
Abstract
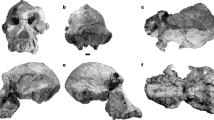
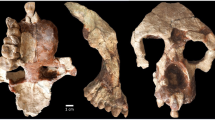
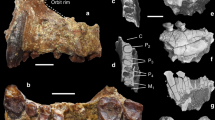
The search for the earliest fossil evidence of the human lineage has been concentrated in East Africa. Here we report the discovery of six hominid specimens from Chad, central Africa, 2,500 km from the East African Rift Valley. The fossils include a nearly complete cranium and fragmentary lower jaws. The associated fauna suggest the fossils are between 6 and 7 million years old. The fossils display a unique mosaic of primitive and derived characters, and constitute a new genus and species of hominid. The distance from the Rift Valley, and the great antiquity of the fossils, suggest that the earliest members of the hominid clade were more widely distributed than has been thought, and that the divergence between the human and chimpanzee lineages was earlier than indicated by most molecular studies.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others
References
Dart, R. A. Australopithecus africanus, the man-ape of South Africa. Nature 115, 195–199 (1925)
Kortlandt, A. New Perspectives on Ape and Human Evolution 9–100 (Stichting voor Psychobiologie, Amsterdam, 1972)
Coppens, Y. Le singe, l'Afrique et l'Homme (Fayard, Paris, 1983)
Leakey, M. G. et al. New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433–440 (2001)
Leakey, M. G., Feibel, C. S., McDougall, I. & Walker, A. C. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature 376, 565–571 (1995)
White, T. D., Suwa, G. & Asfaw, B. Australopithecus ramidus, a new species of hominid from Aramis, Ethiopia. Nature 371, 306–312 (1994)
Haile-Selassie, Y. Late Miocene hominids from the Middle Awash, Ethiopia. Nature 412, 178–181 (2001)
Senut, B. et al. First hominid from the Miocene (Lukeino Formation, Kenya). C. R. Acad. Sci. Paris 332, 137–144 (2001)
Deino, A. L., Tauxe, L., Monaghan, M. & Hill, A. 40Ar/39Ar geochronology and paleomagnetic stratigraphy of the Lukeino and lower Chemeron Formations at Tabarin and Kapcheberek, Tugen Hills, Kenya. J. Hum. Evol. 42, 117–140 (2002)
Brunet, M. et al. The first australopithecine 2500 kilometres west of the Rift Valley (Chad). Nature 378, 273–275 (1995)
Brunet, M. et al. Australopithecus bahrelghazali, une nouvelle espèce d'Hominidé ancien de la région de Koro Toro (Tchad). C. R. Acad. Sci. Paris 322, 907–913 (1996)
Vignaud, P. et al. Geology and palaeontology of the Upper Miocene Toros-Menalla hominid locality, Chad. Nature 418, 152–155 (2002)
McDougall, I. & Feibel, C. S. Numerical age control for the Miocene–Pliocene succession at Lothagam, a hominoid-bearing sequence in the northern Kenya Rift. J. Geol. Soc. Lond. 156, 731–745 (1999)
Ishida, H. & Pickford, M. A new Late Miocene hominoid from Kenya: Samburupithecus kiptalami gen. et sp. nov. C. R. Acad. Sci. Paris 325, 823–829 (1998)
Bonis, L. de & Koufos, G. D. The face and the mandible of Ouranopithecus macedoniensis: description of new specimens and comparisons. J. Hum. Evol. 24, 469–491 (1993)
Pilbeam, D. New hominoid skull material from the Miocene of Pakistan. Nature 295, 232–234 (1982)
Kordos, L. & Begun, D. R. A new cranium of Dryopithecus from Rudabanya, Hungary. J. Hum. Evol. 41, 689–700 (2001)
Tobias, P. V. The Cranium and Maxillary Dentition of Australopithecus (Zinjanthropus) boisei, Olduvai Gorge (Cambridge Univ. Press, London, 1967)
Kimbel, W. H., White, T. D. & Johanson, D. C. Cranial morphology of Australopithecus afarensis; A comparative study based on a composite reconstruction of the adult skull. Am. J. Phys. Anthropol. 64, 337–388 (1984)
Ward, C. V., Leakey, M. G. & Walker, A. Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. J. Hum. Evol. 41, 235–368 (2001)
Kimbel, W. H., Johanson, D. C. & Rak, Y. The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia. Nature 368, 449–451 (1994)
Wood, B. Koobi Fora Research Project: Hominid Cranial Remains Vol. 4 (Clarendon, Oxford, 1991)
Kumar, S. & Hedges, B. A molecular time scale for vertebrate evolution. Nature 392, 917–920 (1998)
Pilbeam, D. in The Primate Fossil Record (ed. Hartwig, W.) 303–310 (Columbia Univ. Press, New York, 2002)
Kimbel, W. H. Systematic assessment of a maxilla of Homo from Hadar, Ethiopia. Am. J. Phys. Anthop. 103, 235–262 (1997)
Uchida, A. Intra-species Variation among the Great-Apes: Implications for Taxonomy of Fossil Hominoids Thesis, Harvard Univ. (1992)
Lockwood, C. A., Kimbel, W. H. & Johanson, D. C. Temporal trends and metric variation in the mandibles and dentition of A. afarensis. J. Hum. Evol. 39, 23–55 (2000)
Lockwood, C. A. Sexual dimorphism in the face of Australopithecus africanus. Am. J. Phys. Anthropol. 108, 97–127 (1999)
Acknowledgements
We thank the Chadian Authorities (Ministère de l'Education Nationale de l'Enseignement Supérieur et de la Recherche, Université de N'djaména, CNAR). We extend gratitude for their support to the French Ministries, Ministère Français de l'Education Nationale (Faculté des Sciences, Université de Poitiers), Ministère de la Recherche (CNRS), Ministère des Affaires Etrangères (Direction de la Coopération Scientifique, Universitaire et de Recherche, Paris, and SCAC Ambassade de France à N'djaména), to the Région Poitou-Charentes, the Département de la Vienne, the Association pour le Prix scientifique Philip Morris, and also to the Armée Française (MAM and Epervier) for logistic support. For giving us the opportunity to work with their collections, we are grateful to the National Museum of Ethiopia, the National Museum of Kenya, the Peabody Museum and Harvard University, the Institute of Human Origins and the University of California. Special thanks to Scanner-IRM Poitou Charentes (P. Chartier and F. Perrin), for industrial scanner to EMPA (A. Flisch Ing. HTL) and to Multimedia Laboratorium-Computer Department, University of Zurich-Irchel (P. Stucki). Many thanks to all our colleagues and friends for their help and discussion, and particularly to F. Clark Howell, A. Garaudel, Y. Haile-Selassie, D. Johanson, W. Kimbel, M. G. Leakey, D. Lieberman, R. Macchiarelli, M. Pickford, B. Senut, G. Suwa, T. White and Lubaka. We especially thank all the other MPFT members who joined us for the field missions, and V. Bellefet, S. Riffaut and J.-C. Bertrand for technical support. We are most grateful to G. Florent for administrative guidance. We dedicate this article to J. D. Clark. All authors are members of the MPFT.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing financial interests.
Rights and permissions
About this article
Cite this article
Brunet, M., Guy, F., Pilbeam, D. et al. A new hominid from the Upper Miocene of Chad, Central Africa. Nature 418, 145–151 (2002). https://doi.org/10.1038/nature00879
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/nature00879
This article is cited by
-
Seven-million-year-old femur suggests ancient human relative walked upright
Nature (2022)
-
Using primate models to study the evolution of human locomotion: concepts and cases
BMSAP (2014)
-
The evolution of the upright posture and gait—a review and a new synthesis
Naturwissenschaften (2010)
-
Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens
Applied Physics A (2006)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
