
Abstract
Self-renewing cell populations such as hematopoietic stem cells and memory B and T lymphocytes might be regulated by shared signaling pathways1. The Wnt–β-catenin pathway is an evolutionarily conserved pathway that promotes hematopoietic stem cell self-renewal and multipotency by limiting stem cell proliferation and differentiation2,3, but its role in the generation and maintenance of memory T cells is unknown. We found that induction of Wnt–β-catenin signaling by inhibitors of glycogen sythase kinase-3β or the Wnt protein family member Wnt3a arrested CD8+ T cell development into effector cells. By blocking T cell differentiation, Wnt signaling promoted the generation of CD44lowCD62LhighSca-1highCD122highBcl-2high self-renewing multipotent CD8+ memory stem cells with proliferative and antitumor capacities exceeding those of central and effector memory T cell subsets. These findings reveal a key role for Wnt signaling in the maintenance of 'stemness' in mature memory CD8+ T cells and have major implications for the design of new vaccination strategies and adoptive immunotherapies.
Similar content being viewed by others

Main
T cell factor-1 (Tcf-1) and lymphoid enhancer-binding factor-1 (Lef-1) are downstream transcription factors of the Wnt–β-catenin signaling pathway. Tcf-1 and Lef-1 are required for normal thymic T cell development, but less is known about Wnt function in mature T cells2,4. Although experiments using a multimerized Tcf-Lef binding site reporter system have revealed that Wnt signaling is active in mature CD8+ T cells, the impact of this pathway on this cell population has yet to be fully elucidated5. At least three lines of evidence indicate that Wnt signaling might regulate the maturation of post-Thymic T lymphocytes. CD8+ T cells from Tcf7−/− mice, which are missing the gene that encodes Tcf-1, show a more differentiated phenotype (CD44high and CD62Llow) than wild-type (WT) T cells6. In addition, expression of Lef1 and Tcf7 decreases with progressive differentiation of CD8+ T cells from naive T cells (TN cells) to central memory T cells (TCM cells) and effector memory T cells (TEM cells) in humans7 and mice (Supplementary Fig. 1). Finally, high levels of Ctnnb1 (which encodes β-catenin), Lef1 and Tcf7 have been detected in T cells with increased potential to form memory cells in vivo8,9. Thus, triggering the activities of the Wnt signaling transcription factors Tcf-1 and Lef-1 might promote stem-like self-renewal capacity in mature T cells.
To test the impact of Wnt–β-catenin signaling on mature CD8+ T cells, we primed TN cells in the presence of the 4,6-disubstituted pyrrolopyrimidine TWS119, a potent inhibitor of the serine-threonine kinase glycogen synthase kinase-3β (Gsk-3β)10. Gsk-3β blockade mimics Wnt signaling by promoting the accumulation of β-catenin, the molecule that tethers the Tcf and Lef transcription factors to targeted DNA2. TWS119 triggered a rapid accumulation of β-catenin (mean 6.8 ± s.d. 1.7-fold increase by densitometry; P < 0.05) (Fig. 1a), augmented nuclear protein interaction with oligonucleotide containing the DNA sequences to which Tcf and Lef bind (Fig. 1b) and sharply upregulated the expression of Tcf7 (ref. 11), Lef1 (ref. 12) and other Wnt target genes including Jun13, Fzd7 (encoding Frizzled-7)14, Nlk (encoding Nemo-like kinase)15 (Fig. 1c). By contrast, T cell activation in the absence of the Gsk-3β inhibitor resulted in the downregulation of the Wnt–β-catenin signaling cascade (Fig. 1a–c). Thus, TWS119 activated the Wnt–β-catenin pathway in naive T cells and reversed the physiological downregulation of Tcf7 and Lef1 induced by T cell activation7.

Naive CD8+ T cells were primed in vitro with antibody to CD3 (2 μg ml−1) and antibody to CD28 (1 μg ml−1) with or without 7 μM TWS119. (a) Western blot analysis of β-catenin and glyceraldehyde-3-phosphate dehydrogenase (Gapdh) in CD8+ T cells treated with or without TWS119. (b) Electrophoretic mobility shift assay of nuclear extract from CD8+ T cells treated with or without TWS119 using P32-labeled oligonucleotide probes (oligo) designed from the Tcf and Lef binding region of the Tcf-1 target gene Fzd7. Unlabeled oligonucleotide probes were used as competitors. (c) Quantitative RT-PCR analysis of the expression of Tcf7, Lef1, Jun, Fzd7 and Nlk in CD8+ T cells treated with or without TWS119. Data are represented as means ± s.e.m. All data are representative of at least two independently performed experiments.
We sought to assess the effect of Wnt signaling on CD8+ T cell differentiation and proliferation. We stimulated carboxyfluorescein succinimidyl ester (CFSE)-labeled CD8+ T cells from the gp100-specific T cell receptor (TCR)-transgenic mouse, pmel-1 (ref. 16), with its cognate antigen in the presence of titrated doses of TWS119 and analyzed them for the expression of the differentiation markers CD44 and CD62 ligand (CD62L). CD44 expression is known to increase with T cell differentiation, whereas CD62L expression is progressively lost17. Treatment of TWS119 increased the frequency of T cells that retained CD62L expression in a dose-dependent manner, indicating that it inhibits CD8+ T cell differentiation (Fig. 2a). Notably, 46% of CD8+ T cells cultured in the presence of the highest concentration of Gsk-3β inhibitor failed to upregulate CD44, maintaining a naive CD44lowCD62Lhigh phenotype (Fig. 2a). Low doses of TWS119 (≤1 μM) preserved CD62L expression without affecting T cell proliferation, whereas higher TWS119 concentrations inhibited cell cycling in a dose-dependent manner (Fig. 2b). Arrested differentiation and proliferation of CD8+ T cells mediated by TWS119 were not secondary to its impact on dendritic cells (DCs) because we observed similar results when we stimulated purified CD8+ T cells in a DC-free system (Supplementary Fig. 2a,b). Similarly to TWS119, the structurally unrelated Gsk-3β inhibitor 6-bromo-substituted indirubin, BIO18,19, inhibited T cell differentiation (Supplementary Fig. 3a) and induced Tcf7 and Lef1 expression (Supplementary Fig. 3b). An analog with a greater Gsk-3β kinase inhibitory specificity, BIO-acetoxime19 retained the observed activity, whereas the N-methylated analog (methyl-BIO)19, which does not block the kinase activity of Gsk-3β, had no effect (Supplementary Fig. 3a,b). These results are in contrast with those obtained by other researchers using lithium chloride20, a less active and less specific Gsk-3β inhibitor19. Because Gsk-3β regulates several signaling pathways other than the Wnt pathway, we sought to more directly test whether the impact of the pharmacological blockade of Gsk-3β was dependent on mimicking the downstream signals of the Wnt–β-catenin pathway. We primed CD8+ T cells in the presence of Wnt3A, a Wnt protein that has been shown to promote hematopoietic stem cell self-renewal and pluripotency via β-catenin accumulation in the cell nucleus21. Similarly to TWS119, Wnt3A itself inhibited T cell differentiation and proliferation (Supplementary Fig. 4). Thus, T cell proliferation and differentiation could be restrained through the activation of the Wnt–β-catenin pathway by the naturally occurring ligand Wnt3A and by the pharamcologic inhibition of Gsk-3β downstream. Nevertheless, these data do not rule out the possibility that Gsk-3β inhibitors regulate T cell differentiation by affecting other pathways in addition to the Wnt pathway.

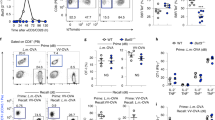
(a–c) Flow cytometry analysis. Numbers represent the percentage of cells in the dot-plot quadrants. (a) CFSE dilution assays (b) and cytokine and 51Cr release assays (c) of CFSE-labeled naive pmel-1 CD8+ T cells primed in vitro with CD8+ T cell depleted splenocytes pulsed with 1 μM hgp10025–33, in conjunction with 10 ng ml−1 IL-2 and titrated doses of TWS119, 4 d after T cell activation. Data in c are represented as means ± s.e.m. E:T, effector to target ratio. (d) Quantitative RT-PCR analysis of the expression of Eomes in CD8+ T cells after priming with CD3-specific and CD28-specific antibodies with or without 7 μM TWS119. Data are represented as means ± s.e.m. (e) Flow cytometry and enumeration of T cell subsets 6 d after vaccination with or without TWS119. WT mice received adoptive transfer of 1 × 106 naive pmel-1 Thy1.1+ CD8+ T cells in conjunction with recombinant fowlpox-based hgp100 vaccine. Mice received four daily doses of TWS119 (at 30 mg kg−1) from day 0 to day 3 or DMSO as control. All data are representative of at least two independently performed experiments.
We sought to evaluate whether the phenotypic arrest of differentiation of pmel-1 CD8+ T cells that was induced by TWS119 was associated with a block in the acquisition of effector functions. It has been previously shown that differentiating CD8+ T cells lose the capacity to produce interleukin-2 (IL-2) as they acquire the ability to kill target cells and release large amounts of interferon-γ (IFN-γ)17,22. We found that TWS119 induced a dose-dependent decrease in T cell–specific killing and IFN-γ release associated with a preservation of the ability to produce IL-2 (Fig. 2c). These functional data confirmed our phenotypic findings that TWS119 was a negative regulator of effector T cell (Teff cell) differentiation. We next assessed whether inhibition of Teff cell development is associated with the suppression of eomesodermin (encoded by Eomes), a master regulator of CD8+ T cell effector function23. We found that exposure of cells to TWS119 abrogated the induced expression of Eomes that occurred within 8 h after T cell priming, indicating that TWS119-mediated suppression of the effector program is an early event (Fig. 2d). Altogether, phenotypic, functional and molecular data indicated that induction of Wnt signaling during T cell priming results in inhibition of CD8+ T cell differentiation into Teff cells. Given our in vitro findings, we tested whether TWS119 can influence the qualities of adoptively transferred pmel-1 CD8+ T cells in response to fowlpox-based gp100 immunization and prevent the induction of highly differentiated, senescent T cells, a major pitfall of current T cell–based vaccines24. We found that there were no differences in frequency or numbers of pmel-1 T cells in spleens 6 d after vaccination, but development of Teff cells was inhibited (Fig. 2e). More notably, TWS119 effectively altered CD8+ T cell differentiation after immunization, as evidenced by increased numbers of CD44highCD62Lhigh TCM cells in responding CD8+ T cells after vaccination (Fig. 2e). However, we cannot ascertain whether the arrest of CD8+ T cell differentiation that we observed in vivo was a direct effect of TWS119 on this cell population or instead indirectly mediated through another cell type.
We observed a cell population that expressed low levels of CD44 and high levels of CD62L on the cell surface when we administered TWS119 in vitro and in vivo (Fig. 2a,e) but it was unclear whether these cells remained naive after antigen encounter or had entered into a primordial memory state that retained some phenotypic traits of TN cells. In a mouse model of graft-versus-host disease, other researchers have described a population of CD44lowCD62Lhigh memory CD8+ T cells that express high cell surface levels of stem cell antigen-1 (Sca-1), B-cell lymphoma protein-2 (Bcl-2) and common IL-2 and IL-15 receptor β chain (CD122)25. Because these cells showed robust self-renewal and the multipotent capacity to generate TCM, TEM and Teff cells, they were designated 'T memory stem cells' (TSCM cells)25. We sought to explore whether the CD44lowCD62Lhigh pmel-1 T cells that we generated in vitro with TWS119 treatment are TSCM cells. We found that the CD44lowCD62Lhigh T cells generated after priming with antigen in the presence of TWS119 had undergone up to four divisions, as indicated by CFSE dilution, and uniformly expressed high cell surface levels of the core phenotypic markers of TSCM cells, namely Sca-1, CD122 and Bcl-2 (Fig. 3a). By contrast, freshly isolated TN cells expressed these markers in small amounts (Fig. 3a). Similarly, Sca-1, CD122 and Bcl-2 were upregulated in CD44lowCD62Lhigh pmel-1 T cells generated in vitro in the presence of BIO-acetoxime (Supplementary Fig. 5) or Wnt3A (Supplementary Fig. 6) and in vivo after vaccination with gp100 and TWS119 administration (Supplementary Fig. 7). These activation and memory T cell markers were not exclusively expressed by TSCM cells but rather defined TSCM cells in the context of cells the were also CD44low and CD62Lhigh (data not shown). Antigen-experienced memory T cells can be distinguished from TN cells not only by phenotype but also by a number of functional properties, including rapid acquisition of effector functions upon antigen rechallenge26,27,28, pronounced cell cycling capacity29, robust homeostatic proliferation30 and independence from major histocompatibility complex (MHC) class I for persistence31 and antitumor activity32. We found that, unlike TN cells, TWS119-induced CD44lowCD62Lhigh T cells rapidly released cytokines (IFN-γ and IL-2) upon antigen encounter (Fig. 3b), had undergone more cell division after adoptive transfer into a lymphoreplete host (Fig. 3c) or into sublethally irradiated or genetically lymphodepleted Tcra−/− (T cell–deficient) or Rag1−/− (T and B cell–deficient) hosts (Fig. 3d), and persisted and mediated tumor destruction (P < 0.05) in B2m−/− mice (which are MHC class I deficient) (Fig. 3e,f). Altogether, these findings indicated that Wnt signaling induces the generation of a TSCM cell–like population that possesses the rapid recall ability, proliferative capacity and MHC class I independence that are characteristic of memory but not naive T cells.

CFSE-labeled, naive pmel-1 CD8+ T cells were primed in vitro with CD8+ T cell–depleted splenocytes pulsed with 1 μM hgp10025–33, in conjunction with 10 ng ml−1 IL-2 and 7 μM TWS119. (a) Flow cytometry analysis of TWS119-treated pmel-1 T cells 4 d after T cell activation and freshly isolated naive pmel-1 T cells. Numbers represent the mean fluorescence intensity in the histogram plots of Sca-1, CD122 and Bcl-2 and the percentage of cells in the dot-plot quadrants and the CFSE histogram. NA, not applicable. (b) Cytokine release assay of sorted CD44lowCD62Lhigh TWS119-treated pmel-1 T cells 5 d after antigenic stimulation and freshly isolated naive pmel-1 T cells. Data are represented as means ± s.e.m. np, nucleoprotein peptide. (c,d) CFSE dilution of sorted CD44lowCD62Lhigh TWS119-treated or freshly isolated naive pmel-1 T cells congenically marked with Thy 1.1+ (c) or Ly5.1+ (d) pmel-1 T cells 1 month after transfer into WT (c) or sublethally irradiated WT, Tcra−/−,or Rag1−/− mice (d). Data are shown on Thy 1.1+ (c) or Ly5.1+ (d) CD8+ lymphocytes. Numbers represent the percentage of cells in the dot-plot quadrants. (e) Flow cytometry analysis of sorted TWS119-treated and naive Ly5.1+ pmel-1 T cells 1 month after transfer into WT or B2m−/− mice. Numbers represent the percentage of cells in the dot-plot quadrants. (f) Tumor size after treatment of myeloablated WT or B2m−/− mice bearing B16 tumors established for 7 d. Mice received syngeneic age-matched, lineage-depleted bone marrow cells. On the next day, 1 × 106 CD44lowCD62Lhigh TWS119-treated or freshly isolated naive pmel-1 T cells were transferred in conjunction with intraperitoneal injection of IL-2. (g,h) Flow cytometry analysis of CFSE-labeled, sorted CD44lowCD62Lhigh TWS119-treated or freshly isolated naive pmel-1 T cells congenically marked with Ly5.1+ 1 month after transfer into sublethally irradiated WT mice. Data are represented as the percentage of CD44lowCD62Lhigh cells as a function of CFSE dilution of two independent experiments (g) or as fraction of cells with any given number of divisions (h). All data are representative of at least two independently performed experiments.
The concept of stemness encompasses the capability both to self-renew and to generate more differentiated, specialized cells. We sought to determine the stemness of TWS119-generated CD44lowCD62Lhigh T cells by evaluating the fate of these cells after adoptive transfer into sublethally irradiated mice. Four weeks later, we found that high percentages of TWS119-generated cells preserved their original CD44lowCD62Lhigh phenotype even after multiple cell divisions, whereas TN cells rapidly acquired CD44 with progressive CFSE dilution (P < 0.02; Fig. 3g,h). The differentiation of TWS119-induced CD44lowCD62Lhigh T cells was not arrested, however, because they acquired high CD44 expression, albeit at a slower pace than TN cells (Fig. 3g,h). Unlike TN cells, TWS119-derived CD44lowCD62LhighSca-1high T cells robustly proliferated (by 28 d after transfer into sublethally irradiated mice, 95% had undergone cell division) (Fig. 3h), enabling their reisolation and transfer to a secondary sublethally irradiated recipient (Supplementary Fig. 8). After 4 weeks, we found that the secondarily transferred CD44lowCD62LhighSca-1high T cells had again regenerated all T cell subsets. Notably, 43% of the cells retained the TSCM phenotype in spite of a second round of robust proliferation in vivo (Supplementary Fig. 8). These findings indicated that Wnt signaling promoted the generation of self-renewing, multipotent TSCM cells.
Having established that the CD44lowCD62Lhigh T cells generated in the presence of TWS119 are bona fide TSCM cells and not merely naive, we sought to compare them with the well-defined memory T cell subsets, TCM cells and TEM cells, in secondary challenge experiments. We adoptively transferred highly purified pmel-1 memory T cell subsets into sublethally irradiated mice in combination with recombinant vaccinia-based gp100 vaccine and IL-2 treatment. As previously reported by our group and others33,34, the TCM cells proliferated more than TEM cells (Fig. 4a). TSCM cells, however, robustly expanded by approximately 200-fold in the spleen alone (Fig. 4a). These levels of expansion were approximately ten times higher than those of TCM cells and approximately 30 times higher than those of TEM cells (Fig. 4a). Furthermore, TSCM cells showed an enhanced survival capacity, as revealed by the frequencies of pmel-1 T cells in the spleens of vaccinated mice 1 month after transfer (Fig. 4b). T cell proliferative and survival capacities have been correlated with tumor responses in mice and humans receiving adoptive T cell–based therapies35,36. To assess whether the enhanced replicative capability of TSCM cells would result in superior antitumor activity, we adoptively transferred limiting numbers of pmel-1 memory T cell subsets into sublethally irradiated hosts bearing B16 tumors, in combination with gp100 vaccine and IL-2. As we previously reported33, TCM cells conferred superior antitumor immunity compared with TEM cells (for tumor regression, P < 0.01; for overall survival, P = 0.0644) (Fig. 4c). Notably, minuscule numbers (4 × 104) of TSCM cells were able to trigger the destruction of bulky tumors (1 cm3 containing ∼1 × 109 cells) and improve survival (TSCM cells versus TCM cells, P < 0.005; TSCM cells versus TEM cells, P < 0.005; Fig. 4c). Thus, adoptive transfer of TSCM cells in combination with tumor antigen vaccination and exogenous IL-2 produced a far more robust and therapeutically noteworthy secondary response compared with the other memory T cell subsets.

Pmel-1 naive CD8+ T cells congenically marked with Thy1.1 were primed in vitro with splenocytes pulsed with 1 μM hgp10025–33, in conjunction with 10 ng ml−1 IL-2 with or without 7 μM TWS119. Five days after antigenic stimulation, TSCM, TCM or TEM cells were sorted on the basis of phenotype (TSCM, CD44lowCD62Lhigh cells; TCM, CD44highCD62Lhigh cells; TEM, CD44highCD62Llow cells). Sublethally irradiated WT mice received 5 × 104 pmel-1 TSCM, TCM or TEM cells in conjunction with a recombinant vaccinia virus encoding hgp100 and intraperitoneal injection of IL-2. (a) Absolute numbers of adoptively transferred pmel-1 T cells (identified by CD8+Thy1.1+ lymphocytes) in the spleens of treated mice. Data are represented as means ± s.e.m. (b) Flow cytometry analysis for the expression of CD8 and Thy1.1 one month after adoptive transfer of T cell memory subsets. (c) Tumor size and survival of sublethally irradiated WT mice bearing B16 tumors established for 10 d (n = 5 for all groups) receiving 4 × 104 pmel-1 TSCM, TCM or TEM cells in conjunction with a recombinant vaccinia virus encoding hgp100 and exogenous IL-2. Data are represented as means ± s.e.m. All data shown are representative of at least two independently performed experiments.
Arrest of lymphocyte differentiation to maintain long-lived, self-renewing antigen-experienced T cells with stem cell-like properties has been postulated as the basis of the continual generation of effector T cells37, but the transcription factors that regulate this process have not been fully elucidated. Although the physiologic role of Wnt signaling in post-Thymic T cell development remains unknown, our data indicate that Wnt can regulate the stemness of CD8+ T cells by suppressing their differentiation into Teff cells. Other circumstantial evidence supports our findings. We recently reported that high levels of the Wnt transcription factors Tcf7 and Lef1 were expressed by T cells whose antigen-induced development into Teff cells had been arrested using IL-21 in the culture medium8. Conversely, Tcf7−/− mice reportedly have an increased frequency of CD44highCD62Llow TEM cells6. Paralleling our own observations in CD8+ T cells, in CD4+ T cells expression of a stabilized form of β-catenin in CD4+ T cells inhibited the proliferation and effector function of the cells in a model of inflammatory bowel disease38. Furthermore, CD4+ T cell memory precursors overexpressed Ctnnb1 and Tcf7 as well as Bcl2 and Il2rb (ref. 9). These factors were all implicated in the formation and maintenance of TSCM cells described here.
Our findings also have parallels in stem cell biology, where Wnt signaling has a pivotal role in promoting self-renewal while limiting proliferation and differentiation2,3. Hematopoietic stem cells exposed to an environment in which Wnt signaling was inhibited by overexpression of Dickkopf1 in the stem cell niche were more proliferative than those in WT mice, but they had lost their capacity for self renewal, as reflected by their inability to reconstitute hosts after serial transplants3. T cell lineage commitment to effector versus memory subsets has recently been linked to the asymmetrical segregation and inheritance of proteins that control cell fate specification39. Although the role of Wnt in the asymmetric division of CD8+ T cells is unknown, unequal localization and activities of Wnt signaling components, including β-catenin, have been implicated in cell fate specification in Caenorhabditis elegans40.
The ability to pharmacologically induce TSCM cells has considerable implications for the field of adoptive immunotherapy. The findings that small numbers of TSCM cells in conjunction with specific vaccination and IL-2 therapy can cause the regression of large established vascularized tumors might reduce the cost and complexity of highly effective therapies based on the adoptive transfer of antitumor T cells35. Using existing technology36, it may be possible to genetically engineer the human counterpart of CD8+ TSCM cells to express T cell receptors or chimeric antigen receptors while pharamacologically mimicking Wnt signaling. This might ultimately allow for the widespread application of adoptive immunotherapy based on multipotent, highly proliferative, tumor-specific TSCM cells. Finally, the modulation of Wnt signaling to induce long-term T cell memory might be crucial for T cell–based vaccines designed to target intracellular pathogens. Data indicate that protection against intracellular pathogens correlates with the induction and maintenance of TCM cell responses, but these responses have not been reproducibly obtained with current vaccination strategies24,28,34. In vivo administration of small molecule agonists of Wnt signaling might be useful to consistently achieve this goal.
Note: Supplementary information is available on the Nature Medicine website.
Methods
Mice and tumor lines. Pmel-1 TCR-transgenic mice (Jackson Laboratories) and pmel-1 ly5.1 double-transgenic mice and pmel-1 Thy1.1 double-transgenic mice were previously described16,17. We obtained B16 (H-2Db), a gp100+ murine melanoma, and MCA-205 (H-2Db), a gp100− sarcoma, from the National Cancer Institute Tumor Repository.
Antibodies, flow cytometry and cell sorting. We purchased all antibodies from BD Biosciences. We performed flow cytometry acquisition on a BD FACSCanto or BD FACSCanto II flow cytometer. We analyzed samples with FlowJo software (Tree Star). We sorted naive and T cell memory subsets with the BD FACSAria cell sorter.
Pharmacologic inhibitors of Gsk-3β and Wnt3A. We reconstituted TWS119, BIO, BIO-acetoxime, Methyl-BIO (EMD Biosciences) and Wnt3a (R&D Systems) according to the manufacturer's instructions and used them in experiments in vitro or in vivo at the various doses.
In vitro activation of CD8+ T cells. We stimulated Pmel-1 splenocytes with 1 μM hgp10025–33 peptide (Anaspec) in culture medium containing 10 ng ml−1 IL-2 (Chiron Corporation). We activated enriched CD8+ T cells isolated from the splenocytes of wild-type mice on plates coated with 0.05–2 μg ml−1 CD3-specific antibody and 1 μg ml−1 of soluble CD28-specific antibody in culture medium containing 10 ng ml−1 IL-2. We separated CD8+ T cells from non-CD8+ T cells with a MACS negative selection kit (Miltenyi Biotech).
Cytokine release and cytolytic assays. We pulsed 2.5 × 104 MCA-205 cells with 1 μM hgp10025–33 or irrelevant influenza nucleoprotein peptide and incubated them overnight with T cells at a 1:1 ratio at 37 °C. We analyzed the supernatants by mouse IFN-γ and mouse IL-2 ELISA kits (Pierce-Endogen). For 51Cr release assays, we pulsed MCA-205 target cells with 1 μM hgp10025–33 or nucleoprotein peptide and 100 μCi of 51Cr (Amersham Biosciences) for 2 h at 37 °C. We incubated labeled cells (1 × 103 cells) with T cells at ratios indicated in Figure 2c for 4 h at 37 °C. We determined maximal and spontaneous 51Cr releases by incubating 1 × 103 labeled targets in either 2% SDS or medium.
Adoptive cell transfer. We performed adoptive cell transfer, immunization with recombinant poxvirus-based vaccines and tumor experiments as previously described16,17,32. We used C57BL/6, Tcra−/−, Rag1−/− (Jackson Laboratories) and B2m−/− (Taconic) mice as recipients. We conducted all mouse experiments with the approval of the National Cancer Institute Animal Use and Care Committee.
Enumeration of adoptively transferred cells. We killed mice on day 6 (Fig. 2e) or days 3, 6 and 9 (Fig. 4a). We collected spleens and homogenized them into a single-cell suspension using the rubber end of a 3-cc syringe and a 40-μm filter cup. We enumerated samples by trypan blue exclusion and analyzed the cells by flow cytometry for CD8 and Thy.1 expression. We calculated the absolute number of pmel-1 cells by multiplying the splenocyte count by the percentage of CD8+Thy1.1+ cells.
Carboxyfluorescein succinimidyl ester proliferation assays. We used CD8+ T cells labeled with 1 μM CFSE (Molecular Probes) in adoptive experiments or stimulated them in vitro as described above.
Real-time reverse transcription-PCR. We isolated RNA with the RNeasy mini kit (Qiagen). We generated complementary DNA by reverse transcription (Applied Biosystems). We performed real-time PCR for all genes with commercially available probes and primers (Applied Biosystems) and a Prism 7900HT (Applied Biosystems). We calculated the levels of gene expression relative to the housekeeping gene encoding β-actin (Actb).
Detection of β-catenin by western blot analysis. We lysed cells in RIPA buffer (Cell Signaling Technology) with protease inhibitor. We quantified the protein concentration by Bio-Rad protein assay. We separated 20 μg of total protein on a 4–12% SDS-PAGE gel followed by standard immunoblotting with antibody to β-catenin (BD Bioscience), antibody to Gapdh (Chemicon International) and horseradish peroxidase–conjugated goat antibodies to mouse IgG (Santa Cruz Biotechnology).
Electrophoretic mobility shift assay. We isolated nuclear extracts with the nuclear extract kit (Active Motif) and then incubated them (5 μg protein) with polydeoxyinosinic-deoxycytidylic acid (1 μg μl−1 in 10 mM Tris-HCL, 1 mM EDTA, pH 8.0), 5 × Binding Buffer (Promega) and γ32P-ATP–labeled oligonucleotide probes (sense: 5′-CACAGAGAAAACAAAGCGCGCTATT-3′; antisense: 5′-AATAGCGCGCTTTGTTTTCTCTGTG-3′), containing the Tcf and Lef binding motif 5′-AACAAAG-3′. We used unlabeled oligonucleotide probes as competitors. We incubated samples for 30 min at 20 °C and loaded them onto the gel (NuPAGE 4–12% or Bis-Tris gels, Invitrogen). We blotted the gel onto a nitrocellulose membrane and exposed the membrane to X-ray film.
Statistical analyses. We compared tumor slopes with the Wilcoxon rank-sum test. We used the Kaplan-Meier method to analyze survival.
References
Luckey, C.J. et al. Memory T and memory B cells share a transcriptional program of self-renewal with long-term hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 103, 3304–3309 (2006).
Staal, F.J., Luis, T.C. & Tiemessen, M.M. WNT signalling in the immune system: WNT is spreading its wings. Nat. Rev. Immunol. 8, 581–593 (2008).
Fleming, H.E. et al. Wnt signaling in the niche enforces hematopoietic stem cell quiescence and is necessary to preserve self-renewal in vivo. Cell Stem Cell 2, 274–283 (2008).
Verbeek, S. et al. An HMG-box-containing T-cell factor required for Thymocyte differentiation. Nature 374, 70–74 (1995).
Jeannet, G. et al. Long-term, multilineage hematopoiesis occurs in the combined absence of β-catenin and γ-catenin. Blood 111, 142–149 (2008).
Schilham, M.W. et al. Critical involvement of Tcf-1 in expansion of Thymocytes. J. Immunol. 161, 3984–3991 (1998).
Willinger, T. et al. Human naive CD8 T cells down-regulate expression of the WNT pathway transcription factors lymphoid enhancer binding factor 1 and transcription factor 7 (T cell factor-1) following antigen encounter in vitro and in vivo. J. Immunol. 176, 1439–1446 (2006).
Hinrichs, C.S. et al. IL-2 and IL-21 confer opposing differentiation programs to CD8+ T cells for adoptive immunotherapy. Blood 111, 5326–5333 (2008).
Williams, M.A., Ravkov, E.V. & Bevan, M.J. Rapid culling of the CD4+ T cell repertoire in the transition from effector to memory. Immunity 28, 533–545 (2008).
Ding, S. et al. Synthetic small molecules that control stem cell fate. Proc. Natl. Acad. Sci. USA 100, 7632–7637 (2003).
Roose, J. et al. Synergy between tumor suppressor APC and the β-catenin-Tcf4 target Tcf1. Science 285, 1923–1926 (1999).
Hovanes, K et al. β-catenin-sensitive isoforms of lymphoid enhancer factor-1 are selectively expressed in colon cancer. Nat. Genet. 28, 53–57 (2001).
Mann, B. et al. Target genes of β-catenin-T cell-factor/lymphoid-enhancer-factor signaling in human colorectal carcinomas. Proc. Natl. Acad. Sci. USA 96, 1603–1608 (1999).
Katoh, M. & Katoh, M. Comparative integromics on FZD7 orthologs: conserved binding sites for PU.1, SP1, CCAAT-box and TCF/LEF/SOX transcription factors within 5′-promoter region of mammalian FZD7 orthologs. Int. J. Mol. Med. 19, 529–533 (2007).
Zeng, Y.A. & Verheyen, E.M. Nemo is an inducible antagonist of Wingless signaling during Drosophila wing development. Development 131, 2911–2920 (2004).
Overwijk, W.W. et al. Tumor regression and autoimmunity after reversal of a functionally tolerant state of self-reactive CD8+ T cells. J. Exp. Med. 198, 569–580 (2003).
Gattinoni, L. et al. Acquisition of full effector function in vitro paradoxically impairs the in vivo antitumor efficacy of adoptively transferred CD8+ T cells. J. Clin. Invest. 115, 1616–1626 (2005).
Sato, N., Meijer, L., Skaltsounis, L., Greengard, P. & Brivanlou, A.H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3–specific inhibitor. Nat. Med. 10, 55–63 (2004).
Meijer, L., Flajolet, M. & Greengard, P. Pharmacological inhibitors of glycogen synthase kinase 3. Trends Pharmacol. Sci. 25, 471–480 (2004).
Ohteki, T. et al. Negative regulation of T cell proliferation and interleukin 2 production by the serine threonine kinase GSK-3. J. Exp. Med. 192, 99–104 (2000).
Willert, K. et al. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature 423, 448–452 (2003).
Pantaleo, G. & Harari, A. Functional signatures in antiviral T-cell immunity for monitoring virus-associated diseases. Nat. Rev. Immunol. 6, 417–423 (2006).
Pearce, E.L. et al. Control of effector CD8+ T cell function by the transcription factor Eomesodermin. Science 302, 1041–1043 (2003).
Appay, V., Douek, D.C. & Price, D.A. CD8+ T cell efficacy in vaccination and disease. Nat. Med. 14, 623–628 (2008).
Zhang, Y., Joe, G., Hexner, E., Zhu, J. & Emerson, S.G. Host-reactive CD8+ memory stem cells in graft-versus-host disease. Nat. Med. 11, 1299–1305 (2005).
Sallusto, F., Geginat, J. & Lanzavecchia, A. Central memory and effector memory T cell subsets: function, generation and maintenance. Annu. Rev. Immunol. 22, 745–763 (2004).
Kaech, S.M. & Wherry, E.J. Heterogeneity and cell-fate decisions in effector and memory CD8+ T cell differentiation during viral infection. Immunity 27, 393–405 (2007).
Klebanoff, C.A., Gattinoni, L. & Restifo, N.P. CD8+ T-cell memory in tumor immunology and immunotherapy. Immunol. Rev. 211, 214–224 (2006).
Tough, D.F. & Sprent, J. Turnover of naive- and memory-phenotype T cells. J. Exp. Med. 179, 1127–1135 (1994).
Surh, C.D. & Sprent, J. Homeostatic T cell proliferation: how far can T cells be activated to self-ligands? J. Exp. Med. 192, F9–F14 (2000).
Murali-Krishna, K. et al. Persistence of memory CD8 T cells in MHC class I–deficient mice. Science 286, 1377–1381 (1999).
Wrzesinski, C. et al. Hematopoietic stem cells promote the expansion and function of adoptively transferred antitumor CD8 T cells. J. Clin. Invest. 117, 492–501 (2007).
Klebanoff, C.A. et al. Central memory self/tumor-reactive CD8+ T cells confer superior antitumor immunity compared with effector memory T cells. Proc. Natl. Acad. Sci. USA 102, 9571–9576 (2005).
Wherry, E.J. et al. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat. Immunol. 4, 225–234 (2003).
Gattinoni, L., Powell, D.J. Jr., Rosenberg, S.A. & Restifo, N.P. Adoptive immunotherapy for cancer: building on success. Nat. Rev. Immunol. 6, 383–393 (2006).
Morgan, R.A. et al. Cancer regression in patients after transfer of genetically engineered lymphocytes. Science 314, 126–129 (2006).
Fearon, D.T., Manders, P. & Wagner, S.D. Arrested differentiation, the self-renewing memory lymphocyte and vaccination. Science 293, 248–250 (2001).
Ding, Y., Shen, S., Lino, A.C., Curotto de Lafaille, M.A. & Lafaille, J.J. β-catenin stabilization extends regulatory T cell survival and induces anergy in nonregulatory T cells. Nat. Med. 14, 162–169 (2008).
Chang, J.T. et al. Asymmetric T lymphocyte division in the initiation of adaptive immune responses. Science 315, 1687–1691 (2007).
Mizumoto, K. & Sawa, H. Two βs or not two βs: regulation of asymmetric division by β-catenin. Trends Cell Biol. 17, 465–473 (2007).
Acknowledgements
This research was supported by the Intramural Research Program of the US National Institutes of Health, National Cancer Institute, Center for Cancer Research. We would like to thank S.A. Rosenberg, C.A. Klebanoff and S. Kerkar for critical review of the manuscript and A. Mixon and S. Farid of the Flow Cytometry Unit for Flow Cytometry analyses and sorting. This study was done in partial fulfillment of a PhD in Biochemistry (to D.C.P.) at the George Washington University, Washington, DC.
Author information
Authors and Affiliations
Contributions
L.G. designed, performed and analyzed experiments and wrote the paper; X.-S.Z. D.C.P., Y.J., C.S.H., Z.Y., C.W., A.B., L.C., L.M.G., C.M.P. and P.M. performed experiments; and N.P.R. designed experiments and wrote the paper.
Corresponding authors
Supplementary information
Supplementary Text and Figures
Supplementary Figs. 1–8 (PDF 832 kb)
Rights and permissions
About this article
Cite this article
Gattinoni, L., Zhong, XS., Palmer, D. et al. Wnt signaling arrests effector T cell differentiation and generates CD8+ memory stem cells. Nat Med 15, 808–813 (2009). https://doi.org/10.1038/nm.1982
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nm.1982
This article is cited by
-
IL-10-expressing CAR T cells resist dysfunction and mediate durable clearance of solid tumors and metastases
Nature Biotechnology (2024)
-
WNT/β-catenin regulatory roles on PD-(L)1 and immunotherapy responses
Clinical and Experimental Medicine (2024)
-
Eribulin promotes proliferation of CD8+ T cells and potentiates T cell-mediated anti-tumor activity against triple-negative breast cancer cells
Breast Cancer Research and Treatment (2024)
-
FOXO1 is a master regulator of memory programming in CAR T cells
Nature (2024)
-
New insights into the stemness of adoptively transferred T cells by γc family cytokines
Cell Communication and Signaling (2023)
