
Abstract
For insects, temperature is a major environmental variable that can influence an individual's behavioral activities and fitness. Drosophila melanogaster is a cosmopolitan species that has had great success in adapting to and colonizing diverse thermal niches. This adaptation and colonization has resulted in complex patterns of genetic variation in thermotolerance phenotypes in nature. Although extensive work has been conducted documenting patterns of genetic variation, substantially less is known about the genomic regions or genes that underlie this ecologically and evolutionarily important genetic variation. To begin to understand and identify the genes controlling thermotolerance phenotypes, we have used a mapping population of recombinant inbred (RI) lines to map quantitative trait loci (QTL) that affect variation in both heat- and cold-stress resistance. The mapping population was derived from a cross between two lines of D. melanogaster (Oregon-R and 2b) that were not selected for thermotolerance phenotypes, but exhibit significant genetic divergence for both phenotypes. Using a design in which each RI line was backcrossed to both parental lines, we mapped seven QTL affecting thermotolerance on the second and third chromosomes. Three of the QTL influence cold-stress resistance and four affect heat-stress resistance. Most of the QTL were trait or sex specific, suggesting that overlapping but generally unique genetic architectures underlie resistance to low- and high-temperature extremes. Each QTL explained between 5 and 14% of the genetic variance among lines, and degrees of dominance ranged from completely additive to partial dominance. Potential thermotolerance candidate loci contained within our QTL regions are identified and discussed.
Similar content being viewed by others


Introduction
For insects, variation in temperature is one of the most important environmental variables that influences the distribution of species in nature (Coussins and Bowler, 1987; Leather et al, 1993; Clarke, 1996). Temperature can vary on a daily, seasonal, or spatial scale (Gibbs et al, 2003) and this variation can have major impacts on an individual's fitness (Hoffmann et al, 2003; Rohmer et al, 2004) as well as its daily behavioral activities (Hoffmann and Parsons, 1991; Gilchrist and Huey, 1999; Gilbert and Huey, 2001; David et al, 2003). Thus, variation in environmental temperature will generally impose stress upon the organism, which may result in the evolution of adaptive genetic mechanisms to cope with temperature extremes in nature (Hoffmann and Parsons, 1991). Drosophila melanogaster is a species that has been very successful in its ability to adapt to novel thermal challenges (David and Capy, 1988) and is currently a widely distributed cosmopolitan species that has successfully colonized and adapted within numerous thermal environments in nature (David and Capy, 1988; Lachaise et al, 1988). Populations of D. melanogaster currently persist on all of the continents and are a part of a network of locally adapted populations that exhibit complex patterns of phenotypic and genetic variation both within and among populations (Hoffmann et al, 2003).
One very important class of locally adapted phenotypes in D. melanogaster is the thermotolerance phenotypes, that is the phenotypes contributing to resistance to high- and low-temperature extremes (Hoffmann et al, 2003). Abundant naturally segregating genetic variance for resistance to high- and low-temperature extremes has been documented in Drosophila by response to artificial or natural laboratory selection (Morrison and Milkman, 1978; Tucic, 1979; Cavicchi et al, 1995; Loeschcke and Krebs, 1996; Gilchrist et al, 1997; Bubli et al, 1998; Gilchrist and Huey, 1999; Norry et al, 2004; Anderson et al, 2005); quantification of genetic variation along clines of resistance to heat and cold stress (Gilbert and Huey 2001; Hoffmann et al, 2002; Ayrinhac et al, 2004; Kimura, 2004); and association of phenotypic variation in thermotolerance with molecular polymorphism or expression of candidate genes affecting thermal-stress resistance (McColl et al, 1996; Krebs and Feder, 1997; Dahlgaard et al, 1998; Anderson et al, 2003). Although these studies have shown that substantial genetic variation exists for heat- and cold-stress resistance phenotypes in natural and laboratory Drosophila populations, it is essential to identify the individual genetic loci underlying this complex variation in order to understand the evolutionary genetic mechanisms influencing adaptation to thermal environments.
To date, studies exploring the genetic architecture of thermotolerance phenotypes have primarily focused on candidate genes known to be involved in the heat-stress response (eg Heat shock proteins (Hsps)). For example, Hsp70 (Feder and Krebs, 1998; Dahlgaard et al, 1998; Bettencourt et al, 2002), Hsp68 (McColl et al, 1996), Hsr-omega (McColl et al, 1996; McColl and McKechnie, 1999; Anderson et al, 2003), and heat-shock factor (Hsf) (Lerman and Feder, 2001) have all been implicated in the resistance to heat stress, through the analysis of allele frequency changes in response to selection on thermotolerance, the analysis of clinal variation in allele frequencies, the analysis of the change in gene expression among populations with divergent heat-stress profiles, or through the analysis of the change in the resistance phenotypes with increased copy number of the candidate locus. In addition, the methusalah (mth) longevity gene has been implicated in resistance to various stressors including heat stress (Lin et al, 1998). Substantially less work has been conducted for cold-stress resistance, possibly because our knowledge of the relevant candidate genes and pathways is limited. Goto (2000, 2001) showed that Senescence marker protein-30 (Smp-30) and Frost (Fst) are both upregulated in response to cold acclimation or cold stress. Anderson et al (2003) demonstrated an association between clinal molecular variation in Hsr-omega and cold-stress resistance. Finally, Greenberg et al (2003) implicated desaturase 2 (desat2) in cold resistance by identifying significant allelic differences between Cosmopolitan and Zimbabwe forms of the gene in a site-directed gene replacement study.
All of the studies highlighted above focused on the role of a small sample of candidate genes in some aspect of resistance to thermal stress. However, an alternative to candidate gene studies is to identify genomic regions (quantitative trait loci (QTL)) influencing the trait between lines with divergent thermotolerance phenotypes. QTL studies have the great advantage of not imposing any a priori prediction on the classes of genes identified in a genome scan, but they also have a distinct disadvantage in that nearly all QTL contain multiple genes. Although the power of the QTL mapping approach is great (Mackay, 2001), it has not been used to genetically dissect any aspect of thermotolerance until recently. Norry et al (2004) used a QTL mapping procedure to identify four QTL (one on chromosome 2, three on chromosome 3) contributing to the variation between lines selected for increased and decreased high-temperature knockdown time. Each QTL identified in this study identified a large genomic region, but each of the QTL contained plausible candidate loci affecting heat resistance, including numerous Hsp genes, Hsr-omega, and Trap1 (Norry et al, 2004).
Here, we present the results of a genome scan for QTL influencing variation for both heat- and cold-stress resistance using a backcross design and composite interval mapping in a panel of recombinant inbred (RI) lines derived from a cross between two strains of D. melanogaster, Oregon-R and 2b (Nuzhdin et al, 1997). Although Oregon-R and 2b were not selected for extreme thermotolerance phenotypes, they are significantly different for both heat- and cold-stress resistance phenotypes (Figure 1). Furthermore, we acknowledge that Oregon-R and 2b represent a limited sample of the genetic variation that may underlie thermotolerance phenotypes in nature; however, localizing QTL in these divergent lines does represent a valuable primary step in understanding the genetic architecture underlying these phenotypes. Using this genome scan for QTL we ask the following questions: (i) What genomic regions influence these ecologically important phenotypes? (ii) What are the allelic effects of these QTL (ie additive, dominance, epistatic effects)? (iii) Are there overlapping or unique genetic architectures for each thermotolerance phenotype or sex? (iv) Do our QTL contain likely thermotolerance candidate loci? (v) To what extent do the locations of QTL affecting variation in heat stress resistance match those detected by Norry et al (2004)?

Parental line chill-coma recovery (a) and heat-stress survival (b) profiles. Males are represented by solid lines and females are represented by dashed lines. The Oregon flies are represented by black lines and the 2b flies are represented by gray lines. The horizontal dotted line represents the 50% recovery/survival time point. Solid arrows represent the average 50% recovery/survival time point of the two parental lines as determined by regression (see Materials and methods).
Materials and methods
Drosophila stocks
Two unrelated inbred parental lines, Oregon (Lindsley and Zimm, 1992) and 2b (Pasyukova and Nuzhdin, 1993), and a population of 81 RI lines derived from these parental lines were used in this experiment. The generation of these RI lines has been described previously (Nuzhdin et al, 1997); so only a brief summary is provided here. Oregon is a standard inbred lab stock that has not been selected for any extreme phenotypes, whereas 2b is an inbred lab stock derived from a selection experiment for decreased male-mating activity (Kaidanov, 1990). The RI lines were established from an initial cross between the Oregon and 2b lines. Offspring from this cross were backcrossed to 2b, and these progeny were randomly mated for four generations to increase the amount of recombination. The backcross to 2b was carried out because the 2b strain is generally less vigorous than the Oregon strain and the backcross helped ensure that the proportion of 2b and Oregon alleles were equally represented within each RI line. The population of RI lines was subsequently created by conducting full-sib mating for 25 generations. Following the 25 generations of inbreeding, the surviving lines were maintained by mass transfer each generation. These RI lines have been maintained since 1996 and hence spontaneous mutations will have occurred. The contribution of new mutations was minimized by maintaining the lines in relatively large population sizes by mass transfer. Further, the input of new mutational variance will be random and will reduce our ability to detect QTL initially segregating between these lines. These RI lines were chosen for the practical reasons that the genotypes of the molecular markers had previously been determined and other life-history QTL have previously been mapped in this population. Although the genetic variation between the Ore and 2b parental lines is a highly restricted sample of naturally occurring variation in thermotolerance, the QTL analyses presented in this study represent an important first step in genetic dissection of heat and cold tolerance.
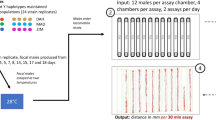
The experimental individuals used in the current study were the offspring of crosses of females from each of the RI lines to males from the two parental lines (Oregon (O) or 2b (B)). This crossing scheme is a modified North Carolina design III (Comstock and Robinson, 1948, 1952), in which the RI lines take the place of the F2 generation in the original North Carolina design III. We utilized this backcross design rather than directly screening the RI lines to avoid the effects of inbreeding depression on fitness-related traits, and to produce heterozygous genotypes to estimate degrees of dominance of QTL. Backcrossing each of the RI lines to both of the parental lines resulted in progeny with three genotypes (OO, OB, BB) at cold and heat tolerance QTL. The OO and OB genotypes occur in the progeny from the backcross to Oregon, whereas the BB and OB genotypes occur in the progeny from the backcross to 2b.
Thermotolerance phenotypes
All of the individuals screened were young flies (ie 5–7 days old), as previous studies have found that thermotolerance phenotypes can change as a function of age (Lamb and McDonald, 1973; David et al, 1998). All experimental individuals were crossed and maintained in vials containing standard cornmeal–agar–molasses medium sprinkled with live yeast to stimulate oviposition. Individuals were maintained at 25oC and 60% humidity until they were used in the experimental assay.
To measure cold tolerance in the parental lines, we utilized a modified version of the assay presented by David et al (1998). Specifically, 25 same-sex individuals were transferred without anesthesia into empty vials and placed in chambers containing melting ice (0°C±0.5). After 3 h, individuals were removed from the cold treatment and returned to room temperature (23°C±0.5), and the chill-coma recovery time was measured by recording the amount of time until an individual was able to stand on its legs. Two replicate vials were measured for each line and sex, yielding a cold tolerance profile for each parental line and sex (Figure 1a). To measure heat tolerance in the parental lines, we measured the percent survival after heat stress, (38°C±0.5). Two replicate vials for each parental line and sex were assayed. Each replicate contained 25 same-sex individuals that were transferred without anesthesia into empty vials and placed in a water bath at 38°C (±0.5) for either 10, 40, 70, 100, 130, or 160 min. After heat stress, these flies were immediately placed in vials containing 5 ml of standard cornmeal–agar–molasses medium and returned to 25°C and 60% humidity for 24 h. After 24 h, the percentage of surviving flies per vial was recorded for each sex, line, and exposure time, generating a heat tolerance profile for each parental line and sex (Figure 1b).
The heat and cold tolerance profiles (Figure 1) were then used to determine the average 50% heat survival or chill-coma recovery time points of the parental lines (Figure 1, dotted line), which was subsequently used as the more rapid assay point for the experimental (backcrossed) RI lines. The average 50% heat survival and chill-coma recovery time points for the parental lines were determined by regressing the percentage of flies that had either survived a heat exposure or recovered from chill coma on the time at 38°C (for heat survival) or at 23°C (for chill-coma recovery). Using this regression, the average 50% time point for both lines and sexes could be predicted for chill-coma recovery and heat-stress survival. The 50% heat survival was 110 min at 38°C (±0.5), whereas the average 50% chill-coma recovery time point for the parental lines was 11 min at room temperature (23°C±0.5). Thus, for the backcrossed RI lines, our assay for heat stress was the percentage of flies alive 24 h after a treatment of 110 min at 38°C. Our assay for cold stress was the percentage of flies recovered from chill coma (ie standing on their legs) at 11 min after being removed from the cold treatment (0°C for 3 h).
Quantitative genetic analyses
To determine if significant genetic variation existed for chill-coma recovery and heat-stress survival between the parental inbred lines (Oregon and 2b), we used a one- and two-way analysis of variance (ANOVA), which modeled the fixed effects of line (Oregon or 2b), sex, and their interaction (for two-way ANOVA) on chill-coma recovery or heat-stress survival at the 50% assay point. To understand the pattern of genetic variation present within the backcrossed RI lines, a set of analyses was performed on the chill-coma recovery and survival after heat stress. Data from the backcrossed RI lines were analyzed hierarchically in four different ANOVAs, so the main effects and their interactions could be examined in the full data as well as subsets. The first analysis was a random effects model, which identified genetic variance among lines (L) within each cross and sex based on the model y=μ+L+ɛ, where y is the thermotolerance phenotype of an RI line within sex and cross, μ is the overall mean, L is the random effect of line, and ɛ is the residual within line variance. Broad-sense heritabilities (H2) within each cross and sex were computed as  where σL2 is the among-line and σE2 the within-line variance component. In the analyses within sex and cross, we also estimated the genetic correlation between percent recovered from chill coma and percent survival after heat stress, where
where σL2 is the among-line and σE2 the within-line variance component. In the analyses within sex and cross, we also estimated the genetic correlation between percent recovered from chill coma and percent survival after heat stress, where  with covCH being the covariance among line means between the percent recovered from chill coma and survival after heat-stress scores within a cross and sex and σC and σH the square roots of the variance components for line term from the reduced-random-effects model within sex and cross for chill coma and heat stress. The next analysis was a mixed-model ANOVA separated by cross to identify genotype-by-sex interactions. In the mixed model separated by cross (Oregon or 2b), the variance in thermotolerance traits was partitioned by line, sex (male or female), and their interaction based on the model y=μ+L+S+L × S+ɛ, where y is the thermotolerance phenotype of an RI line within cross, μ is the overall mean, L is the random effect of line, S is the fixed effect of sex, and ɛ is the residual variance. The line-by-sex interaction term (L × S) from this analysis was used to evaluate the significance of the genotype-by-sex interaction (GSI) and to calculate the cross–sex genetic correlation
with covCH being the covariance among line means between the percent recovered from chill coma and survival after heat-stress scores within a cross and sex and σC and σH the square roots of the variance components for line term from the reduced-random-effects model within sex and cross for chill coma and heat stress. The next analysis was a mixed-model ANOVA separated by cross to identify genotype-by-sex interactions. In the mixed model separated by cross (Oregon or 2b), the variance in thermotolerance traits was partitioned by line, sex (male or female), and their interaction based on the model y=μ+L+S+L × S+ɛ, where y is the thermotolerance phenotype of an RI line within cross, μ is the overall mean, L is the random effect of line, S is the fixed effect of sex, and ɛ is the residual variance. The line-by-sex interaction term (L × S) from this analysis was used to evaluate the significance of the genotype-by-sex interaction (GSI) and to calculate the cross–sex genetic correlation  ) (Robertson, 1959), where covFM is the covariance among line means between males and females within a cross and σF and σM are the square roots of the variance components for line term from the reduced-random-effects model within sex and cross.
) (Robertson, 1959), where covFM is the covariance among line means between males and females within a cross and σF and σM are the square roots of the variance components for line term from the reduced-random-effects model within sex and cross.
To compare the amount of genetic variance within a cross and sex and trait, the coefficient of variance was estimated separately for each trait and sex within each cross. The coefficient of variance was calculated as  where x̄ is the average percent chill-coma recovery or the average survival after heat stress among lines and VL is the variance component among lines within crosses and sexes (Vieira et al, 2000). All analyses were carried out using SAS (SAS Institute, 1988) PROC GLM and VARCOMP procedures.
where x̄ is the average percent chill-coma recovery or the average survival after heat stress among lines and VL is the variance component among lines within crosses and sexes (Vieira et al, 2000). All analyses were carried out using SAS (SAS Institute, 1988) PROC GLM and VARCOMP procedures.
Molecular map
Cytological insertion sites of 81 polymorphic roo transposable elements were determined from each of the RI lines (Nuzhdin et al, 1997; Leips and Mackay, 2000; Vieira et al, 2000). Map positions of the markers were estimated from the observed recombination frequencies (r) between pairs of markers based on Kosambi's map function  , where d is the distance between markers in centimorgans (cM). Chromosome two was separated into two linkage groups as the recombination frequency between marker 50F and 57C was greater than 0.5 (Nuzhdin et al, 1997).
, where d is the distance between markers in centimorgans (cM). Chromosome two was separated into two linkage groups as the recombination frequency between marker 50F and 57C was greater than 0.5 (Nuzhdin et al, 1997).
QTL mapping
Composite interval mapping was used to identify QTL contributing to variation among lines in mean percent chill-coma recovery and mean percent survival after heat stress (Zeng, 1994) using Windows QTL Cartographer version 2.0 (Basten et al, 2003). Composite interval mapping tests the hypothesis that the interval between two adjacent markers contains a QTL influencing the trait, while simultaneously controlling for the effects of QTL outside the test interval. These analyses were conducted on line means for both traits and sexes. The conditioning markers were chosen for each analysis by forward selection/backward elimination stepwise regression. A conditioning window size of 10 cM was used such that only markers 10 cM away from the markers creating the test interval were included in the model. The likelihood-ratio (LR) test statistic is −2 ln(L0/L1), where L0/L1 is the ratio of the likelihood under the null hypothesis (L0, there is no QTL in the test interval) to the alternative hypothesis (L1, there is a QTL in the test interval). The LR test statistic at a genome location is distributed as χ2 with 2 d.f. under the null hypothesis and was evaluated every cM.
The significance level for each analysis to infer the presence of a QTL was determined by permutation. Empirical distributions of the LR test statistics under the null hypothesis of no association between test intervals and trait values were obtained for each analysis by randomly permuting the trait data 1000 times and calculating the maximum LR statistic across all intervals for each permutation. From this empirical distribution of the LR statistic under the null hypothesis, a significance threshold was calculated at an α=0.05 by determining the LR value demarcating the top 5% of the empirical distribution. Thus, the LR statistics calculated from the observed data that exceed this threshold indicates the presence of a QTL at this genomic location (Churchill and Doerge, 1994; Doerge and Churchill, 1996).
QTL effects
Estimates of the additive effects (a=(BB−OO)/2) and dominance deviations (d=OB−(OO+BB)/2) of each QTL for each sex and trait are provided by the QTL mapping analysis. The presence of pairwise epistatic effects was evaluated using a simple linear model of y=mx+my+mxmy+ɛ, where mx and my are the genotypes of markers x and y within a cross and sex.
Candidate genes
Positional candidate genes were identified by scanning the known and computationally predicted genes contained within each QTL region for loci with known effects on resistance to temperature stress or thermal and environmental sensation phenotypes, as represented in the FlyBase gene ontology database (FlyBase Consortium, 2003).
Results
Thermotolerance phenotypes in the parental lines
The chill-coma recovery and heat-stress survival profiles of the parental lines (Oregon and 2b; Figure 1) were used to determine the average 50% recovery or survival time points for each of the parental lines (Figure 1, dashed line). This average 50% time point was subsequently used in the high-throughput assay for the backcrossed RI lines. The average 50% chill-coma recovery time point for the parental lines was 11 min at room temperature (23°C±0.5), whereas the average 50% heat-survival time point was 110 min at 38°C±0.5.
The Oregon and 2b parental lines exhibited significantly different phenotypes for both the 50% chill-coma recovery (F1,4=48.21, P=0.0023; Figure 1) and heat-stress survival time (F1,4=236.26, P<0.0001; Figure 1). For chill-coma recovery, Oregon individuals had an average recovery, percentage of 81.17% (±4.2) in females and 82.95% (±4.0) in males, whereas 2b individuals had an average recovery percentage of 31.58% (±4.8) in females and 35.27% (±5.4) in males (Table 1). For survival after heat stress, Oregon individuals had an average survival of 30.0% (±6.0) in females and 16.0% (±5.2) in males, whereas 2b individuals had an average survival of 94.0% (±3.4) in females and 86.0% (±5.0) in males (Table 1). However, neither the main effect of sex nor the interaction of sex and line significantly influenced either of the thermotolerance phenotypes in the parental lines.
Quantitative genetic variation
Among lines: There was significant genetic variation in both thermotolerance phenotypes among the lines within both crosses and sexes (VL; Table 2; Figure 2); however, the magnitude of differences among lines was cross and sex specific (Table 2; Figure 2). Additionally, the broad-sense heritabilities (H2; Table 2) varied with each cross and sex. Estimates of H2 for chill-coma recovery ranged from 0.175 (RI × Oregon ♂) to 0.353 (RI × 2b ♂), whereas estimates of H2 for heat-stress survival ranged from 0.121 (RI × Oregon and RI × 2b ♀) to 0.323 (RI × Oregon ♂). This range of genetic variation is also reflected in the estimates of the coefficients of genetic variation (CVG; Table 2). The estimated genetic correlations (within sex and backcross) between percent recovery from chill coma and percent alive after heat stress (rG; Table 2) ranged from −0.036 (RI × Oregon ♂) to 0.189 (RI × Oregon ♀). However, none of these estimates was significantly different from zero, implying that unique genetic architectures exist in this population for chill-coma recovery and survival after heat stress (Table 2).

Distribution of phenotypes among the backcross RI lines for chill-coma recovery in females (a) and males (b) and for survival after heat stress in females (c) and males (d). Gray bars represent lines backcrossed to the 2b parental line, and the cross-hatched bars represent lines backcrossed to the Oregon parental line. Gray arrows represent the mean of the 2b parental line, and black arrows represent the mean of the Oregon parental line.
Genotype-by-sex interactions: Four separate mixed-model ANOVAs (separated by cross and trait) were used to test for interactions among the genotypes (lines) and sex. In these analyses (within each cross; Table 3), significant effects of line, sex, and line-by-sex interactions were identified that were cross and trait specific (Table 3). For chill-coma recovery, there were significant differences between the sexes in the 2b cross (in which females exhibit an increase of approximately 13% in the percentage of flies recovered from chill coma), but not in the Oregon cross. The main effect of line was significant within both the Oregon and 2b crosses (Table 3; Figure 2); however, the interaction of line and sex was significant in the cross to 2b but not in the cross to Oregon (Table 3). For survival after heat stress, there were highly significant differences between the sexes in both crosses. Females from both backcrossed populations exhibited an increase in survival of ∼42% on average. The main effect of line was significant within both the Oregon and 2b backcrossed flies (Table 3; Figure 2), whereas the interaction of line and sex was significant only within the 2b cross (Table 3). Additionally, the estimated cross–sex genetic correlation (rGS; Table 3) within each backcrossed population was 0.538 (Ore) and 0.576 (2b) for recovery from chill coma, whereas rGS was 0.145 (Ore) and 0.375 (2b) for survival after heat stress, suggesting overlapping but not identical genetic architectures between the sexes for each thermotolerance phenotype (Table 3; Figure 3).

Cross–sex correlations among line means for both backcrossed populations for percent recovered for chill coma (a) and percent survival after heat stress (b).
QTL mapping
Genome scan for thermotolerance QTL
We used composite interval mapping to localize the QTL influencing the variation in the percentage of flies recovered from chill coma and the percentage of flies surviving after heat stress among the backcrossed RI lines. Three QTL influencing chill-coma recovery were identified in females. Two of these QTL were on the second chromosome at cytological locations 34E–35B and 38E–46C, whereas the other one was on the third chromosome between cytological locations 76B and 87B (Table 4; Figure 4). In males, a single chill-coma QTL was detected on the second chromosome at the cytological location 35B–48D (Table 4; Figure 4). Two QTL affecting survival after heat stress were identified in females. The first was on the second chromosome between cytological locations 50B and 50F, whereas the second was at the left arm of the third chromosome at 63A–65A (Table 4; Figure 4). Three heat-stress QTL were identified in males: one on the second chromosome at 38A–48D and two on the third chromosome at cytological locations 61A–65A and 85E–88E (Table 4; Figure 4).

Genome scan for thermotolerance QTL. LR test statistic for chill coma (a) and survival after heat stress (b) plotted against chromosome position (cM) for the three major chromosomes. Dashed horizontal lines represent the significance threshold for each trait as determined by permutation, and triangles represent cytological marker locations.
Two QTL have significant overlap between the sexes for the same trait, whereas one QTL overlaps across the traits. The QTL common to both sexes within a trait are at the cytological location 63A–65A (QTL H2; Table 4; Figure 4) for survival after heat stress and at 34E–48D for recovery from chill coma (QTL C1/2; Table 4; Figure 4). The single QTL that overlaps across the traits is located at the cytological location 35B–48D and is detected in both sexes for recovery from chill coma (QTL C1/2; Table 4; Figure 4), but only in males for survival after heat stress (QTL H3: Table 4; Figure 4). These common QTL suggest overlapping but not identical genetic architectures influencing each thermotolerance phenotype within each sex.
QTL effects
We tested for epistatic effects between all pairwise combinations of markers used in our composite interval mapping, and did not detect any significant epistasis (after correction for multiple tests) among any pair of markers or QTL. We also estimated the additive effect (a) and dominance deviation (d) at the LR peak (Figure 4) for each identified QTL. The additive effect (a=(BB−OO)/2) estimates the magnitude and direction of the QTL effect, whereas the dominance deviation (d=OB−(OO+BB)/2) estimates the allelic effects on the Oregon and 2b alleles in the heterozygous (OB) genotypic class relative to the mean of the homozygous classes (BB and OO). Almost all of the QTL are either trait or sex specific. The estimated additive effects and percentage of the phenotypic variance explained were generally larger in males than females (Table 4). On average, each of the QTL identified in this genome screen explained approximately 10% of the phenotypic variance for a particular trait.
For percent recovery from chill coma, half of the identified QTL in both males and females exhibit additive effects that are consistent with the variation measured among the Oregon and 2b parental lines (Figure 1; Table 1). Two of four identified QTL decrease resistance to chill coma (QTL C1 (females), C1/2 (males)); Table 4) in 2b relative to Oregon, whereas two female-specific QTL (QTL C2 and C3; Table 4) act in the opposite direction, increasing the resistance to chill coma in 2b relative to Oregon. The chill-coma QTL in both sexes all exhibit some degree of partial dominance (Table 4). For percent survival after heat stress, most of the identified QTL in both males and females exhibit additive effects that are also consistent with the observed parental line divergence in heat-stress resistance (Table 1; Figure 1). Three of four QTL increase resistance to heat stress (QTL H2, H3, H4; Table 4) in 2b relative to Oregon, whereas one female-specific QTL (QTL H1; Table 4) exhibits a slight decrease in resistance to heat stress in 2b relative to Oregon. The heat-stress QTL exhibit a range of dominance deviations including essentially additively acting QTL (QTL H2 (in males) and H4; Table 4), to partially dominant QTL (QTL H1, H2 (in females), and H3; Table 4).
Discussion
Although thermotolerance phenotypes are known to be of great ecological and evolutionary significance in natural populations (Cohet et al, 1980; Krebs and Feder, 1997; Gilbert et al, 2001; Gilbert and Huey, 2001; Hoffmann et al, 2002, 2003; Ayrinhac et al, 2004; Kimura, 2004), substantially less is known about the genetic architecture underlying this important and complex quantitative genetic variation (Hoffmann et al, 2003). We have examined the genetic architecture of two ecologically important thermotolerance phenotypes, survival after heat stress and recovery from chill coma, in D. melanogaster. In this study, we have estimated the quantitative genetic parameters and localized QTL influencing variation in heat- and cold-stress resistance in a mapping population of RI lines, each of which was backcrossed to both of their parental lines (Oregon-R and 2b). We found significant genetic variation for both thermotolerance phenotypes among lines in our mapping population and we were able to detect seven QTL. Each of these QTL mapped to regions on the second and third chromosomes, and they exhibit relatively little colocalization between the two traits or sexes, suggesting that overlapping but generally unique genetic architectures underlie resistance to high- and low-temperature extremes within each sex. The detected QTL accounted for a modest portion (14–28%) of the total quantitative genetic variance among lines for each of these phenotypes, and the estimated additive effects of the QTL are consistent with direction of the divergence between the parental line phenotypes. That is, the cold tolerance QTL generally tend to decrease resistance to chill coma in 2b relative to Oregon, whereas the heat-stress QTL tend to increase resistance to heat stress in 2b relative to Oregon. Finally, all of the identified QTL exhibit either additive or partially dominant allelic effects.
One other study has used QTL analyses to identify genomic regions influencing heat-stress resistance phenotypes (Norry et al, 2004), and no previous work has attempted to localize QTL influencing cold-stress resistance in D. melanogaster. The data presented in this study confirm and complement previous QTL and candidate gene studies investigating the genetic control of temperature-stress resistance phenotypes (reviewed by Hoffmann et al, 2003). We detected QTL that encompass reasonable candidate loci that also exhibit colocalization with other heat-stress QTL (Norry et al, 2004), and we also localized new QTL to genomic regions where thermotolerance candidate loci have not been extensively investigated. Two of the four QTL identified in the study of Norry et al (2004) colocalize with our survival-after-heat-stress QTL: our QTL H3 (38E–48D; in males only) and QTL H4 (85E–88E; in males only). Additionally, a third QTL from Norry et al (2004) localized in a region of poor marker coverage in their study (66E–67D10) was very close to QTL H2 (61A–65A; in both sexes) detected in this study. This colocalization of QTL between two studies with mapping populations that have very different genetic histories implies that the QTL common to both studies represent genomic regions with a general role in heat-stress resistance.
One striking result from our data is the overall lack of colocalization between cold and heat tolerance QTL. Although the high- and low-temperature extremes are simply components of a spectrum of possible temperatures that an individual may encounter during its lifespan, our results suggest that the underlying genetic mechanisms for dealing with diverse temperature extremes are generally unique. In this study, only a single QTL exhibits colocalization for both cold- and heat-stress resistance. This QTL (cold QTL C1/2 and heat QTL H3; Table 4; Figure 4) was localized to the middle of chromosome 2. In males, this QTL has a large effect (Table 4; Figure 4), accounting for 14 and 12% of the genetic variance in cold- and heat-stress resistance, respectively. However, the additive effects of this QTL are antagonistic between cold and heat resistance; that is, homozygous 2b individuals exhibit a decreased resistance to cold and an increased resistance to heat, whereas the homozygous Oregon individuals exhibit the opposite pattern. Additionally, this region was also identified in an independent mapping population and was shown to influence high-temperature knockdown resistance (Norry et al, 2004). Thus, this region is a general thermotolerance QTL that exhibits a tradeoff in thermal-stress resistance in one sex. Although we do detect colocalization of both heat and cold QTL in this region, the actual genetic causal effect of this colocalization (ie close linkage or pleiotropy) can only be determined by further high-resolution mapping. Thus, with our current data, it is impossible to determine if this general thermotolerance QTL represents a single or multiple linked loci influencing heat and cold tolerance. Similar genetic tradeoffs between heat- and cold-stress resistance have been identified on the right arm of chromosome 3 at the locus Hsr-omega (Anderson et al, 2003). However, given the general lack of colocalization between heat and cold resistance QTL in this study and the small number of studies that have detected and/or localized tradeoffs in thermotolerance phenotypes (Hoffmann et al, 2002; Anderson et al, 2003; Kimura, 2004), it seems that individual loci affecting tradeoffs between heat- and cold-stress resistance may be the exception rather than the rule for thermotolerance loci. Future evaluation of the presence or absence of general thermotolerance QTL and their effects under diverse temperature stresses will ultimately rely on the dissection of the QTL and the identification of the individual thermotolerance candidate genes (Mackay, 2001).
In this study, we have not genetically dissected each of our QTL to the level of the individual gene using deficiency complementation or high-resolution recombination mapping (Pasyukova et al, 2000), but we have identified several candidate genes based on their mutant phenotypic effects on various components of thermotolerance and thermal sensation (FlyBase Consortium, 2003). For example, Trap1 (42C) maps within the general thermotolerance QTL (QTL C1/2 and H3). Trap1 is a heat-shock protein-related gene that has similar properties to Hsp90 (Felts et al, 2000; Pflanz and Hoch, 2000), and has thus been proposed to be involved in general defense response and response to various stressors (FlyBase Consortium, 2003). Our colocalization of both heat and cold QTL to this region of chromosome 2 further supports these roles for Trap1. Furthermore, screens of co-isogenic single P-element insertion lines (Bellen et al, 2004) for heat- and cold-stress resistance have implicated Trap1 in the response to thermal stress (TJ Morgan, ED Ozoy, LH Duncan, and TFC Mackay, unpublished data).
In addition to the regions of complete overlap between cold QTL C1/2 and heat QTL H3, there are also unique candidate genes within each QTL. For example, pickpocket (ppk; 35B) maps within the cold QTL C1 in females and QTL C1/2 in males. ppk encodes a gene product that directly influences sodium channel activity (Adams et al, 1998), and has also been implicated in larval thermal and mechano-sensation (Adams et al, 1998; Kim et al, 2002b). Another candidate gene involved in mechano-sensation, no-mechanoreceptor potential A (nompA; 47F), also maps within the cold QTL C1/2 in males. nompA encodes a large transmembrane protein thought to be involved in the formation of mechanical linkage required to transmit mechanical stimuli (eg tactile or auditory environmental stimuli; Chung et al, 2001). Finally, two genes involved in the catecholamine biosynthesis pathway, Catecholamines up (Catsup; 37B) and Dopa decarboxylase (Ddc; 37C), are contained within the cold QTL C1/2 in males. The catecholamine pathway is responsible for neurotransmitter production and melanization and sclerotization of the cuticle (Wright, 1987), and has also been implicated in the response to various stressors including temperature stress (Baden et al, 1996; Sabban and Kvetnansky, 2001). Furthermore, genes in this pathway have been implicated as positional candidate genes affecting variation in lifespan (De Luca et al, 2003).
Sex-specific QTL effects appear to be a common feature of the genetic architecture of complex traits. QTL with sex-specific effects have been found in Drosophila for sensory bristle number (Dilda and Mackay, 2002), lifespan (Nuzhdin et al, 1997; Leips and Mackay, 2000; Vieira et al, 2000), and olfactory behavior (Fanara et al, 2002). In this study, five of the seven QTL detected were sex specific. The two QTL common to both sexes were the cold QTL C1/2 (34E–48D) discussed above and the heat QTL H2 (61A–65A). The heat QTL H2 embraces a genomic region containing numerous positional candidate genes involved in stress resistance and response to heat. These include methuselah (mth; 61C), which has been shown to extend lifespan and increase stress resistance in mutant lines (Lin et al, 1998), and Hsp83 (63B) and DnaJ-1 (64E), both of which are upregulated in response to heat stress (Leemans et al, 2000). Thus, the tip of the left arm of chromosome 3 represents a genomic region with important effects on general heat-stress resistance.
We also localized three QTL that were identified for a single trait and sex combination. These include the cold QTL C3 (76B–87B) and the heat QTL H1 (50B–50F) in females, and the heat QTL H4 (85E–88E) in males. Three genes that have previously been implicated in various responses to cold map within cold QTL C3 (79B–87B): Frost (Fst; 85E), Sialic acid phosphate synthase (Sas; 87B), and desat2 (87B). Fst is upregulated during cold stress (Goto, 2001). Sas has been implicated in ice binding and response to freezing (Kim et al, 2002a). desat2 is involved in cuticular hydrocarbon biosynthesis (Coyne et al, 1999), and Cosmopolitan and Zimbabwe alleles of this locus vary in their cold tolerance (Greenberg et al, 2003). Heat shock protein cognate 5 (Hsc70-5; 50E) colocalizes with the heat QTL H1 (50B–50F) in females and four heat-stress genes colocalize with heat QTL H4 (85E–88E) in males (Hsp70 (87A), heat shock protein cognate 2 (87D), heat shock protein cognate 4 (88E), and the lifespan gene foxo (88A; Giannakou et al, 2004)).
In the current study, we have conducted the first genome scan for QTL influencing the ecologically important phenotypes of heat- and cold-stress resistance in D. melanogaster. Our results have localized three cold-stress QTL and four heat-stress QTL to the second and third chromosomes. Most of the identified QTL exhibited either trait- or sex-specific effects, suggesting that although resistances to cold and heat stress represent physiological responses to extremes of a continuous thermal spectrum, the genetic architectures underlying each of the phenotypes are generally unique. Although we identified seven QTL in this study, we must acknowledge that QTL identified from this mapping population almost certainly represent only a subset of all of the thermotolerance loci. Furthermore, the RI lines utilized in this current study were chosen for practical reasons; the genotypes of the molecular markers had previously been determined in these lines and other life-history QTL have previously been mapped in this population. Thus, these QTL may not represent the genomic regions influencing potential adaptive evolution in nature. However, this study is the first step in beginning the process of genetically dissecting two ecologically important phenotypes in D. melanogaster. Future analyses will build on this genome scan through the use of genome-wide screens of single P-element insertion lines, genome-wide expression profiling, and further QTL analyses of populations with highly divergent naturally derived thermotolerance phenotypes.
References
Adams CM, Anderson MG, Motto DG, Price MP, Johnson WA, Welsh MJ (1998). Ripped pocket and pickpocket, novel Drosophila DEG/ENaC subunits expressed in early development and in mechanosensory neurons. J Cell Biol 140: 143–152.
Anderson AA, Collinge JE, Hoffman AA, Kellett M, McKechnie SW (2003). Thermal tolerance trade-offs associated with the right arm of chromosome 3 and marker by the Hsr-omega gene in Drosophila melanogaster. Heredity 90: 195–202.
Anderson AA, Hoffman AA, McKechnie SW (2005). Response to selection for rapid chill-coma recovery in Drosophila melanogaster: physiology and life-history traits. Genet Res Camb 85: 15–22.
Ayrinhac A, Debat V, Gilbert P, Kister AG, Legout H, Moreteau B et al (2004). Cold adaptation in geographical populations of Drosophila melanogaster: phenotypic plasticity is more important than genetic variability. Funct Ecol 18: 700–706.
Baden HP, Kollias N, Anderson RR, Hopkins T, Raftery L (1996). Drosophila melanogaster larvae detect low doses of UVC radiation as manifested by a writhing response. Arch Insect Biochem Phys 32: 187–196.
Basten CJ, Weir BS, Zeng ZB (2003). QTL Cartographer: A Reference Manual and Tutorial for QTL Mapping Program in Statistical Genetics. Department of Statistics, Bioinformatics Research Center, North Carolina State University: Raleigh, NC.
Bellen HJ, Levis RW, Liao G, He Y, Carlson JW, Tsang G et al (2004). The BDGP gene disruption project: single transposon insertions associated with 40% of Drosophila genes. Genetics 167: 761–781.
Bettencourt BR, Kim IY, Hoffman AA, Feder ME (2002). Response to natural and laboratory selection at the Drosophila Hsp70 genes. Evolution 56: 1796–1801.
Bubli O, Imasheva AG, Loeschcke V (1998). Selection for knock-down resistance to heat in Drosophila melanogaster at high and low larval densities. Evolution 56: 619–625.
Cavicchi S, Guerra D, La Torre V, Huey RB (1995). Chromosomal analysis of heat-shock tolerance in Drosophila melanogaster evolving at different temperatures in the laboratory. Evolution 49: 676–684.
Chung YD, Zhu J, Han Y, Kernan MJ (2001). nompA encodes a PNS-specific, ZP domain protein required to connect mechanosensory dendrites to sensory structures. Neuron 29: 415–428.
Churchill GA, Doerge RW (1994). Empirical threshold values for quantitative trait mapping. Genetics 13: 963–971.
Clarke A (1996). The influence of climate chance of the distribution and evolution of organisms. In: Johnston IA, Bennett AF (eds) Animals and Temperature: Phenotypic and Evolutionary Adaptation. Cambridge University Press: Cambridge. pp 377–407.
Cohet Y, Vouidibio J, David JR (1980). Thermal tolerance and geographic distribution: a comparison of cosmopolitan and tropical endemic Drosophila species. J Therm Biol 5: 69–74.
Comstock RE, Robinson HF (1948). The components of genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4: 254–266.
Comstock RE, Robinson HF (1952). Estimation of average dominance of genes. In: Gowen JW (ed) Heterosis. Iowa State College Press: Ames, IA. pp 494–516.
Coussins AR, Bowler K (1987). Temperature Biology of Animals. Chapman & Hall: New York.
Coyne JA, Wicker-Thomas C, Jallon JM (1999). A gene responsible for a cuticular hydrocarbon polymorphism in Drosophila melanogaster. Gen Res Camb 73: 189–203.
Dahlgaard J, Loeschcke V, Michalak P, Justesen J (1998). Induced thermotolerance and associated expression of the heat-shock protein Hsp70 in adult Drosophila melanogaster. Funct Ecol 12: 786–793.
David JR, Capy P (1988). Genetic variation of Drosophila melanogaster natural populations. Trends Genet 4: 106–111.
David JR, Gilbert P, Moreteau B, Gilchrist GM, Huey RB (2003). The fly that came in from the cold: geographic variation of recovery time from low-temperature exposure in Drosophila subobscura. Funct Ecol 17: 425–430.
David JR, Gilbert P, Pla E, Petacy G, Karan D, Moreteau B (1998). Cold stress tolerance in Drosophila: analysis of chill coma recovery in D. melanogaster. J Therm Biol 23: 291–299.
De Luca M, Roshina NV, Geiger-Thornsberry GL, Lyman RF, Pasyukova EG, Mackay TFC (2003). Dopa decarboxylase (Ddc) affects variation in Drosophila longevity. Nat Genet 34: 429–433.
Dilda CL, Mackay TFC (2002). The genetic architecture of Drosophila sensory bristle number. Genetics 162: 1655–1674.
Doerge RW, Churchill GA (1996). Permutation tests for multiple loci affecting a quantitative character. Genetics 142: 285–294.
Fanara JJ, Robinson KO, Rollmann S, Anholt RRH, Mackay TFC (2002). Vanaso is a quantitative trait locus for Drosophila olfactory behavior. Genetics 162: 1321–1328.
Feder ME, Krebs RA (1998). Natural and genetic engineering of the heat shock protein Hsp70 in Drosophila melanogaster: consequences for thermotolerance. Am Zool 38: 503–517.
Felts SJ, Owen BAL, Nguyen PM, Trepel J, Donner DB, Toft DO (2000). The hsp90-related protein TRAP1 is a mitochondrial protein with distinct functional properties. J Biol Chem 275: 3305–3312.
FlyBase Consortium (2003). The FlyBase database of the Drosophila genome projects and community literature. Nucleic Acids Res 31: 172–175. http://flybase.org.
Giannakou ME, Goss M, Junger MA, Hafen E, Leevers SJ, Partridge L (2004). Long-lived Drosophila with overexpressed dFOXO in adult fat body. Science 305: 361.
Gibbs AG, Perkins MC, Markow TA (2003). No place to hide: microclimates of sonoran desert Drosophila. J Therm Biol 28: 353–362.
Gilbert P, Huey RB (2001). Chill-coma temperature in Drosophila: effects of developmental temperature, latitude and phylogeny. Phys Biochem Zool 74: 429–434.
Gilbert P, Moreteau B, Pétavy G, Karan D, David JR (2001). Chill-coma tolerance, a major climatic adaptation among Drosophila species. Evolution 55: 1063–1068.
Gilchrist GW, Huey RB (1999). The direct response of Drosophila melanogaster to selection on knockdown temperature. Heredity 83: 15–29.
Gilchrist GW, Huey RB, Partridge L (1997). Thermal sensitivity of Drosophila melanogaster: evolutionary responses of adults and eggs to laboratory natural selection at different temperatures. Phys Zool 70: 403–414.
Goto SG (2000). Expression of Drosophila homologue of senescence marker protein-30 during cold acclimation. J Insect Physiol 46: 1111–1120.
Goto SG (2001). A novel gene that is up-regulated during recovery from cold shock in Drosophila melanogaster. Gene 270: 259–264.
Greenberg AJ, Moran JR, Coyne JA, Wu CI (2003). Ecological adaptation during incipient speciation revealed by precise gene replacement. Science 302: 1754–1757.
Hoffmann AA, Anderson AA, Hallas R (2002). Opposing clines for high and low temperature resistance in Drosophila melanogaster. Ecol Lett 5: 614–618.
Hoffmann AA, Parsons PA (1991). Evolutionary Genetics of Environmental Stress. Oxford University Press: Oxford.
Hoffmann AA, Sorensen JG, Loeschcke V (2003). Adaptation of Drosophila to temperature extremes: bringing together quantitative and molecular approaches. J Thermal Biol 28: 175–216.
Kaidanov LZ (1990). The rules of genetical alterations in Drosophila melanogaster inbred lines determined by selection. Arh Biol Nauka 42: 131–148.
Kim K, Lawrence SM, Park J, Pitts L, Vann WF, Betenbaugh MJ et al (2002a). Expression of a functional Drosophila melanogaster N-acetylneuraminic acid (Neu5Ac) phosphate synthase gene: evidence for endogenous sialic acid biosynthetic ability in insects. Glycobiology 12: 73–83.
Kim MJ, Zinkevich N, Israel P, Pettus J, Ainsley J, Bosenko D et al (2002b). Molecular mechanisms of thermosensory signal transduction. Drosophila Research Conference, San Diego, CA, Vol 43, p 784A.
Kimura MT (2004). Cold and heat tolerance of drosophilid flies with reference to their latitudinal distributions. Oecologia 140: 442–449.
Krebs RA, Feder ME (1997). Natural variation in the expression of the heat-shock protein Hsp70 in a population of Drosophila melanogaster and its correlation with tolerance of ecologically relevant thermal stress. Evolution 51: 173–179.
Lachaise D, Cariou M, David JR, Lemeunier F, Tsacas L, Ashburner M (1988). Historical biogeography of the Drosophila melanogaster species subgroup. Evol Biol 22: 159–225.
Lamb MJ, McDonald RP (1973). Heat tolerance changes with age in normal and irradiated Drosophila melanogaster. Exp Gerontol 8: 207–217.
Leather S, Walter K, Bale J (1993). The Ecology of Insects Overwintering. Cambridge University Press: Cambridge.
Leemans R, Egger B, Loop T, Kammermeier L, He H, Hartmann B et al (2000). Quantitative transcript imaging in normal and heat-shocked Drosophila embryos by using high-density oligonucleotide arrays. Proc Natl Acad Sci USA 97: 12138–12143.
Leips J, Mackay TF (2000). Quantitative trait loci for life span in Drosophila melanogaster: interactions with genetic background and larval density. Genetics 155: 1773–1788.
Lerman DN, Feder ME (2001). Laboratory selection at different temperature modifies heat-shock transcription factor (HSF) activation in Drosophila melanogaster. J Exp Biol 204: 315–323.
Lin YJ, Seroude L, Benzer B (1998). Extended life-span and stress resistance in the Drosophila mutant methuselah. Science 282: 943–946.
Lindsley DL, Zimm GG (1992). The Genome of Drosophila melanogaster. Academic Press: San Diego.
Loeschcke V, Krebs RA (1996). Selection for heat-shock resistance in larval and in adult Drosophila buzzatii: comparing direct and indirect responses. Evolution 50: 2354–2359.
Mackay TFC (2001). Quantitative trait loci in Drosophila. Nat Rev Genet 2: 11–20.
McColl G, McKechnie SW (1999). The Drosophila heat shock Hsr-omega gene: an allele frequency cline detected by quantitative PCR. Mol Biol Evol 16: 1568–1574.
McColl G, Hoffmann AA, McKechnie SW (1996). Response of two heat shock genes to selection for knockdown heat resistance in Drosophila melanogaster. Genetics 143: 1615–1627.
Morrison WW, Milkman R (1978). Modification of heat resistance in Drosophila by selection. Nature 273: 49–50.
Norry FM, Dahlgaard J, Loeschcke V (2004). Quantitative trait loci affecting knockdown resistance to high temperature in Drosophila melanogaster. Mol Ecol 13: 3585–3594.
Nuzhdin SV, Pasyukova EG, Dilda CL, Zeng ZB, Mackay TFC (1997). Sex-specific quantitative trait loci affecting longevity in Drosophila melanogaster. Proc Natl Acad Sci USA 94: 9734–9739.
Pasyukova EG, Nuzhdin SV (1993). Doc and copia instability in an isogenic Drosophila melanogaster stock. Mol Gen Genet 240: 302–306.
Pasyukova EG, Viera C, Mackay TFC (2000). Deficiency mapping of quantitative trait loci affecting longevity in Drosophila melanogaster. Genetics 156: 1129–1146.
Pflanz R, Hoch M (2000). dtrap-1 encodes a novel member of the heat shock super family of proteins and is expressed in derivatives of all three germ layers during Drosophila embryogenesis. Mech Dev 96: 219–222.
Robertson A (1959). The sampling variance of the genetic correlation coefficient. Biometrics 15: 469–485.
Rohmer C, David JR, Moreteau B, Joly D (2004). Heat induced male sterility in Drosophila melanogaster: adaptive genetic variations among geographic populations and role of the Y chromosome. J Exp Biol 207: 2735–2743.
Sabban EL, Kvetnansky R (2001). Stress-triggered activation of gene expression in catecholaminergic systems: dynamics of transcriptional events. Trends Neurosci 24: 91–98.
SAS Institute (1988). SAS/STAT User's Guide, Release 6.12 Edition. SAS Institute: Cary, NC.
Tucic N (1979). Genetic capacity for adaptation to cold resistance at different developmental stages of Drosophila melanogaster. Evolution 33: 350–358.
Vieira C, Pasyukova EG, Zeng ZB, Hackett JB, Lyman RF, Mackay TFC (2000). Genotype–environment interaction for quantitative trait loci affecting life span in Drosophila melanogaster. Genetics 154: 213–227.
Wright TRF (1987). The genetics of biogenic amine metabolism, sclerotization, and melanization, in Drosophila melanogaster. Adv Genet 24: 127–222.
Zeng ZB (1994). Precision mapping of quantitative trait loci. Genetics 136: 1457–1468.
Acknowledgements
We thank LH Duncan, SL Heinsohn, and RH Wilson for assistance with flies and EG Pasyukova and SV Nuzhdin for creating the RI lines used in this study. We also thank two anonymous reviewers for helpful comments on this manuscript. This work was supported by National Institutes of Health Grants F32-GM0666 to TJM and GM-45344 and GM-45146 to TFCM.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Morgan, T., Mackay, T. Quantitative trait loci for thermotolerance phenotypes in Drosophila melanogaster. Heredity 96, 232–242 (2006). https://doi.org/10.1038/sj.hdy.6800786
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.hdy.6800786
Keywords
This article is cited by
-
Pleiotropic fitness effects of the lncRNA Uhg4 in Drosophila melanogaster
BMC Genomics (2022)
-
False and true positives in arthropod thermal adaptation candidate gene lists
Genetica (2021)
-
Adaptation of Drosophila subobscura chromosomal inversions to climatic variables: the Balkan natural population of Avala
Genetica (2021)
-
Measuring the evolutionary potential of a winter-active parasitic wasp to climate change
Oecologia (2020)
-
Stage-specific genotype-by-environment interactions for cold and heat hardiness in Drosophila melanogaster
Heredity (2019)
