
Abstract
OBJECTIVE: Based on the inhibitory effects of teasaponin on pancreatic lipase activity in vitro, this study was performed to clarify whether teasaponin prevented obesity induced in mice by a high-fat diet for 11 weeks.
DESIGN: For in vitro experiments, assay for the inhibitory effects of teasaponin on pancreatic lipase activity was performed by measuring the rate of release of oleic acid from triolein in an assay system using triolein emulsified with lecithin, gum arabic, Triton X-100 or 4-methylumbelliferyloleate. For in vivo experiments, female ICR mice were fed a high-fat diet with or without 0.5% teasaponin for 11 weeks.
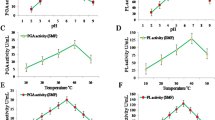
RESULTS: Teasaponin competitively inhibited the hydrolysis of triolein emulsified with lecithin, gum arabic, Triton X-100 or 4-methylumbelliferyloleate. Teasaponin inhibited the elevations of plasma triacylglycerol levels 3, 4 and 5 h after oral administration of lipid emulsion containing corn oil. Teasaponin suppressed the increases in body, parametrial adipose tissue weights and diameter in adipose cell size induced by a high-fat diet. Furthermore, feeding a high-fat diet plus teasaponin had no effect on stool frequency and content, but significantly increased triacylglycerol contents in feces as compared to feeding a high-fat diet.
CONCLUSIONS: The anti-obesity effects of teasaponin in high-fat diet-treated mice may be partly mediated through delaying the intestinal absorption of dietary fat by inhibiting pancreatic lipase activity.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


References
Kimura Y, Okuda H, Mori K, Okuda T, Arichi S . Effects of extracts of various kinds of tea on lipid metabolic injury in rats fed peroxidized oil J Jpn Soc Nutri Food Sci 1984 37: 223–232 (In Japanese).
Okuda T, Kimura Y, Yoshida T, Hatano T, Okuda H, Arichi S . Studies on the activities of tannins and related compounds from medicinal plants and drugs. I. Inhibitory effects on lipid peroxidation in mitochondria and microsomes of liver Chem Pharm Bull 1983 31: 1625–1631.
Han L-K, Takaku T, Li J, Kimura Y, Okuda H . Anti-obesity action of oolong tea Int J Obes Relat Metab Disord 1999 23: 98–105.
Kitagawa I, Hori K, Motozawa T, Murakami T, Yoshikawa M . Structures of new acylated oleanene-type triterpene oligoglycosides, theasaponins E1 and E2, from the seeds of tea plant, Camellia sinensis (L.) O. Kuntze Chem Pharm Bull 1998 46: 1901–1906.
Hirsch J, Gallian E . Methods for the determination of adipose cell size in man and animals J Lipid Res 1968 9: 110–119.
Akagi M, Fukuishi N, Kan T, Sagesaka YM, Akagi R . Anti-allergic effect of tea-leaf saponin (TLS) from tea leaves (Camellia sinensis var. sinensis) Biol Pharm Bull 1997 20: 565–567.
Sagesaka-Mitane Y, Sugiura T, Miwa Y, Yamaguchi K, Kyuki K . Effect of tea-leaf saponin on blood pressure of spontaneously hypertensive rats Yakugaku zasshi 1996 116: 388–395.
Sagesaka YM, Uemura T, Suzuki Y, Sugiura T, Yoshida M, Yamaguchi K, Kyuki K . Antimicrobial and anti-inflammtory actions of tea-leaf saponin Yakugaku zasshi 1996 116: 238–243.
Verger R . Pancreatic lipase. In: Borgstrom B and Brockman HL (eds). Lipase Elsevier: Amsterdam 1984 83–150.
Drent ML, Popp-Snijders C, Ader HJ, Jansen JB, van der Veen EA . Lipase inhibition and hormonal status, body composition and gastrointestinal processing of a liquid high-fat mixed meal in moderately obese subjects Obes Res 1995 3: 573–581.
Drent ML, van der Veen EA . First clinical studies with orlistat: a short review Obes Res 1995 3: (Suppl 4): 623s–625s.
Drent ML, Larsson I, William Olsson T, Quaade F, Czubayko F, von Bergmann K, Strobel W, Sjostrom L, van der Veen EA . Orlistat (Ro 18-0647), a lipase inhibitor, in the treatment of human obesity: a multiple dose study Int J Obes Relat Metab Disord 1995 19: 221–226.
Hauptman JB, Jeunet FS, Hartmann D . Initial studies in humans with the novel gastrointestinal lipase inhibitor Ro 18-0647 (tetrahydrolipstatin) Am J Clin Nutr 1992 55: (Suppl 1): 309s–313s.
Kalivianakis M, Elstrodt J, Havinga R, Kuipers F, Stellaard F, Sauer PJ, Vonk RJ, Verkade HJ . Validation in an animal model of the carbon 13-labeled mixed triglyceride breath test for the detection of intestinal fat malabsorption J Pediatr 1999 135: 444–450.
Han L-K, Kimura Y, Okuda H . Reduction in fat storage during chitin–chitosan treatment in mice fed a high-fat diet Int J Obes Relat Metab Disord 1999 23: 174–178.
Hill JO, Lin D, Yakubu F, Peters JC . Development of dietary obesity in rats: influence of amount and composition of dietary fat Int J Obes Relat Metab Disord 1992 16: 321–333.
Lemonnier D . Effect of age, sex and site on the cellularity of the adipose tissue in mice and rats rendered obese by a high-fat diet J Clin Invest 1972 51: 2907–2915.
Obst BE, Schemmel RA, Czajka-Narins D, Merkel R . Adipocyte size and number in dietary obesity resistant and susceptible rats Am J Physiol 1981 240: E47–E53.
Segal R, Mansour M, Zaitschek DV . Effect of ester groups on the haemolytic action of some saponins and sapogenins Biochem Pharmac 1966 15: 1411–1416.
Abe H, Sakaguchi M, Konishi H, Tani T, Arichi S . The effects of saikosaponins on biological membranes. 1. The relationship between the structures of saikosaponins and haemolytic activity Planta Med 1978 34: 160–166.
Abe H, Sakaguchi M, Anno M, Arichi S . Erythrocyte membrane stabilization by plant saponins and sapogenins Naunyn Schmiedebergs Arch Pharmac 1981 316: 262–265.
Kim Yu A, Akoev VR, Elemesov RE . Hyperosmotic hemolysis and antihemolytic activity of the saponin fraction and triterpene glycosides from Panax ginseng C. A. Meyer. Membr Cell Biol 2000 14: 237–251.
Nguyen TD, Villard PH, Barlatier A, Elsisi AE, Jouve E, Duc NM, Sauze C, Durand A, Lacarelle B . Panax vietnamensis protects mice against carbon tetrachloride-induced hepatotoxicity without any modification of CYP2E1 gene expression Planta Med 2000 66: 714–719.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Han, LK., Kimura, Y., Kawashima, M. et al. Anti-obesity effects in rodents of dietary teasaponin, a lipase inhibitor. Int J Obes 25, 1459–1464 (2001). https://doi.org/10.1038/sj.ijo.0801747
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.ijo.0801747
