
Gauna, C. et al. Am. J. Physiol. Endocrinol. Metab. 293, E697–E704 (2007).
Acknowledgements
We would like to thank J.E. Baker, B. Berger and J. Holland for their outstanding technical support. This manuscript was supported by the Leibniz Graduate College (H.K.) and by the US National Institutes of Health/National Institute of Diabetes and Digestive and Kidney Diseases Grant R01-DK069987 (to M.H.T.).
Author information
Authors and Affiliations
Contributions
H.K., P.T.P. and J.A.G. designed and performed most of the experiments and wrote the manuscript; P.J.S. generated mouse models; J.A.W. performed and interpreted mass spectrometry analyses; T.A.C. performed gene expression analyses; A.S., H.-G.J., R.J.J. and J.E.H. designed experiments, interpreted data and wrote the manuscript; and M.L.H. and M.H.T. conceptualized, analyzed and interpreted all studies and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
J.A.G., P.J.S., T.A.C., J.A.W., J.E.H. and M.L.H. are employees and stock holders of the Eli Lilly Company.
Supplementary information
Supplementary Text and Figures
Supplementary Methods and Supplementary Figs. 1–7 (PDF 424 kb)
Rights and permissions
About this article
Cite this article
Kirchner, H., Gutierrez, J., Solenberg, P. et al. GOAT links dietary lipids with the endocrine control of energy balance. Nat Med 15, 741–745 (2009). https://doi.org/10.1038/nm.1997
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nm.1997
This article is cited by
-
Methylation analysis by targeted bisulfite sequencing in large for gestational age (LGA) newborns: the LARGAN cohort
Clinical Epigenetics (2023)
-
Childhood sexual abuse is associated with higher total ghrelin serum levels in adulthood: results from a large, population-based study
Translational Psychiatry (2023)
-
Ghrelin hormone a new molecular modulator between obesity and glomerular damage
Molecular Biology Reports (2023)
-
The novel ghrelin receptor inverse agonist PF-5190457 administered with alcohol: preclinical safety experiments and a phase 1b human laboratory study
Molecular Psychiatry (2020)
-
Ghrelin and liver disease
Reviews in Endocrine and Metabolic Disorders (2020)
