
Abstract
Replying to: A. S. L. Rodrigues Nature 450, 10.1038/nature06374 (2007)
Rodrigues1 criticizes our demonstration2 of low congruence in the global distributions of rare and threatened vertebrates on the grounds that we excluded locations where species counts were zero from our analyses. In practice, this makes no substantive difference to our conclusions. Some sample locations are not inhabited by any organisms of interest: such locations can inflate measures of covariation and association because their values for parameters of interest (in this case, zero counts of species) are identical. This bias has long been known (as ‘the double-zero problem’3); many ecological techniques and studies exclude double-zero data for this reason.
Similar content being viewed by others


Main
We are happy to clarify that we excluded double-zero data. We concur with Rodrigues1 that including double-zero data increases the observed correlation coefficient (r). Indeed, it cannot fail to do so: r becomes a function of the ratio of double-zero to non-zero data. For example, given our 19,564 terrestrial grid cells, if fewer than 13,043 cells (two-thirds of the world’s surface) are filled with perfectly negatively correlated richnesses and the rest with double zeroes, a positive correlation is observed. In any such case, including double-zero data would be to predetermine the result: our comparison of rare mammals and amphibians involved only 3,184 non-zero cells. This is sufficient justification for our choice of method.
Treating all cells as informative, as Rodrigues proposes, becomes analogous to the analysis of binary presence–absence data: it examines associations between cells that have any species of interest, rather than the relative diversities of those cells. Such questions are better addressed by explicitly converting species counts to binary presence–absence data before analysis. The analyses would rely more on the veracity of data on absence, and the metrics of association will depend on the relative abundance of zero to non-zero data.
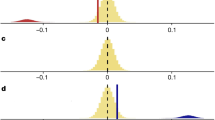
Rodrigues also criticizes our description of network performance; our two contrasting methods are shown in Fig. 1. Our definition equates performance with the trade-off between network size and species capture. Rodrigues prefers a measure that quantifies the effectiveness of a network given its size. Both measures describe different aspects of reserve network performance. We prefer our definition in this case because we regard the comparison of absolute network sizes to be important. For example, under Rodrigues’ definition, the optimal network for rare mammals and the global 200 scheme (points M and G in Rodrigues’ Fig. 1; ref. 1) have an effectiveness of around 70% and 50%, respectively, despite differing in area by more than an order of magnitude.

Rodrigues’ preferred measure1 (blue) evaluates the difference between the maximum possible (upper grey line) and mean random (lower grey line) species capture, given the size of the network in question (black dot). Our preferred measure2 (red) evaluates the ratio of species captured to the number of cells in the network.
We agree that good progress has been made in conservation planning and make no claim that the prospects from existing plans are dismal. We do argue, however, that the efficiency of conservation schemes can be improved by detailing where species across multiple higher taxa are found.
References
Rodrigues, A. S. L. Effective global conservation strategies. Nature 450 10.1038/nature06374 (2007)
Grenyer, R. et al. Global distribution and conservation of rare and threatened vertebrates. Nature 444, 93–96 (2006)
Legendre, L. & Legendre, P. Numerical Ecology 2nd edn (Elsevier Science, Amsterdam, 1998)
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Grenyer, R., Orme, C., Davies, T. et al. Grenyer et al. reply. Nature 450, E20 (2007). https://doi.org/10.1038/nature06375
Issue Date:
DOI: https://doi.org/10.1038/nature06375
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
