
Abstract
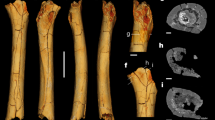
Bipedalism has traditionally been regarded as the fundamental adaptation that sets hominids apart from other primates. Fossil evidence demonstrates that by 4.1 million years ago1, and perhaps earlier2, hominids exhibited adaptations to bipedal walking. At present, however, the fossil record offers little information about the origin of bipedalism, and despite nearly a century of research on existing fossils and comparative anatomy, there is still no consensus concerning the mode of locomotion that preceded bipedalism3,4,5,6,7,8,9,10. Here we present evidence that fossils attributed to Australopithecus anamensis (KNM-ER 20419)11 and A. afarensis (AL 288-1)12 retain specialized wrist morphology associated with knuckle-walking. This distal radial morphology differs from that of later hominids and non-knuckle-walking anthropoid primates, suggesting that knuckle-walking is a derived feature of the African ape and human clade. This removes key morphological evidence for a Pan–Gorilla clade, and suggests that bipedal hominids evolved from a knuckle-walking ancestor that was already partly terrestrial.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


References
Leakey, M. G., Fiebel, C. S., McDougall, I. & Walker, A. C. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature 376, 565–571 (1995).
White, T. D., Suwa, G. & Asfaw, B. Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia. Nature 371, 306– 312 (1994).
Keith, A. Man's posture: its evolution and disorders. Br. Med. J. 1, 451–454, 499–502, 545–548, 587–590, 624–626, 669–672 (1923).
Washburn, S. L. Behavior and the origin of man. Proc. R. Anthropol. Inst. Gr. Br. Ireland 3, 21–27 ( 1967).
Marzke, M. W. Origin of the human hand. Am. J. Phys. Anthropol. 34 , 61–84 (1971).
Tuttle, R. H. Darwin's apes, dental apes, and the descent of man: normal science in evolutionary anthropology. Curr. Anthropol. 15, 389– 398 (1974).
Stern, J. T. Jr Before bipedality. Yrbk Phys. Anthropol. 19, 59–68 (1975).
Fleagle, J. G. et al. Climbing: a biomechanical link with brachiation and bipedalism. Symp. Zool. Soc. Lond. 48, 359– 373 (1981).
Rose, M. D. in Origine(s) de la Bipédie Chez les Hominidés (eds Coppens, Y. & Senut, B.) 37–48 (Centre National de la Recherche Scientifique, Paris, 1991).
Gebo, D. L. Climbing, brachiation, and terrestrial quadrupedalism: historical precursors of hominid bipedalism. Am. J. Phys. Anthropol. 101, 55–92 (1996).
Heinrich, R. E., Rose, M. D., Leakey, R. E. & Walker, A. C. Hominid radius from the Middle Pliocene of Lake Turkana, Kenya. Am. J. Phys. Anthropol. 92, 139–148 (1993).
Johanson, D. C. et al. Morphology of the Pliocene partial hominid skeleton (AL288-1) from the Hadar Formation, Ethiopia. Am. J. Phys. Anthropol. 57, 403–451 (1982).
Jenkins, F. A. Jr & Fleagle, J. G. in Primate Functional Morphology and Evolution (ed. Tuttle, R. H.) 213–231 (Mouton, The Hague, 1975).
Tuttle, R. H. Knuckle-walking and the evolution of hominoid hands. Am. J. Phys. Anthropol. 26, 171–206 ( 1967).
Tuttle, R. H. Quantitative and functional studies on the hands of the anthropoidea: I. The Hominoidea. J. Morphol. 128, 309– 363 (1969).
Susman, R. L. & Stern, J. T. Jr Telemetered electromyography of flexor digitorum profundus and flexor digitorum superficialis in Pan troglodytes and implications for interpretation of the O.H. 7 hand. Am. J. Phys. Anthropol. 50, 565–574 (1979).
Schmitt, D. Forelimb mechanics as a function of substrate type during quadrupedalism in two anthropoid primates. J. Hum. Evol. 26, 441–457 (1994).
Richmond, B. G. Ontogeny and Biomechanics of Phalangeal Form in Primates. Thesis, State Univ. New York at Stony Brook (1998).
Stern, J. T. Jr & Susman, R. L. The locomotor anatomy of Australopithecus afarensis. Am. J. Phys. Anthropol. 60, 279–317 ( 1983).
Susman, R. L. Comparative and functional morphology of hominoid fingers. Am. J. Phys. Anthropol. 50, 215–235 (1979).
Begun, D. R. Knuckle-walking ancestors. Science 259, 294 (1993).
Andrews, P. J. & Martin, L. B. Cladistic relationships of extant and fossil hominoids. J. Hum. Evol. 16, 101–118 (1987).
Begun, D. R. Miocene fossil hominids and the chimp-human clade. Science 257, 1929–1933 (1992).
Ruvolo, M. Molecular phylogeny of the hominoids: inferences from multiple independent DNA sequence data sets. Mol. Biol. Evol. 14, 248–265 (1997).
Dainton, M. & Macho, G. A. Did knuckle walking evolve twice? J. Hum. Evol. 36, 171– 194 (1999).
Hunt, K. D. The postural feeding hypothesis: an ecological model for the origin of bipedalism. S. Afr. J. Sci. 9, 77– 90 (1996).
Hewes, G. W. Food transport and the origin of hominid bipedalism. Am. Anthropol. 63, 687–710 ( 1961).
Lovejoy, C. O. The origin of man. Science 211, 341– 350 (1981).
Napier, J. R. & Davis, P. The forelimb skeleton and associated remains of Proconsul africanus. Foss. Mamm. Afr. 16, 1–70 (1959).
Grine, F. E. & Susman, R. L. Radius of Paranthropus robustus from Member 1, Swartkrans Formation, South Africa. Am. J. Phys. Anthropol. 84, 229–248 (1991).
Acknowledgements
We thank L. Aiello, E. Delson, B. Demes, J. Fleagle, F. Grine, W. Jungers, S. Larson, D. Lieberman, O. Pearson, D. Pilbeam, J. Polk, E. Sarmiento, R. Susman and B. Wood for providing valuable comments; F. Grine, L. Gordon, R. Thorington, R. Potts, A. Walker, and C. Ward for providing access to specimens in their care; and The Henry Luce Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Richmond, B., Strait, D. Evidence that humans evolved from a knuckle-walking ancestor. Nature 404, 382–385 (2000). https://doi.org/10.1038/35006045
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/35006045
This article is cited by
-
Mittel- bis langfristige, funktionelle Ergebnisse nach Hemiresektion-Interpositions-Arthroplastik (nach Bowers) des distalen Radioulnargelenks
Die Orthopädie (2022)
-
The biomechanical importance of the scaphoid-centrale fusion during simulated knuckle-walking and its implications for human locomotor evolution
Scientific Reports (2020)
-
Femoral ontogeny in humans and great apes and its implications for their last common ancestor
Scientific Reports (2018)
-
Anatomie und Biomechanik des distalen Radioulnargelenks
Der Orthopäde (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
