
Abstract
A proposed mechanism in the pathogenesis of biliary atresia involves an initial virus-induced, progressive T cell–mediated inflammatory obliteration of bile ducts. The aim of this study was to characterize the inflammatory environment present within the liver of infants with biliary atresia to gain insight into the role of a primary immune-mediated process versus a nonspecific secondary response to biliary obstruction. Frozen liver tissue obtained from patients with biliary atresia, neonatal giant cell hepatitis, total parenteral nutrition (TPN)–related cholestasis, choledochal cysts, and normal control subjects was used for fluorescent immunohistochemistry studies of cellular infiltrates, cytokine mRNA expression, and in situ hybridization for localization of cytokine-producing cells. Immunohistochemistry revealed increases in CD8+ and CD4+ T cells and Kupffer cells (CD68+) in the portal tracts of biliary atresia. Reverse transcription–PCR analysis of biliary atresia tissue showed a Th1-type cytokine profile with expression of IL-2, interferon-γ, tumor necrosis factor-α, and IL-12. This profile was not seen in normal, neonatal hepatitis or choledochal cyst livers but was present in TPN-related cholestasis. In situ hybridization revealed that the Th1 cytokine–producing cells were located in the portal tracts in biliary atresia and in the parenchyma of TPN-related cholestasis. A distinctive portal tract inflammatory environment is present in biliary atresia, involving CD4+ Th1 cell–mediated immunity. The absence of similar inflammation in other pediatric cholestatic conditions suggests that the portal tract inflammation in biliary atresia is not a secondary response to cholestasis but rather indicates a specific immune response involved in the pathogenesis of biliary atresia.
Similar content being viewed by others
Main
Biliary atresia is a progressive, inflammatory cholangiopathy of infancy that culminates in fibrosis and obliteration of both the extrahepatic and intrahepatic bile ducts. Two forms of biliary atresia have been proposed: an embryonic/fetal form in which there seems to be defective morphogenesis of the extrahepatic bile duct and a perinatal/acquired form in which a normally developed bile duct seems to undergo pre- or postnatal injury and fibro-obliteration (1). The most effective treatment for biliary atresia is the surgical construction of a portoenterostomy if performed before 2 to 3 mo of age, followed by orthotopic liver transplantation if progressive liver disease develops despite surgery. Currently, ∼70–80% of biliary atresia patients will eventually require liver transplantation, attesting to the need to better understand the underlying cause and pathogenesis of this disease.
The cause of biliary atresia remains unknown. Current theories of pathogenesis have included but are not limited to genetic defects in ductal development (embryonic/fetal form) (2), occult viral infection, and immune-mediated ductal destruction (perinatal/acquired form) (3–5). Many investigators have sought to identify a viral cause of biliary atresia. Evidence for reovirus type 3 infection in biliary atresia was demonstrated in some studies (6–8) but not substantiated by others (9,10). Likewise, group C rotavirus was found in patients with biliary atresia by Riepenhoff-Talty et al. (11); however, another study failed to identify this virus (12). Conflicting studies have also been reported for cytomegalovirus (13,14).
Histologic evidence at the time of diagnosis of biliary atresia supports the primary role of the immune system in mediating injury to the bile duct epithelium. At diagnosis, the extrahepatic bile duct is partially or entirely obliterated with residual inflammatory cells present within the remnant, whereas the intrahepatic ductal lesion consists of an ongoing inflammatory response composed of periductal lymphocytes invading the ducts (15,16). Previous studies have found that the inflammatory milieu present in the portal tracts consists of Kupffer cells, (17–19) CD4+ T cells (20,21), and natural killer (NK) cells (21). However, controversy persists as to the cause of this portal tract inflammation, and it remains unclear whether biliary atresia is a primary immune-mediated disease or the inflammation is a secondary response to bile stasis and biliary obstruction.
The immune response to a viral infection or an autoimmune response is generally mediated by a T helper cell (CD4+) type 1 (Th1) inflammatory process. When naïve CD4+ T cells encounter antigen, they have the potential to differentiate into either Th1 cells (cell-mediated immunity) or Th2 cells (humoral-mediated immunity). Cytokines characteristic of a Th1 response include IL-2, interferon (IFN)-γ, tumor necrosis factor (TNF)-α, and IL-12 (from macrophages), whereas Th2 cytokines include IL-4 and IL-5. TNF-α is associated with Th1 responses and can be produced by both Th1 cells and activated macrophages that have been stimulated by the Th1-derived cytokine IFN-γ (22). Although previous reports have demonstrated the presence of CD4+ and CD8+ T cells in portal tracts of biliary atresia patients (21,23), characterization of the cytokine milieu, important in understanding the immune pathogenesis of disease, has not been performed. The aim of this study was to characterize the inflammatory environment present within the liver of infants with biliary atresia to gain insight into the potential role of a primary immune-mediated process in the pathogenesis of this disease.
METHODS
Study subjects and liver tissue retrieval.
Between January 1998 and February 2003, liver tissue was collected from three groups of patients:1) biliary atresia (perinatal form) at the time of Kasai portoenterostomy (n = 10); 2) other cholestatic diseases, including a) infants who had a history of chronic total parenteral nutrition (TPN) administration and underwent an intraoperative cholangiogram for persistent cholestasis (n = 4), b) infants who had severe cholestasis requiring intraoperative cholangiogram to rule out biliary atresia and were found to have neonatal giant cell hepatitis (NH; one idiopathic, two Nieman-Pick disease; n = 3), and c) an infant and children with choledochal cysts (CDCs) at the time of cyst excision (n = 4); and 3) children with normal livers at the time of a mesenterico-left portal vein bypass procedure (n = 4). None of the biliary atresia patients had any associated nonhepatic congenital anomalies. Infants with TPN-related cholestasis, NH, and CDCs were chosen as appropriate cholestatic disease control subjects for comparison to biliary atresia because of clinical and histologic similarities. The clinical presentations of these four disorders share many features, including obstructive jaundice, pale or acholic stools, and laboratory indices of cholestasis. Histologic features common to all four disorders include bile duct proliferation, portal tract bile duct plugging and inflammation, and variable degrees of portal tract fibrosis (24). Liver tissue obtained from children who underwent surgery (mesenterico-left portal vein bypass) for treatment of portal hypertension represented the normal liver control group.
Liver tissue for this study was obtained in the following manner. In the operating room, wedge liver biopsies were divided and a portion was placed immediately in OCT freezing media or wrapped in aluminum foil, snap-frozen in liquid nitrogen, and stored at −70°C until analysis. All tissue specimens underwent RNA extraction, and immunohistochemistry studies were performed on specimens that had adequate amount of tissue. Tissue was also processed for light microscopy for clinical evaluation. This study was approved by the Institutional Review Board at Children's Memorial Hospital and the Colorado Multi-Institutional Review Board. Informed consent was obtained from parents.
Fluorescent immunohistochemistry.
Frozen liver tissue was sectioned (7-μm thickness) and placed on electrostatic microscope slides, dried at room temperature, and fixed in 2% paraformaldehyde/PBS. Endogenous tissue peroxidases were abolished with 6% hydrogen peroxide. Nonspecific binding of antibody to the Fc receptor was blocked with purified antibody to CD32 (FcR) or 20% normal human serum. All MAb to human antigens were purchased from BD PharMingen (San Jose, CA, U.S.A.), and optimal antibody concentrations (low background, high signal) were determined for each antibody tested. Primary antibodies were biotinylated anti-CD3 (IgG1; clone UCHT1; 1 μg/mL), FITC anti-CD4 (IgG1; clone RPA-T4; 2 μg/mL), biotinylated anti-CD8 (IgG1; clone RPA-T8; 1 μg/mL), anti- CD20 (IgG2b; clone2H7; 5 μg/mL), anti-CD68 (IgG2b; clone Y1/82A; 5 μg/mL), and anti-NK1 (IgM; DAKO-clone NK1; 5 μg/mL). Biotinylated mouse isotype controls included IgG1 (MOPC-31C), IgG2b (MPC-11), and IgM (G155–228). Isotype controls were tested at the various concentrations noted above. MAb to cytokeratin 7 (clone OV-TL 12/30; 2 μg/mL) was purchased from DAKO labs (Glostrup, Denmark). MAb were diluted in TNB blocking buffer and incubated with serial sections of liver for 60 min, washed in PBS × 3, followed by 30-min incubation with the secondary antibody conjugated to horseradish peroxidase (HRP). Secondary antibodies were purchased from BD PharMingen and consisted of biotinylated anti-mouse IgM (rat IgG2b; II/41), streptavidin-HRP, goat HRP–conjugated anti-mouse IgG, or HRP-conjugated anti-FITC. After extensive washing, the tyramide system amplification substrate (NEN Life Sciences, Boston, MA, U.S.A.) that reacts with HRP and is conjugated to a fluorochrome (fluorescein-green or cyanine 3-red) was applied for 5 min. The tissue was then counterstained with the nuclear stain DAPI and sealed with VectaShield (Vector Laboratories, Burlingame, CA, U.S.A.) to maintain integrity of the tissue as well as the fluorescence. Cellular staining was visualized under ×100 and ×200 magnification with the Zeiss Axioplan2 microscope (Thornwood, NY, U.S.A.). Portal tracts were identified by characteristic vessel formation and bile duct epithelial nuclear staining and verified by serial sections with cytokeratin 7 staining, which is specific for bile duct epithelium. Furthermore, each specimen had a serial section with an isotype antibody control. The portal tracts were easily identified within this section because the surrounding hepatocytes display a faint nonspecific background staining, whereas the portal tracts do not. Digital photographs were obtained with autocalibration of exposure time through the Slidebook analysis program (Denver, CO, U.S.A.). Photographs were taken of every portal tract (2–7 tracts per specimen) and selected hepatic lobule areas (two areas per specimen) for each antibody experiment from each specimen. Analysis of each tissue specimen was repeated twice.
Quantification and statistical analysis of portal tract cellular infiltrates.
All portal tracts were photographed at ×200 magnification. The investigator who quantified the number of positive fluorescent cells per portal tract was blinded to the diagnosis of the patient. The mean total number of cells per portal tract per patient was calculated from portal tracts examined. The mean portal cell count from each patient was totaled as a group (normal, other cholestatic disease, biliary atresia), and the mean and SD were determined. Comparison of biliary atresia versus other cholestatic diseases and biliary atresia versus normal liver was analyzed statistically with the ANOVA and Tukey-Kramer Multiple Comparisons Test (InStat program, San Diego, CA, U.S.A.). Differences in means were considered significant at p < 0.05.
RNA extraction and reverse transcription–PCR.
Snap-frozen tissue was lysed with Trizol (Invitrogen Life Technologies, Carlsbad, CA, U.S.A.), and RNA was extracted with chloroform, washed with ethanol, and resuspended in DEPC-treated water. Quantification of RNA was performed with a spectrophotometer, and the purity of RNA was verified with a 260/280 ratio between 1.8 and 2.2. The entire RNA sample was then treated with deoxyribonuclease 1 (Sigma Chemical Co., St. Louis, MO, U.S.A.) to remove any contaminating genomic DNA. Verification of lack of DNA contamination was performed by elimination of reverse transcriptase during cDNA synthesis in control samples. cDNA synthesis was performed with 2 μg of RNA using the Advantage reverse transcription (RT) for PCR kit from Clontech (Palo Alto, CA, U.S.A.). RT-PCR was performed using 2 μl of cDNA, HotStarTaq-polymerase (Qiagen, Valencia, CA, U.S.A.), 10×Taq buffer,10× dNTP mix, and cytokine-specific primer pairs from R&D Systems (sequences patented; Minneapolis, MN, U.S.A.). Product sizes were as follows: glyceraldehyde-3-phosphate dehydrogenase, 576 bp; IL-2, 357 bp; IFN-γ, 453 bp; TNF-α, 414 bp; IL-12p40, 559 bp; IL-4, 300 (long isoform) ± 252 (short isoform) bp; and IL-5, 271 bp. Thermal cycler settings were 94° for 15 min and 40 cycles of 94° for 45 s, 55° for 45 s, and 72° for 45 s, followed by 72° for 10 min. PCR products were analyzed by 2% agarose gel electrophoresis and photographed with BioRad Fluor-S MultiImager (Hercules, CA, U.S.A.) with a 15- s exposure. All PCR work was repeated twice for each sample.
Fluorescent in situ hybridization.
Snap-frozen tissue was cut into 7-μm sections, fixed in ice-cold 4% paraformaldehyde, and incubated with hydrogen peroxide followed by 100% ethanol. Digoxin-labeled, cytokine-specific (IL-2, IFN-γ, TNF-α) antisense oligonucleotide probes (GeneDetect, Auckland, New Zealand) were applied to the sections and incubated for 2 d in a 37° humidified chamber. After extensive washing with SSC/DTT, the tissue was blocked with 20% human serum followed by application of anti–digoxin-HRP (Roche Laboratories, Indianapolis, IN, U.S.A.) for 1 h. Cyanine 3–tyramide system amplification substrate was then added followed by counterstaining with DAPI (nuclear stain). The sections were air-dried, and Vectashield and a coverslip were applied. Photographs were taken as described above.
RESULTS
Subject characteristics.
Liver tissue was analyzed from patients with biliary atresia at the time of Kasai portoenterostomy (n = 10) or with NH (n = 3), TPN-related cholestasis (n = 4), CDC (n = 4), and normal livers (n = 4). Table 1 presents demographic data and serum bilirubin, GGTP, and ALT values from each patient group. All patients with biliary atresia, NH, TPN-related cholestasis, and CDC had conjugated hyperbilirubinemia and elevated serum GGTP and ALT levels. Histology of biliary atresia, NH, and TPN-related cholestasis revealed marked bile ductular proliferation with bile plugs and varying degrees of portal tract fibrosis. CDC liver tissue showed mild ductal proliferation, and normal liver tissue showed no pathologic findings.
Increase in CD8+ and CD4+ T cells and Kupffer cells in the portal tracts of patients with biliary atresia.
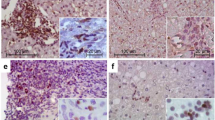
We determined the cell types present in the portal tracts of patients with biliary atresia and control subjects using fluorescent immunohistochemical staining. Portal tracts were identified by characteristic vessel formation and bile duct epithelium and confirmed by cytokeratin 7 staining. Immunohistochemical analysis of cell surface expression of CD4 (T cell), CD8 (T cell), CD20 (B cell), CD68 (macrophage/Kupffer cell), and NK1 was performed. It is known that both Kupffer cells and sinusoidal cells also have surface expression of CD4 (25). Therefore, to determine whether the CD4+ cells were in fact T cells, we performed double staining for expression of CD3 (present on all T cells). Surface expression staining revealed extensive infiltration of CD3+4+ and CD8+ T cells as well as Kupffer cells in the portal tracts of patients with biliary atresia compared with normal livers and the cholestatic disease controls (Fig. 1). There was no observable difference in the expression of CD20+ cells or NK1+ cells among the groups, with only a few portal tract CD20+- and NK1+-positive cells (not shown). Isotype controls were negative for cell-specific surface staining. Quantitative analysis of portal tract infiltrates was performed for each cell type (Fig. 2). The portal areas in biliary atresia contained significantly greater (p < 0.01) mean numbers of CD3+4+, CD8+, and CD68+ cells compared with normal livers or other infantile cholestatic diseases.

Fluorescent immunohistochemistry reveals an increase in CD8+ T cells, CD3+/CD4+ T cells, and Kupffer cells in the portal tracts in biliary atresia. Frozen sections from normal liver (n = 4), other neonatal cholestatic liver diseases (TPN, n = 4; CDC, n = 1; NH, n = 3), and biliary atresia (n = 8) were incubated with MAb, amplified with fluorescein-labeled substrate (green), and counterstained with nuclear stain DAPI (blue; ×200 magnification). Portal tracts were identified by characteristic vessel formation and bile duct epithelial nuclear staining and verified by serial sections with cytokeratin 7 staining, which is specific for bile duct epithelium. In normal livers, the portal tracts are seen in the center of each photograph. In other cholestatic diseases and biliary atresia, the portal tracts are expanded and encompass the entire field at ×200 magnification. (A) Portal tract regions were verified by bile duct epithelium surface expression of cytokeratin 7. (B) CD8+ T cells are more abundant in biliary atresia portal tracts. (C) CD4+ cell surface expression is found not only on T cells but also on sinusoidal cells and Kupffer cells. For delineating the CD4+ T cells, the tissue was double stained with anti-CD3 + antibody (T cell–specific marker; red) and CD4+ antibody (green). Double-positive cells (CD3+CD4+) are yellow. There is an increase in the number of CD3+ CD4+ cells in biliary atresia portal tracts. The single-stained red cells (CD3+CD4+) in biliary atresia reflect the CD8+ T cells. Also note the single staining CD4+ sinusoidal cells in the parenchyma surrounding the portal tract in normal liver controls. (D) CD68+ cells (macrophage/Kupffer cells) are increased in the portal tracts of biliary atresia. Note the CD68+ staining of cells in the parenchyma of controls.

Quantification of cell number in portal tracts reveals significant increases in CD8+, CD3+CD4+, and CD68+ cells in biliary atresia. Frozen sections from normal liver (n = 4), other cholestatic liver diseases (n = 8), and biliary atresia (n = 8) were analyzed for total number of cells per portal tract staining positive for CD8+ (A), CD3+ CD4+ (B), CD68+ (C), CD20+ (B cell; D), and NK1 (E). Every portal tract from each frozen section was analyzed. The number of portal tracts per section ranged from two to seven. Mean cell counts per portal tract from each patient group are shown with SDs. Statistically significant (p < 0.01) increases in CD8+, CD3+CD4+, and CD68+ cells were found in the portal tracts of biliary atresia patients compared with both normal liver controls and other neonatal cholestatic diseases.
Th1-type inflammatory cytokines in biliary atresia.
We next analyzed the expression of cytokines induced in the livers of patients with biliary atresia and control subjects with RT-PCR. As shown in Figure 3, biliary atresia tissue was associated with mRNA expression characteristic of Th1-type inflammatory cytokines: IL-2 (8 of 10 biliary atresia specimens), IFN-γ (7 of 10), TNF-α (10 of 10), and IL-12 (7 of 10). Only one sample was positive for IL-4, and none was for IL-5 expression. Normal liver tissue, NH, and CDC liver tissue did not show a Th1 cytokine profile. Normal liver tissue was negative for IL-2, IFN-γ, IL-12, and IL-5. Faint expression of TNF-α was found in one control and IL-4 in two controls. CDC liver tissue was likewise negative for IL-2, IFN- γ, IL-12, and IL-5. One specimen was positive for TNF-α, and two of four specimens were positive for IL-4. One specimen from the NH group was positive for TNF-α and IL-4; the other two samples revealed no cytokine expression. It is interesting that TPN-related CDC specimens showed a Th1 inflammatory cytokine profile similar to that seen in biliary atresia with mRNA expression of IL-2 (two of four specimens), IFN- γ (three of four), TNF-α (three of four), and IL-12 (two of four). IL-4 was found in one specimen, and there was no expression of IL-5.

Biliary atresia is associated with a Th1 inflammatory cytokine profile. RNA from liver of biliary atresia patients (BA, n = 10), normal controls (N; n = 4), CDC patients (n = 4), NH (n = 3), and TPN-related cholestasis patients (n = 4) were analyzed for the presence of cytokine-specific mRNA transcripts by RT-PCR. Th1 cytokine (IL-2, IFN-γ, TNF-α, and IL-12) and Th2 cytokine (IL-4 and IL-5) PCR products are shown for each patient. Biliary atresia and TPN-related cholestasis were associated with a Th1 inflammatory cytokine profile that was not present in normal, NH, or CDC livers.
Increases in inflammatory cells in the parenchyma of TPN-related cholestasis compared with normal livers or biliary atresia.
As described above, portal tract infiltrates in biliary atresia had significant increases in CD4+ and CD8+ T cells as well as Kupffer cells (CD68+) compared with patients with TPN-related cholestasis, yet both biliary atresia and TPN-related cholestasis were associated with a Th1 inflammatory cytokine profile. To determine further the localization of specific inflammatory cells in TPN-related cholestasis, histochemical analysis was performed of the intrahepatic lobules, revealing a marked increase in cell surface staining for CD8+ and CD68+ cells in the parenchyma of TPN-related cholestatic livers compared with the other groups (Fig. 4). There was no observable difference among the three groups in the number of CD4+, CD20+, or NK1+ cells in the hepatic lobule (data not shown). Thus, it seemed that the inflammatory cells in the hepatic lobule of TPN-related cholestasis were most likely responsible for the Th1 cytokine production. Therefore, further analysis using in situ hybridization was performed to localize the Th1 cytokine–producing cells in biliary atresia and TPN-related cholestasis.

Fluorescent immunohistochemistry reveals an increase in CD8+ and CD68+ cells in the parenchyma of TPN-related cholestasis. Frozen sections from normal livers, TPN-related cholestasis, and biliary atresia were analyzed for cell surface expression of CD4, CD8, CD20, CD68, and NK1 within the parenchyma of the liver. Increased expression of CD8+ and CD68+ cells was observed in the parenchyma (hepatic lobule) of patients with TPN-related cholestasis compared with both normal and biliary atresia livers (×100 magnification). There was no observable difference among the groups in the expression of CD4+, CD20+, or NK1 cells in the parenchyma (data not shown).
Th1-cytokine production by portal tract inflammatory cells in biliary atresia but not TPN-related cholestasis.
Biliary atresia and TPN-related cholestasis liver specimens, which were positive by RT-PCR for expression of Th1 cytokines, were analyzed for localization of cytokine-producing cells. Frozen sections were hybridized with fluorescent-tagged cytokine-specific oligonucleotide probes. Control tissue without exposure to the probes was negative for fluorescent signal. All biliary atresia specimens analyzed revealed marked expression of IL-2, IFN-γ, and TNF-α confined to cells within the portal tracts. In contrast, all TPN-related cholestasis specimens analyzed showed expression of IL-2, IFN-γ, and TNF-α in cells within the parenchyma and not in the portal tracts. Figure 5 shows an example of Th1 cytokine expression in the portal tracts of biliary atresia and the lack of expression of these cytokines in the portal tracts of TPN-related cholestasis (parenchyma staining not shown). The cytokine mRNA expression studies combined with the in situ hybridization findings clarified that the Th1 cellular inflammatory portal tract environment was exclusive to biliary atresia and not found in normal livers or the other two cholestatic liver diseases.

In situ hybridization reveals that cells in the portal tracts of biliary atresia produce Th1 cytokines. Frozen sections of biliary atresia and TPN-related cholestatic liver tissue were hybridized with cytokine-specific, digoxin-labeled antisense probes followed by anti-digoxin/HRP conjugation and cyanine 3 (red) labeled substrate. Sections were counterstained with the nuclear stain DAPI (blue) and viewed by fluorescent microscopy (×200 magnification). Expression for IL-2, IFN-γ, and TNF-α mRNA was present in the portal tracts of biliary atresia liver tissue but was absent in the portal tracts of TPN-related cholestatic liver tissue.
DISCUSSION
The results of this study demonstrate that the portal tract cellular infiltrate present in biliary atresia at the time of diagnosis represents a CD4+ Th1 cell–mediated inflammatory process. Within biliary atresia livers, the portal tracts are infiltrated with a characteristic inflammatory cell population, consisting of Kupffer cells, CD4+ T cells, and CD8+ T cells with local production of IL-2, IFN-γ, and TNF-α. This process is distinctive to biliary atresia and was not found in liver from normal control subjects or CDC patients (representing another cause of extrahepatic bile duct obstruction) or from infants with NH or TPN-related cholestasis (representing similar portal tract cholestatic histology in infancy). The absence of this inflammatory process in other neonatal cholestatic conditions suggests that the portal tract inflammation in biliary atresia is not simply an immune response to the presence of cholestasis but rather is an important pathophysiologic event involved in the pathogenesis of biliary atresia.
Our hypothesis that Th1 cell–mediated inflammation is central to the pathogenesis of biliary atresia is supported by recent studies. Urushihara et al. (18) observed increased numbers and size of Kupffer cells in the liver associated with elevated serum IL-18 at the time of diagnosis of biliary atresia. IL-18 (IFN-γ–inducing factor) is a macrophage-derived cytokine that works in concert with IL-12 to promote Th1 cell differentiation in the inflammatory setting. Bezerra et al. (26) used microarray analysis to show that liver of infants with biliary atresia expressed several genes involved in lymphocyte differentiation, with an overexpression of osteopontin, another cytokine in the Th1 pathway. They further determined by RT-PCR that 65% of biliary atresia livers expressed IFN-γ compared with none of controls with idiopathic neonatal intrahepatic cholesta-sis. However, tissue localization of cells expressing IFN-γ was not possible with the techniques used, no other cytokines were investigated, and the only controls were infants with idiopathic neonatal hepatitis, unlike the CDC control subjects in the present study.
The predominance of CD8+ T cells in the portal tracts of biliary atresia livers supports a role for cytotoxic CD8+ T cells in mediating the bile duct epithelial damage. In this regard, Ohi et al. (23) examined CD8+ T cells in biliary atresia livers and found that CD8+ T cells infiltrated the proliferating portal tract bile ducts in 95% of the biliary atresia cases. However, they concluded that the CD8+ T cells were not actively cytotoxic because immunohistochemistry staining for granzyme, perforin, and FasL were negative. However, CD8+ cytotoxic T cells also produce cytokines and chemokines with effector functions that do not directly involve target cell death. For example, CD8+ T cells produce IFN-γ and TNF-α and the chemokines MIP-1α, MIP-1β, and RANTES. Taken together, these molecules mediate the promotion of inflammatory responses, activation of macrophages, and direct antiviral effects to infected cells (27). The role of the CD8+ T cells in the ongoing destruction of bile duct epithelium in biliary atresia warrants further investigation.
Our findings support the role of immune-mediated destruction of bile ducts within the liver of patients with the perinatal/acquired form of biliary atresia. This study adds to the observations of Ohya et al. (28), who described an association in biliary atresia between degeneration of intrahepatic bile ducts and the lymphocytic infiltration of biliary epithelial cells. In that study, 31 patients with biliary atresia had these findings compared with only 2 of 9 patients with CDC and none of 5 patients with idiopathic neonatal hepatitis. From our study, it can be inferred further that this lymphocytic infiltrate is primarily composed of CD4+ and CD8+ T cells. Further investigation will be needed to determine whether the lymphocytes are clonal and reactive to a specific antigen (e.g. viral or self-bile duct epithelial antigen) and the nature of the antigenic epitopes.
The findings in this study may shed light on the potential therapeutic use of immunosuppressive therapy in the treatment of biliary atresia. Recent noncontrolled studies using corticosteroid therapy after the Kasai procedure suggested an improved overall survival rate compared with historical controls (29–31). First, corticosteroids regulate the expression of many genes with a net anti-inflammatory effect, including a reduction in inflammatory mediator secretion, such as TNF-α and nitric oxide. Second, corticosteroids inhibit inflammatory cell migration to sites of inflammation by inhibiting the expression of adhesion molecules (32). The ultimate result of these anti-inflammatory effects would be to inhibit further bile duct injury and to slow the development of biliary cirrhosis. Carefully conducted controlled studies are needed to determine the effects of corticosteroid therapy on immune injury and outcome in biliary atresia.
In conclusion, this study offers strong evidence for a Th1-mediated immune response focused within the portal tracts in biliary atresia that is not present in other, similar neonatal cholestatic disorders. This unique immune pattern suggests that the observed immune repertoire is not merely a stereotypic response to the accumulation of biliary constituents in the neonatal cholestatic liver but rather an indicator of specific immune processes involved in the pathogenesis of biliary atresia. Further elucidation of the nature and the source of antigenic stimulation of the Th1 response in the biliary tree of these patients will be essential for advancing our understanding of this disorder and developing new strategies toward effective treatment.
Abbreviations
- CDC:
-
choledochal cyst
- HRP:
-
horseradish peroxidase
- IFN:
-
interferon
- NH:
-
neonatal giant cell hepatitis
- NK:
-
natural killer
- TNF:
-
tumor necrosis factor
- TPN:
-
total parenteral nutrition
References
Stein JE, Vacanti JP 1994 Biliary atresia and other disorders of the extrahepatic biliary tree. In: Suchy FJ (ed) Liver Disease in Children. Mosby, London, pp 426–442.
Tan CEL, Driver M, Howard ER, Moscoso GJ 1994 Extrahepatic biliary atresia: a first trimester event? Clues from light microscopy and immunohistochemistry. J Pediatr Surg 29: 808–814.
Horwitz MS, Sarvetnick N 1999 Viruses, host responses and autoimmunity. Immunol Rev 169: 241–253.
Sokol RJ, Mack CL 2001 Etiopathogenesis of biliary atresia. Semin Liver Dis 21: 517–524.
Schreiber RA, Kleinman RE 1993 Genetics, immunology and biliary atresia: an opening or a diversion?. J Pediatr Gastroenterol Nutr 16: 111–113.
Morecki R, Glaser JH, Cho S, Balistreri WF, Horwitz MS 1982 Biliary atresia and reovirus type 3 infection. N Engl J Med 307: 481–484.
Morecki R, Glaser JH, Johnson AB, Kress Y 1984 Detection of reovirus type 3 in the porta hepatis of an infant with extrahepatic biliary atresia: ultrastructural and immunocytochemical study. Hepatology 4: 1137–1142.
Tyler KL, Sokol RJ, Oberhaus SM, Le M, Karrer FM, Narkewicz MR, Tyson RW, Murphy JR, Low R, Brown WR 1998 Detection of reovirus RNA in hepatobiliary tissues from patients with extrahepatic biliary atresia and choledochal cysts. Hepatology 27: 1475–1482.
Brown WR, Sokol RJ, Levin MJ, Silverman A, Tamaru T, Lilly JR, Hall RJ, Cheney M 1998 Lack of correlation between infection with reovirus 3 and extrahepatic biliary atresia or neonatal hepatitis. J Pediatr 113: 670–676.
Steele MI, Marshall CM, Lloyd RE, Randolph VE 1995 Reovirus 3 not detected by reverse transcriptase mediated polymerase chain reaction analysis of preserved tissue from infants with cholestatic liver disease. Hepatology 21: 697–702.
Riepenhoff-Talty M, Gouvea V, Evans MJ, Svensson L, Hoffenberg E, Sokol RJ, Uhnoo I, Greenberg SJ, Schakel K, Zhaori G, Fitzgerald J, Chong S, el-Yousef M, Nemeth A, Brown M, Piccoli D, Hyams J, Ruffin D, Rossi T 1996 Detection of group C rotavirus in infants with extrahepatic biliary atresia. J Infect Dis 174: 8–15.
Bobo L, Ojeh C, Chiu D, Machado A, Colombani P, Schwarz K 1997 Lack of evidence for rotavirus by polymerase chain reaction/enzyme immunoassay of hepatobiliary samples from children with biliary atresia. Pediatr Res 41: 229–234.
Fischler B, Ehrnst A, Forsgren M, Orvell C, Nemeth A 1998 The viral association of neonatal cholestasis in Sweden: a possible link between cytomegalovirus infection and extrahepatic biliary atresia. J Pediatr Gastroenterol Nutr 27: 57–64.
Jevon GP, Dimmic JE 1999 Biliary atresia and cytomegalovirus infection: a DNA study. Pediatr Dev Pathol 2: 11–14.
Bill AH, Haas JE, Foster GL 1977 Biliary atresia: histopathologic observations and reflections upon its natural history. J Pediatr Surg 12: 977–982.
Haas JE 1978 Bile duct and liver pathology in biliary atresia. World J Surg 2: 561–569.
Kobayashi H, Puri P, O'Brian DS, Surana R, Miyano T 1997 Hepatic overexpression of MHC class II antigens and macrophage-associated antigens (CD68) in patients with biliary atresia of poor prognosis. J Pediatr Surg 32: 590–593.
Urushihara N, Iwagaki H, Yagi T, Kohka H, Kobashi K, Morimoto Y, Yoshino T, Tanimoto T, Kurimoto M, Tanaka N 2000 Elevation of serum interleukin-18 levels and activation of Kupffer cells in biliary atresia. J Pediatr Surg 35: 446–449.
Tracy TF, Dillon P, Fox ES, Minnick K, Vogler C 1996 The inflammatory response in pediatric biliary disease: macrophage phenotype and distribution. J Pediatr Surg 31: 121–125.
Broome U, Nemeth A, Hultcrantz R, Scheynius A 1997 Different expression of HLA-DR and ICAM-1 in livers from patients with biliary atresia and Byler's disease. J Hepatol 26: 857–62.
Davenport M, Gonde C, Redkar R, Koukoulis G, Tredger M, Mieli-Vergani G, Portmann B, Howard ER 2001 Immunohistochemistry of the liver and biliary tree in extrahepatic biliary atresia. J Pediatr Surg 36: 1017–1025.
Eagar TN, Tompkins SM, Miller SD 2001 Helper T-cell subsets and control of the inflammatory response. In: Rich RR, Fleisher TA, Shearer WT, Kotzin BL, Schroeder HW Jr (eds) Clinical Immunology Principles and Practice. Mosby, London, pp 16.1–16.12
Ahmed AF, Ohtani H, Nio M, Funaki N, Shimaoda S, Nagura H, Ohi R 2001 CD8+ T cells infiltrating into bile ducts in biliary atresia do not appear to function as cytotoxic T cells: a clinicopathological analysis. J Pathol 193: 383–389.
Desmet VJ 2001 The cholangiopathies. In: Suchy FJ, Sokol RJ, Balistreri WF (eds) Liver Disease in Children. Lippincott Williams & Wilkins, Philadelphia, pp 39–62.
Knolle PA, Gerken G 2000 Local control of the immune response in the liver. Immunol Rev 174: 21–34.
Bezerra JA, Tiao G, Ryckman FC, Alonso M, Sabla GE, Shneider B, Sokol RJ, Aronow BJ 2002 Genetic induction of proinflammatory immunity in children with biliary atresia. Lancet 360: 1653–1659.
Altman JD, Lippolis JD 2001 Cytotoxic T-cell function. In: Rich RR, Fleisher TA, Shearer WT, Kotzin BL, Schroeder HW Jr (eds) Clinical Immunology Principles and Practice. Mosby, London, pp 17.1–17.9
Ohya T, Fujimoto T, Shimomura H, Miyano T 1995 Degeneration of intrahepatic bile ducts with lymphocyte infiltration into biliary epithelial cells in biliary atresia. J Pediatr Surg 30: 515–518.
Muraji T, Higashimoto Y 1997 The improved outlook for biliary atresia with corticosteroid therapy. J Pediatr Surg 32: 1103–1107.
Dillon PW, Owings E, Cilley R, Field D, Curnow A, Georgeson K 2001 Immuno-suppression as adjuvant therapy for biliary atresia. J Pediatr Surg 36: 80–85.
Meyers RL, Book LS, O'Gorman MA, Jackson WD, Black RE, Johnson DG, Matlak ME 2003 High-dose steroids, ursodeoxycholic acid, and chronic intravenous antibiotics improve bile flow after Kasai procedure in infants with biliary atresia. J Pediatr Surg 38: 406–411.
Janeway CA, Travers P, Walport M, Shlomchik 2001 Immunobiology 5: The Immune System in Health and Disease. Garland Publishing, New York, pp 554–555.
Author information
Authors and Affiliations
Corresponding author
Additional information
This study was supported by the National Institutes of Health Grant KO8 DK60710-02, National Institute of Neurological Disorders and Stroke Grant R01NS-23349, Glaxo-SmithKline Institute for Digestive Health 2002 Basic Science Research Award, Johnny Jenna Foundation (Brookfield, IL, U.S.A.), and Liver Foundation for Kids (Lemont, IL, U.S.A.).
Rights and permissions
About this article
Cite this article
Mack, C., Tucker, R., Sokol, R. et al. Biliary Atresia Is Associated with CD4+ Th1 Cell–Mediated Portal Tract Inflammation. Pediatr Res 56, 79–87 (2004). https://doi.org/10.1203/01.PDR.0000130480.51066.FB
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/01.PDR.0000130480.51066.FB
