
Abstract
Glucocorticosteroid hormones induce apoptosis in lymphocytes. Therefore, glucocorticoids are commonly used as immunosuppressive and chemotherapeutic agents. This review examines many facets of the process by which glucocorticoids induce apoptosis. This process is divided into three stages, an initiation stage that involves glucocorticoid receptor-mediated gene regulation, a decision stage that involves the counterbalancing influence of prosurvival and proapoptotic factors, and the execution stage which involves caspase and endonuclease activation. Many aspects of glucocorticoid-induced apoptosis, such as mitochondrial dysfunction and caspase activation, are important steps in virtually all forms of apoptosis. But the process glucocorticoid-induced apoptosis differs from other forms of apoptosis in terms of initiation at the transcriptional level and involvement of the multicatalytic proteasome and calcium. Moreover, the abundant opportunity for crosstalk between the glucocorticoid receptor and other signaling pathways increases the complexity of glucocorticoid-induced apoptosis and its regulation.
Similar content being viewed by others

Introduction
The induction of apoptosis in thymocytes by glucocorticosteroid hormones is one of the earliest recognized forms of apoptosis.1 But progress in understanding corticosteroid-induced apoptosis has lagged behind remarkable advances in understanding other forms of apoptosis, such as that induced by the ligands of death receptors (e.g., Fas/CD95, tumor necrosis factor). In part, this is because corticosteroid-induced apoptosis is initiated by a steroid receptor-mediated change in gene expression, but specific genes that mediate cell death in response to corticosteroid treatment have not been identified.
Recent findings have shed light on events in the cell death pathway downstream from gene regulation, including the caspases responsible for the execution phase of cell death, the unexpected involvement of the multicatalytic proteasome in the death process, the suppression of prosurvival transcription factors (e.g., AP-1, c-myc, NF-κB), and crosstalk between T cell receptor and cytokine signaling pathways. Moreover, evidence that mitochondrial dysfunction lies downstream of caspase activation in corticosteroid-induced apoptosis challenges the generalization that mitochondrial dysfunction is the central decision point in all forms of apoptosis.
This review focuses primarily on insights gained in recent years regarding the mechanism of corticosteroid-induced apoptosis. Ultimately, the goal of investigating this form of apoptosis is to better understand the role of corticosteroid hormones in physiological regulation of the immune system and in the treatment of lymphoid leukemias and lymphomas.
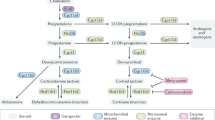
To organize this review, corticosteroid-induced apoptosis is arbitrarily divided into three stages: the initiation stage, the decision stage, and the execution stage (Figure 1). The initiation stage involves corticosteroid receptor-mediated changes in gene expression, while the execution stage involves activation of caspases, or other proteases and endonucleases. The decision stage includes those events that ultimately result in a commitment to cell death. A decision to die appears to be made by counterbalancing actions of cytokines, which induce transcription factors that promote cell proliferation and survival, versus the actions of corticosteroids, which oppose the expression of these transcription factors. An important feature of corticosteroid-induced apoptosis is involvement of the multicatalytic proteasome, which appears to play an active role in promoting cell death by degrading factors necessary for cell survival.

Stages of glucocorticoid-induced apoptosis. Between the Initiation Stage and the Execution Stage lies the Decision Stage in which counterbalancing factors determine the commitment of the cell to live or die. Recent findings indicate that repression of prosurvival transcription factors and proteasome-mediated degradation of apoptosis inhibitory factors may play a crucial role in this stage
The initiation stage
Glucocorticoid receptor mediated changes in gene transcription
In this review, the initiation stage is defined as the sequence of events leading up to and including direct regulation of gene transcription by the corticosteroid receptor (Figure 2). But what genes are regulated so that the lymphoid cell ultimately dies? The answer to this question has evaded investigators up to the present time. In part, the difficulty has been knowing what to look for: the induction of a gene or set of genes that produce cell death as a direct or indirect consequence of their biological action, or the repression of genes necessary for maintenance of cell survival, or a combination of both? One specific point of confusion in understanding corticosteroid-induced apoptosis has to do with whether the transactivation or transrepression functions of the corticosteroid receptor initiate the process of cell death. Evidence in favor of each is summarized here.

The Initiation Stage. There is considerable evidence that glucocorticoid-induced apoptosis is initiated by the transactivation activity of the glucocorticoid receptor, but the specific genes regulated in this process are not yet identified. RAP46/BAG1 represses the transactivation activity of the glucocorticoid receptor, and therefore inhibits glucocorticoid-induced apoptosis. Glucocorticoid-induced apoptosis is also repressed by T cell receptor (TCR) signaling, which appears to alter the transactivation activity of the glucocorticoid receptor. On the other hand, DAP3 increases glucocorticoid-induced apoptosis by enhancing the transactivation activity of the glucocorticoid receptor
Evidence for involvement of the glucocorticoid receptor transrepression activity in glucocorticoid-induced apoptosis
The concept that corticosteroid-induced apoptosis is mediated by the transrepression activity of the glucocorticoid receptor was largely based on the work of Helmberg et al.2 These investigators reported that mutant glucocorticoid receptors defective in transactivation function, but fully competent at interfering with AP-1 (activating protein-1) activity, mediated apoptosis induction by corticosteroids. Since the AP-1 transcription factor regulates expression of genes involved in cell growth, differentiation and transformation,3,4 their findings suggested that interference with transcription factors required for cell survival, such as AP-1 (activating protein-1), may contribute to apoptosis induction.2 This interpretation is consistent with earlier findings indicating that in cytokine (IL-2)-dependent CTLL cells there is a correlation between the presence of AP-1 DNA binding activity and repression of corticosteroid-induced apoptosis.5 In the absence of interleukin-2, corticosteroid treatment stimulates AP-1 degradation and induces apoptosis.5
The repression of AP-1 transcription factor activity in corticosteroid-treated lymphocytes is accompanied by repression of other prosurvival transcription factors, including the nuclear factor of activated T lymphocytes (NFAT).6 NFAT is composed of two components, NFATp and AP-1; the dexamethasone-induced decrease in NFAT activity is due to a decrease in AP-1 activity.6 Also, Thompson and coworkers7 have provided substantial evidence that downregulation of c-Myc accompanies the induction of apoptosis by corticosteroids, and that c-Myc overexpression inhibits corticosteroid-induced apoptosis.
Evidence for involvement of the glucocorticoid receptor transactivation activity in glucocorticoid-induced apoptosis
Recent evidence from several laboratories indicates that the transactivation activity of the glucocorticoid receptor is required to mediate corticosteroid-induced apoptosis. Chapman et al 8 found that chimeric receptors containing the potent VP16 and E1A viral transactivation domains in place of the corticosteroid receptor amino terminus enhanced the sensitivity of lymphoid cells to corticosteroid-induced apoptosis, supporting a role for transactivation in apoptosis induction. Ramdas et al 9 addressed the question by comparing human leukemic T cells that express either wild type corticosteroid receptor or receptor that has a mutation that is activation deficient, but that retains the ability to repress AP-1 activity. This mutant failed to mediate corticosteroid-induced apoptosis, indicating that the transactivation function of the receptor is essential for corticosteroid-induced apoptosis. In a similar approach in vivo, Reichardt et al10 engineered a point mutation in the corticosteroid receptor in mice that impairs corticosteroid receptor dimerization, and hence inactivates the transactivation function of the receptor. In mice with this receptor mutation, corticosteroid-induced apoptosis of thymocytes was inhibited. Since this mutant receptor retains transrepression activity, these studies provide strong evidence that the transactivation function of the corticosteroid receptor is necessary for corticosteroid-induced apoptosis.
Additional evidence for the importance of corticosteroid receptor transactivation function was provided by experiments using RAP46 to inhibit receptor function.11 RAP46 binds to the receptor hinge region and inhibits DNA binding and transactivation by the receptor. Overexpression of RAP46 in S49 mouse lymphoma cells inhibited corticosteroid-induced apoptosis. Conversely, corticosteroid-induced apoptosis and transactivation were enhanced after treating S49 cells with the immunosuppressive rapamycin, which down-regulates the cellular levels of BAG-1, the mouse homolog of RAP46. These findings identify RAP46 as a protein that controls corticosteroid-induced apoptosis through negative regulatory action on the transactivation property of the corticosteroid receptor. In the reverse situation, the proapoptotic death-associated protein (DAP3) associates with the corticosteroid receptor and promotes corticosteroid-induced apoptosis by stimulating ligand-induced transcriptional activation by the receptor.12
What genes mediate glucocorticoid-induced apoptosis?
The target genes whose transactivation or transrepression initiates the cell death process remain uncertain. Candidate genes may come from unexpected sources. For example, one potential clue has been provided by the observation that thymocytes from mice homozygous for the autosomal recessive mutation Wasted (wst/wst) have increased sensitivity to corticosteroid-induced apoptosis.13 It has been suggested that elevated thymocyte apoptosis may be a major contributor to the lymphoid dysfunction and ultimate death in wst/wst mice. Identification of the mutant gene and its function in mice with the Wasted phenotype may possibly reveal insight into genes critical for induction of apoptosis in thymocytes.
The glucocorticoid receptor, a zinc-finger transcription factor, is under autoregulatory control and its level is increased following corticosteroid treatment in human T-leukemia cells.14 Although corticosteroid receptor upregulation appears to be an important component of cell death induction, its mechanism of action in this regard is presently unclear. Whether or not corticosteroid receptor upregulation might contribute to repression of prosurvival transcription factors is presently unknown, although this is an intriguing possibility in view of recognized cross-talk between the corticosteroid receptor and other transcription factor complexes.15,16
The decision stage
Multicatalytic proteasome
Recent findings have generated considerable interest in the role of the multicatalytic proteasome in corticosteroid-induced apoptosis. The proteasome is a multicatalytic protease complex located in both the cytoplasm and nucleus that degrades proteins targeted for destruction by polyubiquitination.17,18,19 Although much less is known about the role of the proteasome in apoptosis, compared to the vast body of information regarding caspases, evidence in primitive organisms has suggested a fundamental involvement of the proteasome in apoptosis. For example, in the hawkmoth, Manduca sexta, apoptosis of intersegmental muscles is associated with elevated ubiquitin gene expression and proteasome activity.20,21,22
Genetic evidence for involvement of the proteasome in apoptosis has been provided by investigation of a dominant mouse mutation ‘fused toes’, characterized by partial syndactyly of the limbs and thymic hyperplasia.23 Both morphological abnormalities are attributed to impaired regulation of programmed cell death, due to decreased expression of a novel gene, Ft1, which encodes a protein related to ubiquitin-conjugating enzymes.
In mammalian cells, evidence that proteasome activity plays a role in apoptosis induction comes primarily from work on the induction of apoptosis in thymocytes by corticosteroids. Grimm et al 24 demonstrated that proteasome inhibitors block corticosteroid-induced cleavage of poly-ADP ribose polymerase (PARP), a downstream caspase target, and apoptosis in thymocytes (Figure 3). These findings raised the possibility that the proteasome may either degrade regulatory proteins that normally inhibit the apoptotic pathway, or may proteolytically activate proteins that promote cell death.

Proteasome inhibitors block glucocorticoid-induced apoptosis. Pretreatment of thymocytes with the proteasome inhibitors lactacystin or MG132 inhibits glucocorticoid-induced apoptosis
In corticosteroid-induced apoptosis of thymocytes, proteasome activity appears to be involved at a step preceding mitochondrial changes and caspase activation. Beyette et al 25 have shown that thymocytes are rich in proteasomes, and that a chymotryptic component of proteasome activity decreases following dexamethasone treatment. Moreover, proteasome inhibitors prevented disruption of the mitochondrial transmembrane potential and also prevented exposure of phosphatidylserine and nuclear DNA fragmentation.26 Pharmacologic stabilization of the mitochondrial permeability transition pore, or inhibition of caspases, did not prevent the activation of proteasomes.26,27 Proteasome activation in dexamethasone-treated thymocytes is inhibited by overexpression of Bcl-2, as well as by protein synthesis inhibitors and antioxidants.27 Involvement of the proteasome in apoptosis induction is a distinguishing characteristic of the corticosteroid-induced death pathway versus the Fas-mediated cell death pathway where the proteasome appears dispensable.27
Although proteasome activity appears to play a proapoptotic role in corticosteroid-treated lymphocytes, proteasome targets in this context are just now being identified. One of these targets, as mentioned in the preceding section, is the transcription factor c-Fos. In a T cell lymphoma line, degradation of the transcription factor c-Fos by the proteasome was found to be a relatively early step in corticosteroid-induced apoptosis that preceded caspase-3 activation and DNA fragmentation and was inhibited by Bcl-2 overexpression.28 A mutant form of c-Fos that evades degradation by the proteasome inhibited corticosteroid-induced apoptosis, suggesting that c-Fos degradation contributed to apoptosis induction.28 Moreover, Ivanov et al 29 found that the inhibition of corticosteroid-induced apoptosis in thymocytes by the proteasome-specific inhibitor lactacystin is associated with stabilization of AP-1, NF-κB and NUR-77 against proteasome-mediated degradation.
The role of the proteasome in corticosteroid-induced apoptosis is not limited to the degradation of prosurvival transcription factors. Another concept regarding the role of the proteasome in corticosteroid-induced apoptosis comes from the work of Grassilli et al.30 They found that polyamine levels decrease in thymocytes undergoing apoptosis in response to dexamethasone treatment due to accelerated degradation of ornithine decarboxylase by the proteasome. Inhibition of proteasome function preserves polyamine synthesis in association with inhibition of corticosteroid-induced apoptosis.30
Another target of the proteasome in corticosteroid-induced apoptosis is the cyclin dependent kinase inhibitor, p27Kip1.31 Following treatment with dexamethasone, the level of p27Kip1 in thymocytes decreases due to proteasome-mediated degradation, a process that is inhibited by Bcl-2 overexpression and accelerated by Bax. Finally, degradation of apoptosis inhibitory proteins, c-IAP1 and XIAP, by the proteasome has been described in thymocytes undergoing apoptosis in response to dexamethasone.32 Moreover, mutant forms of IAP that are not degraded by the proteasome had an inhibitory effect on apoptosis induction by dexamethasone, suggesting that IAP degradation promotes dexamethasone-induced apoptosis in thymocytes. The IAP's appear to bind directly to caspases and inhibit their activation.33 Hence, the proteasome may contribute to apoptosis induction by abrogating the apoptosis inhibitory effect of IAP's.
In summary, the multicatalytic proteasome appears to play an important role in the decision of a lymphoid cell to die following corticosteroid treatment. Recognized targets of the proteasome include transcription factors that regulate genes necessary for cell proliferation (e.g., c-Fos), enzymes whose activity is essential for cell proliferation (e.g., ornithine decarboxylase), cell cycle regulatory proteins (e.g., p27Kip1), and proteins that normally repress caspases (e.g., IAP's). Bcl-2 has been reported to inhibit the increase in proteasome activity associated with corticosteroid-induced apoptosis, as well as the degradation of c-Fos and p27Kip1. Therefore, regulation of proteasome-mediated degradation of factors essential for cell survival may be a point of control for Bcl-2 in the apoptotic pathway.
Signaling pathway crosstalk
Cellular signaling pathways influence each other, producing complex integrating networks. Integration of multiple signals is important for signal transducers like the corticosteroid receptor that evoke different effects in different cells and physiologic settings.34 Crosstalk between other signaling pathways and the corticosteroid receptor signaling pathway modulates corticosteroid-induced apoptosis, and serves as a major determinant of the cell's decision to live or die when exposed to corticosteroids. There are two ways by which signaling crosstalk might regulate apoptosis. Activation of prosurvival signaling pathways may repress corticosteroid-induced apoptosis. Alternatively, corticosteroids may repress prosurvival signaling pathways. Hence, the balance between opposing prosurvival and prodeath signaling pathways may determine the ultimate fate of cells. Several examples of this type of signaling crosstalk are summarized here.
Survival factors produced by thymic epithelial cells suppress corticosteroid-induced apoptosis.35 More specifically, interferon alpha and interleukin-6 inhibit dexamethasone-induced apoptosis in plasma cells and myeloma cells.36,37,38 Interleukins-9, -4, and -6 inhibit apoptosis in dexamethasone-treated thymocytes and thymoma cells.39,40 Also, interleukin-15 inhibits dexamethasone-induced apoptosis in activated T and B cells41 and insulin-like growth factors protect myeloma cells from dexamethasone-induced apoptosis.42 Thus, a number of different cytokines counteract the induction of apoptosis by corticosteroids (Figure 4).

Cytokines counteract glucocorticoid-induced apoptosis. Insulin like growth factors (IGF), interferons, and interleukins have been shown to inhibit glucocorticoid-induced apoptosis
Interleukins may counteract corticosteroid-induced death signals by increasing expression of transcription factors that mediate expression of survival genes (Figure 5). Interleukin-2-mediated protection of T cell leukemia cells from dexamethasone-induced apoptosis correlates with induction of the DNA binding and transactivation functions of AP-1.43 Also, interleukin-6 and interferon alpha may repress corticosteroid-induced apoptosis by activating mitogen-activated protein kinase and phosphatidylinositol 3-kinase pathways.44

Cytokines increase AP-1 and NF-κB activities. Cytokines (e.g., IL-2) counteract glucocorticoid-induced apoptosis by inducing expression of transcription factors that regulate genes involved in cell proliferation
An example of how the balance between prosurvival signals and prodeath signals works to regulate death decisions revolves around NF-κB. The NF-κB transcription factor family is required for expression of many cytokines and the immunosuppressive action of corticosteroids is mediated, at least in part, through interference with NF-κB activity and, therefore, with inhibition of cytokine production. In human leukemic T cells, dexamethasone treatment induces synthesis of Iκ-Bα an inhibitor of NF-κB, which correlates with apoptosis induction.9 In CD4+CD8+ thymocytes in vivo, dexamethasone treatment induces expression of Iκ-Bα and Iκ-Bβ and downregulates NF-κB DNA binding activated by intrathymic signals.45 The downregulation of NF-κB DNA binding precedes cell death, suggesting that NF-κB may be important for the survival of immature thymocytes.45 The negative effect of corticosteroids on NF-κB activity is counterbalanced by the cytokines, interleukins-2 and -4, that are reported to rescue lymphocytes from corticosteroid-induced apoptosis by inhibiting I-κB induction by dexamethasone.46 In summary, this is one example of the ‘Yin and Yang’ of survival decisions. Corticosteroids promote cell death by inducing Iκ-Bα thereby inhibiting NF-κB activity; cytokines inhibit cell death by inhibiting the induction of I-κB by corticosteroids (Figure 6). Although interference with NF-κB activity in lymphocytes was shown to be mediated by corticosteroid induction of the inhibitor protein I-κB,47 subsequent studies indicate that hormone-induced IκB synthesis and inhibition of NF-κB activity are separable biochemical events and that the glucocorticoid receptor can directly repress NF-κB activity.48,49,50,51

Glucocorticoid hormones counteract the action of cytokines. The glucocorticoid hormone dexamethasone induces expression of IκB, which inhibits the function of NF-κB, a transcription factor necessary for expression of prosurvival cytokines. Recent findings indicate that the glucocorticoid receptor may directly repress NF-κB activity independent of I-κB (see text)
Another example of how corticosteroid-induced apoptosis is regulated by crosstalk with other signaling pathways involves protein kinase C (PKC).52 Protein kinase C (PKC) includes several subfamilies of enzymes including calcium-dependent protein kinase C (cPKC) and calcium-independent novel PKC (nPKC). Glucocorticoid-induced apoptosis was inhibited by non-isoform-selective PKC inhibitors but not by cPKC-specific inhibitors. Thus, nPKC isoforms appear to be involved in apoptosis induction, while activation of calcineurin and cPKC are capable of inhibiting corticosteroid-induced apoptosis. Moreover, an activator of cAMP-activated protein kinase has been reported to block dexamethasone-induced apoptosis and caspase-3 activation.53
T cell receptor signaling can also regulate corticosteroid-induced apoptosis. Engagement of the T cell receptor (TCR) induces apoptosis by a mechanism that involves induction of Fas ligand, which in turn interacts with its receptor and activates caspase-8.54 Recognition that TCR signaling and corticosteroid signaling pathways are mutually antagonistic led Jamieson et al 55 to investigate the crosstalk pathway for inhibition of corticosteroid-induced apoptosis by TCR signaling. Their findings indicate that TCR activation of the mitogen-activated protein kinase/extracellular signal regulated kinase (MEK/ERK) cascade via Ras is necessary and sufficient to inhibit corticosteroid-mediated cell death in immortalized T cells and thymocyte cell lines, and in primary T cells. Moreover, activation of various components of the TCR pathway (Ras, MEK1) altered the transcriptional regulatory activity of the corticosteroid receptor. However, another potential explanation of how Ras represses corticosteroid-induced apoptosis may involve Ras-mediated stabilization of c-Myc protein stability. As noted above, a decrease in c-Myc protein level has been implicated as a mechanism of corticosteroid-induced apoptosis. Recently, Sears et al 56 reported that Ras enhances the stability of c-Myc by inhibiting proteasome-mediated degradation.
Recent findings indicate that the glucocorticosteroid receptor binds to 14-3-3, a cytoplasmic protein that interacts with a wide range of signaling molecules.57 The corticosteroid receptor and Raf-1, a downstream effector of the signaling factor Ras, are found in a protein complex (the ‘receptosome’), providing an opportunity for the corticosteroid receptor to crosstalk with the Raf-Ras signaling pathway.58 These interactions may be involved in regulation of cell proliferation, and induction of cell death by corticosteroids, although experimental evidence to support this concept is presently lacking.
Involvement of BCL-2 family members in the cell death decision
Corticosteroid-induced apoptosis is both positively and negatively regulated by members of the Bcl-2 protein family. Bcl-2 -/- knockout mice display fulminant apoptosis of the thymus and accelerated apoptosis of thymocytes in response to dexamethasone.59 Moreover, Bcl-2 overexpression inhibits events associated with corticosteroid-induced apoptosis, including caspase activation and mitochondrial dysfunction.60,61,62
However, the mechanism by which Bcl-2 inhibits apoptosis remains uncertain, although many hypotheses have been put forth. Currently, the most prevalent theories focus on the concept that Bcl-2 acts on the outer mitochondrial membrane to preserve mitochondrial function.63,64 But this concept does not readily explain the antiapoptotic action of Bcl-2 in corticosteroid-treated thymocytes, where mitochondrial dysfunction appears not to be a central mediator of cell death.
Perhaps one of the more revealing studies of the role of Bcl-2 in corticosteroid induced apoptosis was reported by Brunet et al.65 These investigators employed the CEM C7A human leukemia line to investigate dexamethasone-induced apoptosis. Their findings indicate that loss of clonogenicity, which represents commitment to cell death, is separable from the appearance of caspase activation and subsequent apoptotic markers. Significantly, caspase inhibitors did not prevent commitment to cell death, but Bcl-2 did. These findings indicate that Bcl-2 works upstream of caspase activation to inhibit commitment to cell death, and that the survival function of Bcl-2 is not at the level of caspase inhibition.
A Bcl-2 family-regulated link between cell cycle and cell death has been proposed, based on recent evidence that Bax and Bcl-2 modulate Cdk2 (cyclin dependent kinase) activation during dexamethasone-induced apoptosis in thymocytes.31 Following treatment with dexamethasone, the level of p27Kip1 in thymocytes decreases and the level of Cdk2 kinase activity increases. Cdk2 activity is crucial for induction of apoptosis in thymocytes by corticosteroids.66 The degradation of p27Kip1 by the proteasome is regulated by the Bcl-2 family. Bcl-2 overexpression delays the degradation of p27Kip1, whereas Bax overexpression accelerates its degradation. Moreover, Cdk2 activation during apoptosis is a highly regulated process under the control of known apoptosis regulators such as Bax and Bcl-2. Bcl-2 overexpression delayed the increase in Cdk2 kinase activity associated with dexamethasone-induced apoptosis, whereas Bax overexpression had the opposite effect.
Thus, induction of thymocyte apoptosis by dexamethasone activates biochemical machinery that is shared with the normal cell cycle, i.e., Cdk2 and p27Kip1. That Bax and Bcl-2 are able to modulate the levels of p27Kip1 and Cdk2 identifies both of these factors as downstream components of a common biochemical pathway leading to apoptosis. Furthermore, these findings draw a mechanistic distinction between corticosteroid-induced apoptosis and apoptosis induction in thymocytes by Fas/CD95, as the latter does not involve changes in Cdk2 and p27Kip1 and is not inhibited by Bcl-2.31
Bcl-2 may also function by regulating proteasome-mediated degradation of prosurvival transcription factors. Two examples are c-Fos and NF-κB. Evidence that Bcl-2 inhibits dexamethasone-induced degradation of c-Fos by the proteasome, published by He et al,28 was discussed earlier. Feinman et al 67 reported that dexamethasone-induced apoptosis of multiple myeloma cells is accompanied by decreased NF-κB DNA binding activity which is preserved by Bcl-2 overexpression. In this situation, Bcl-2 did not prevent I-κB induction by dexamethasone; thus, it is possible that Bcl-2 stabilizes NF-κB by inhibiting proteasome-mediated degradation.
An intriguing question concerns the involvement of proapoptotic members of the Bcl-2 family in corticosteroid-induced apoptosis. The proapoptotic family consists of members like Bax and Bak, which share significant homology with Bcl-2, and the BH3 domain only members which have only the BH3 domain in common with Bcl-2.63 The latter family includes Bid, Bad, and Bim. A pattern of apoptosis signaling is emerging in which proapoptotic members of the Bcl-2 family transmit death signals from upstream initiator pathways to downstream effector pathways. The proapoptotic proteins are held in check by different mechanisms in the healthy cell, and then released to induce apoptosis in response to apoptotic signals.
For example, in response to engagement of death receptors by their ligands (e.g., Fas, tumor necrosis factor), Bid undergoes caspase-8-mediated cleavage which exposes its BH3 domain in the form of an active fragment that triggers mitochondrial dysfunction and cytochrome c release.68 A second example is Bax, which is located in the cytoplasm and translocated to mitochondria in response to apoptotic signals (e.g., growth factor withdrawal).63 A third example is Bad, which in the healthy cells exists as an inactive, phosphorylated form, in association with cytoplasmic 14-3-3. Apoptotic signals that trigger calcium release from intracellular stores (e.g., thapsigargin) activate Bad by inducing its dephosphorylation via the calcium-dependent protein phosphatase, calcineurin.69 Bad dephosphorylation and activation are also induced by growth factor withdrawal. Activated Bad translocates to mitochondria, inducing mitochondrial dysfunction and cytochrome c release.
Although in each of these examples activation of a proapoptotic Bcl-2 family member triggers a mitochondria-dependent apoptotic pathway, other proapoptotic family members may trigger cell death through mitochondria/cytochrome c-independent mechanisms. For example, the BH3 peptide of Bak appears to activate effector caspases through a cytochrome c-independent mechanism.70
Based on evidence that proapoptotic Bcl-2 family members mediate apoptosis induction by a broad range of apoptotic signals, it is intriguing to speculate that corticosteroid-induced apoptosis may be mediated through activation of proapoptotic Bcl-2 family members. Based on findings in the Bid deficient knockout mouse, Bid is not necessary for induction of thymocyte apoptosis by dexamethasone.71 Also, findings in the Bax deficient knockout mouse indicate that Bax is not required for dexamethasone-induced thymocyte apoptosis.72 However, dexamethasone treatment shifts the subcellular location of Bax from a soluble to a membrane-bound form.73 Thus, Bax may play a role in corticosteroid-induced apoptosis.
Recent findings have focused attention on the potential roles of Bak, which structurally resembles Bax, and two BH3 domain only family members, Bad and Bim, in corticosteroid-induced apoptosis. In human lymphoid leukemia cell lines, dexamethasone treatment, as well as staurosporine and etoposide treatment, induced a conformational change in Bak that preceded changes in mitochondrial membrane potential and was not inhibited by Z-VAD.fmk.74 In chicken thymocytes, differential display analysis uncovered a nucleotide sequence that was induced by dexamethasone and that demonstrated limited homology to Bad.75 Also, dexamethasone treatment induces elevated levels of Bax in thymocytes.76 Another BH3 domain-only member, Bim, has been implicated in corticosteroid-induced apoptosis by studies in a Bim knockout mouse showing that Bim deficiency inhibits the induction of apoptosis in thymocytes by dexamethasone.77
Thus, in the model in Figure 7, it is suggested that proapoptotic Bcl-2 family members, including Bim, Bak, Bax, or Bad, may trigger the caspase cascade. A critical question that remains to be solved is how one or more of these proapoptotic proteins are activated following corticosteroid treatment. In addition, it remains to be determined whether these proapoptotic proteins trigger APAF-1/caspase-9 activation through a mitochondria-dependent pathway involving cytochrome c release, or a mitochondria-independent pathway. As noted below, the current weight of evidence favors a mitochondria/cytochrome c-independent pathway to caspase activation in corticosteroid-induced apoptosis.

Proposed sequence of events in the execution phase of glucocorticoid-induced apoptosis. Proapoptotic members of the Bcl-2 family have been implicated in mediating glucocorticoid-induced apoptosis, which appears to proceed through either a caspase-9 dependent, caspase-3 independent pathway, or a caspase-9 independent, caspase-3 dependent pathway. Note that the sequence of caspase activation is different than that associated with Fas/TNF-induced apoptosis. Also, current evidence suggests that apoptosis induction in glucocorticoid-treated lymphoid cells is cytochrome c independent, and that mitochondrial dysfunction is a downstream event not necessary for the commitment to cell death
In summary, both antiapoptotic and proapoptotic members of the Bcl-2 family are involved in regulating corticosteroid-induced apoptosis. However, their site of action is yet to be determined. Although in many forms of apoptosis we have learned that Bcl-2 family members act primarily at the level of mitochondria, in the case of corticosteroid-induced apoptosis it appears that Bcl-2 family members may regulate degradation of cell cycle factors and transcription factors by the proteasome.
Changes in calcium and potassium homeostasis
Calcium has been implicated as a mediator of corticosteroid-induced thymocyte apoptosis for a number of years, yet the specific role of calcium in apoptosis has not been defined.78,79 Basically, two types of evidence support a role of calcium in corticosteroid-induced apoptosis: first, inhibitors of calcium-activated proteases, or calpains, have been reported to inhibit dexamethasone-induced apoptosis in thymocytes;80 second, alterations of intracellular calcium homeostasis have been detected in lymphoid cells undergoing corticosteroid-induced apoptosis.60,78,79,81,82
The major intracellular reservoir of calcium in non-muscle cells is the endoplasmic reticulum (ER). Corticosteroid treatment is associated with a decline in the calcium concentration within the ER lumen, which contributes to a decrease in total cellular calcium.60,81,82 Moreover, recent findings have implicated the inositol trisphosphate receptor (IP3R) in corticosteroid-induced apoptosis. The IP3R is an IP3-gated calcium release channel in the ER membrane. Lymphocytes undergoing apoptosis in response to dexamethasone were found to have increased levels of IP3R expression, and antisense-mediated repression of IP3R expression was reported to inhibit dexamethasone-induced apoptosis.83 Also, IP3R-deficient T cells are resistant to apoptosis induction by dexamethasone.84 Therefore, these findings suggest that calcium release from the ER, via the IP3R, produces cytoplasmic calcium elevation and ER calcium pool depletion that triggers downstream effector pathways of apoptosis.
An alternative theory is that dexamethasone treatment induces expression of a plasma membrane calcium channel, the P2X receptor, resulting in an elevation of cytosolic calcium.85 However, recent findings suggest that P2X receptor expression is not altered by dexamethasone treatment, that the P2X receptor agonist, ATP, does not induce apoptosis in thymocytes, and that P2X receptor antagonists do not block corticosteroid-induced apoptosis.86
The potassium ion is also implicated in corticosteroid-induced apoptosis. The potassium ion contributes to maintenance of cell volume, and volume loss is a characteristic feature of apoptosis in thymocytes.87 Two phases of volume loss have been identified in CEM human T cell leukemia cells treated with dexamethasone: the first is a reversible phase, associated with net loss of potassium ions, while the second phase coincides with chromatin condensation.88 Potassium efflux enhances apoptosis in thymocytes.89 Potassium at normal intracellular levels inhibits both apoptotic DNA fragmentation and caspase-3 activation.82 Recent findings indicate that thymocyte apoptosis is accompanied by gross perturbations of plasma membrane potential related to loss of cytosolic potassium.90 Furthermore, an inhibitor of plasma membrane potassium channels (tetrapentylammonium) was found to be an effective inhibitor of dexamethasone-induced apoptosis. This inhibitor prevented dissipation of mitochondrial membrane potential, loss of cytosolic potassium, phosphatidylserine exposure on the cell surface, and chromatin condensation, as well as caspase and endonuclease activation.90 These findings suggest that potassium channels contribute significantly to the regulation of some but not all pathways leading to thymocyte apoptosis.
Sphingomyelin hydrolysis and ceramide
Ceramide is produced under a variety of conditions that lead to apoptosis.91,92 Corticosteroid-induced apoptosis is no exception, as dexamethasone treatment of normal thymocytes causes a dose-dependent increase in the concentration of endogenous ceramide.93 The induction of ceramide appears to be secondary to induction of both acidic and neutral sphingmyelinases. Although recent evidence has pointed to phospholipid scrambling in the plasma membrane as the mechanism of sphingomyelin activation during apoptosis,94 studies in corticosteroid-treated thymocytes suggest that sphingomyelinase activation and ceramide generation proceed by a different mechanism.93
In thymocytes, dexamethasone treatment rapidly induces diacylglycerol through a protein kinase C and G-protein-dependent phosphatidylinositol-specific phospholipase C (PI-PLC). This event appears necessary for acid sphingomyelinase activation.93 The increase in ceramide was quite rapid, being detected within 5 min of hormone addition and reaching a maximum level at 15 min after dexamethasone treatment. Moreover, monesin, an endolysosomotropic agent that alkalinizes lysosomes inhibited the induction of sphingomyelinase activity, ceramide generation and apoptosis in dexamethasone treated thymocytes.93
These metabolic changes are inhibited by the glucocorticoid receptor antagonist RU486 and are therefore dependent upon the glucocorticoid receptor. In contrast to the recent concept that ceramide generation is secondary to caspase activation,92 in dexamethasone-treated thymocytes caspase-3 activation was blocked by inhibitors of both PI-PLC and acid sphingomyelinase.93 These findings argue that ceramide generation may occur upstream of caspase activation in dexamethasone-treated thymocytes.
A provocative observation was that cycloheximide and actinomycin D inhibited dexamethasone-induced apoptosis, but not dexamethasone-induced diacylglycerol and ceramide generation, and induction of caspase-3 activity.93 Based on these findings, the authors suggest that transcription and protein synthesis are required for dexamethasone-induced apoptosis and these events are downstream in the pathway after diacylglycerol and ceramide generation and caspase activation.
Could it be that there is more than one pathway to apoptosis in corticosteroid treated lymphocytes, and that the pathway involving ceramide-mediated caspase-3 activation is one of these pathways? This concept is at least consistent with evidence, summarized below, for a caspase-3-dependent cell death pathway and a caspase-3-independent pathway in corticosteroid-treated lymphocytes.
Execution stage
Caspases
Apoptosis is mediated by caspases, a family of proteases that cleave substrates at aspartate residues. Caspases are grouped into the initiator caspases and effector caspases. For example, in Fas-induced apoptosis the initiator caspase, caspase-8, is activated by recruitment to a complex composed of the cytoplasmic portion of the Fas receptor in conjunction with adapter molecules that contain death domains. Caspase-8 activates effector caspases (e.g., caspase-3) by one of two routes: a mitochondria-dependent pathway and a mitochondria-independent pathway.33 In the former, caspase-8-mediated cleavage of Bid, a proapoptotic member of the Bcl-2 family, converts Bid to an active form that triggers cytochrome c release from mitochondria. Cytochrome c binds APAF1, thereby activating another initiator caspase, caspase-9, which in turn activates effector caspases (e.g., caspase-3). In the mitochondria-independent pathway, caspase-8 is initially activated by the same mechanism, and then directly cleaves and activates caspase-3, bypassing mitochondria and cytochrome c release.
These well-established apoptotic pathways provide a conceptual model against which mechanistic features of corticosteroid-induced apoptosis can be compared. A recent mutational analysis of corticosteroid-resistant clones of the WEHI7.2 mouse lymphoma line provided genetic evidence for shared components in the dexamethasone- and Fas-mediated apoptotic pathways.95 At a biochemical level, certain elements of the Fas pathways appear to function in corticosteroid-induced apoptosis, although a number of gaps in our knowledge blur the picture considerably.
Perhaps the most certain conclusions regarding the role of specific caspases in corticosteroid-induced apoptosis are provided by knockout mouse experiments. These experiments indicate that both APAF1 and caspase-9 are essential for corticosteroid-induced apoptosis. Thymocytes from knockout mice deficient in either APAF196 or caspase-997,98 are resistant to corticosteroid-induced apoptosis. On the other hand, caspase-1 deficiency99 and caspase-3 deficiency100 do not prevent corticosteroid-induced apoptosis. This indicates that caspase-3 is not required for corticosteroid-induced apoptosis.
Although knockout mouse studies define critical elements of corticosteroid-induced apoptotic pathway, they also reveal the complexity of the pathway. For example, in caspase-9 -/- thymocytes treated with dexamethasone, pro-caspases-2, -3, -7 and -8 underwent processing to a limited extent, and cleavage of PARP (poly-ADP-ribose polymerase) and Rb (the retinoblastoma protein) was observed.97 Thus, there appear to be two alternative pathways to corticosteroid-induced apoptosis. One pathway, which appears to predominate in thymocytes, is caspase-9 dependent and caspase-3 independent, while the other is caspase-9 independent and caspase-3 dependent.
Also, knockout studies have revealed an important difference between Fas-induced apoptosis and corticosteroid-induced apoptosis. Proteolytic activation of procaspase-8 was markedly reduced in Apaf-1 -/- thymocytes treated with dexamethasone, whereas procaspase-8 processing in response to Fas was not diminished. These findings suggest that, in the case of corticosteroid-treated thymocytes, procaspase-8 activation is downstream of Apaf-1-mediated caspase-9 activation.
Studies using caspase inhibitors complement the findings in knockout mice. Corticosteroid-induced apoptosis in both thymocytes and lymphoma cells is inhibited by the broad spectrum caspase inhibitor, Z-VAD.fmk.61,101,102,103,104 Also, the baculovirus p35 protein, which inhibits caspase activity, blocks corticosteroid-induced apoptosis.105
Although there is good agreement regarding the inhibition of corticosteroid-induced apoptosis by broad spectrum caspase inhibitors, there is disagreement regarding the effect of the more specific caspase-3 inhibitor, Z-DEVD.fmk. One report has suggested that corticosteroid-induced apoptosis can be inhibited by the caspase-3 inhibitor, Z-DEVD.fmk,61 whereas other reports have failed to detect inhibition of corticosteroid-induced apoptosis by Z-DEVD.fmk.104,106 Although such discrepancies may be due to differences in experimental methods, they could also arise if a caspase-3-dependent pathway predominates in certain lymphoid subtypes, whereas a caspase-3-independent pathway predominates in other lymphoid subtypes.
Moreover, reports differ with regard to results of caspase-3 activity assays following corticosteroid treatment, with one report detecting caspase-3 activation in corticosteroid-treated cells107 and another report indicating that caspase-6, rather than caspase-3, is activated in corticosteroid-induced apoptosis.106 Again, these findings are consistent with the results of knockout mouse studies and suggest that there may be more than one pathway to corticosteroid-induced apoptosis, one in which caspase-3 predominates and one in which caspase-6 predominates.
The relative importance of caspase-6 over caspase-3 in corticosteroid-induced apoptosis is highlighted by a recent study by Komoriya et al108 using cell-permeable fluorogenic caspase substrates, with the goal of ordering the sequence of events in the caspase cascade in vivo. The findings of this study demonstrate that the sequence of caspase activation is different in corticosteroid-induced apoptosis compared to Fas-induced apoptosis. After dexamethasone treatment there is a pattern of sequential caspase activation with time, as expected for the caspase cascade proposed from biochemical studies, with caspase-9, followed by caspase-1, caspase-6, caspase-8, and caspases-3 and -7. The order of caspase activation for anti-Fas-treated thymocytes was less clear-cut, but distinct from dexamethasone-treated thymocytes with caspase-8 initially activated and caspase-9 activated at a latter stage.
It is important to keep in perspective that caspase activation and caspase-mediated protein cleavage are components of the execution stage of programmed cell death. In the case of corticosteroid-induced apoptosis critical events preceding caspase activation determine whether the cell is to live or to die. This point is emphasized by work showing that caspase inhibitors block proteolysis of endogenous substrates and reduce nuclear condensation, but do not alter dexamethasone-induced changes in clonogenicity. Therefore, it appears that commitment to cell death precedes key biochemical and morphological markers of apoptosis by several hours, and that separate regulators govern cellular commitment to clonogenic death and the subsequent execution phase characterized as apoptosis.65
In summary, based on the constellation of findings from knockout mice, inhibitor studies and caspase assays, the prevailing pathway to corticosteroid induced apoptosis appears to be caspase-9-dependent and caspase-3-independent (Figure 7). This pathway involves APAF-1-mediated activation of caspase-9 (the initiator caspase), followed by activation of caspases-1 and -6 (the effector caspases). An alternative pathway that is caspase-9-independent and caspase-3-dependent may also exist, but is less well defined. A critical question that remains to be answered is how corticosteroid treatment triggers activation of the caspase cascade and which proapoptotic Bcl-2 family members (e.g., Bim, Bak, Bax, Bad) are involved in this process.
Mitochondrial dysfunction
Dexamethasone treatment induces loss of mitochondrial membrane potential in thymocytes, followed by increased superoxide generation.109,110 Also, dexamethasone treatment induces loss of mitochondrial membrane potential in T-cell hybridoma cells.62 However, it appears that mitochondrial dysfunction occurs downstream of caspase activation in corticosteroid-induced apoptosis.
Perhaps the most convincing evidence for this conclusion comes from gene knockout studies. Caspase-9 deficiency in the caspase-9 knockout mouse prevented loss of mitochondrial membrane potential following dexamethasone treatment.97 Less certain are studies of the effect of the broad spectrum caspase inhibitor, Z-VAD.fmk, on changes in mitochondrial function in dexamethasone-treated cells. One study reported that Z-VAD.fmk prevented loss of mitochondrial membrane potential and production of reactive oxygen species,106 whereas another study reported that Z-VAD.fmk did not prevent dexamethasone-induced changes in mitochondrial membrane potential.65 Surprisingly, only one reported study has examined the role of cytochrome c release in corticosteroid-induced apoptosis. This study, employing dexamethasone-sensitive multiple myeloma lines, indicated that cytochrome c release was not detected in association with apoptosis induction by dexamethasone.111
In summary, the role of mitochondria in corticosteroid-induced apoptosis is uncertain, but based on the collective evidence available at the present time, mitochondrial dysfunction does not appear to be a central step in the initiation of the cell death pathway in corticosteroid-induced apoptosis. Although mitochondrial dysfunction is detected during the evolution of apoptosis in dexamethasone-treated cells, it appears to occur downstream of commitment to cell death, and may be induced by caspase activity (Figure 7). Thus, it will be important in future studies to look for a mitochondria-independent pathway to cell death in corticosteroid-treated cells.
Clinical actions of corticosteroids and corticosteroid resistance in leukemia and lymphoma
Corticosteroids are employed frequently in the treatment of both acute and chronic lymphocytic leukemia, and a variety of different lymphomas, due to their ability to induce apoptosis. Recent advances in understanding the mechanism of corticosteroid-induced apoptosis have been applied to better understand the role of corticosteroids in lymphoid malignancy.
Perhaps the most revealing are studies of corticosteroid treatment in acute lymphoblastic leukemia (ALL). A recent randomized trial, reviewed by Gaynon and Carrel,112 found that corticosteroids are clearly essential for optimal event free survival, and deferral of steroid treatment until the second month of therapy reduced survival. Also, the in vitro and in vivo response of leukemic cells to corticosteroids is highly predictive of outcome. At relapse, loss of in vitro sensitivity to corticosteroids is common and out of proportion to loss of sensitivity to other agents. Moreover, in a large group of children with newly diagnosed ALL, in vitro resistance to prednisone predicted a poor response to in vivo prednisone monotherapy and worse long term outcome.113
The mechanism of corticosteroid resistance in human leukemia and lymphoma cells is poorly understood. A recent report identifying a somatic mutation of the glucocorticoid receptor gene in leukemic blasts from a patient with acute lymphoblastic leukemia, and a cell line derived from the same patient, suggests that there may be value in searching for additional receptor mutations and testing their association with corticosteroid resistance.114 In view of recent findings, described above, indicating that Ras activity can repress corticosteroid-induced apoptosis in vitro,55 it is interesting to recall an earlier clinical study showing that activating N-Ras point mutations are detected in some cases of dexamethasone-resistant acute lymphoblastic leukemia.115
The possibility that expression levels of Bcl-2 family members may be of prognostic significance has also been tested in recent clinical trials. In cell lines, the Bax/Bcl-2 ratio correlated with sensitivity to dexamethasone.116 In bone marrow derived blasts, the Bcl-2 and Bax levels were highly variable, although the Bax/Bcl-2 ratio appeared to be more important than the Bcl-2 level as a predictor of drug-induced apoptosis.
In a clinical trial of childhood ALL reported by Salomons et al,117 blasts from patients with newly diagnosed childhood ALL were examined for levels of Bcl-2 protein family expression. Expression levels of apoptosis inducers (Bad, Bak) and inhibitors (Bcl-xL, Mcl-1) were highly variable in blasts from the 78 children with newly diagnosed ALL tested. Protein expression levels of the Bcl-2 family were not found to correlate with in vitro resistance to drugs, including prednisone. Moreover, neither blast reduction following one week of prednisone monotherapy, nor long term disease-free survival showed a correlation with Bcl-2 family protein expression.
A different conclusion was reached in a study of newly diagnosed and relapsed ALL reported by Haarman et al.118 In this study, in vitro assays of drug sensitivity were performed at the time of diagnosis and at the time of relapse. At time of initial diagnosis, Bcl-2 expression was not associated with increased in vitro drug resistance. But at the time of relapse, increased expression of Bcl-2 correlated with increased resistance to corticosteroid-induced cell death.
In summary, basic understanding of the mechanism of corticosteroid-induced apoptosis is presently being applied to better understand and predict that action of corticosteroids in the clinical setting. A more in depth understanding of the fundamental cell death pathway, and its regulation, will no doubt lead to even greater insight into mechanisms of corticosteroid resistance in the clinical setting, and perhaps to novel modes of therapeutic intervention.
Summary and speculation
The mechanism of corticosteroid-induced apoptosis is only partially understood, and there are many questions to be answered. First, although there is considerable evidence that receptor-mediated regulation of gene transactivation is important, the identity of specific genes or gene networks involved in initiating the cell death process is unknown. At the other end of the spectrum, there is considerable knowledge about which caspases are involved in cell death execution, and the sequence of their activation. However, the mechanism that initiates the caspase cascade is unknown. Although certain proapoptotic members of the Bcl-2 family have been implicated, how they actually trigger caspase activation is uncertain. Although it is tempting to postulate that cytochrome c release from mitochondria is the triggering event, as it is in many forms of apoptosis, there is surprisingly little information about cytochrome c release in corticosteroid-induced apoptosis. The only report suggests that cytochrome c is not released from mitochondria during the course of corticosteroid-mediated apoptosis. Could it be that corticosteroids activate caspases through an APAF-1/caspase-9 dependent, but mitochondria/cytochrome c-independent mechanism?
Between the initiation stage of cell death and the execution stage of cell death lies a mystery. How do transcriptional changes mediated by the corticosteroid receptor lead the cell down a path to death? This review has summarized a number of events that occur in lymphoid cells following exposure to a corticosteroid hormone. Each event, when viewed in isolation, could provide basis for a hypothesis about what governs the cell death decision. However, upon closer examination at least one common theme emerges from the reported observations: corticosteroids appear to oppose the action of growth factors.
This concept is illustrated in Figure 8, and is described as the ‘Yin and Yang’ of corticosteroid-induced apoptosis. It may be the balance between the prosurvival action of growth factors and the proapoptotic action of corticosteroids that ultimately determines a cell's fate. Growth factors increase the activity of a number of transcription factors that mediate expression of genes involved in cell proliferation. Corticosteroids appear to decrease the activity of these same transcription factors. All of these transcription factors are targets of the multicatalytic proteasome. Hence, corticosteroid treatment of susceptible lymphoid cells may mimic growth factor withdrawal by targeting critical transcription factors for accelerated degradation by the multicatalytic proteasome. The role of the proteasome in corticosteroid-induced apoptosis is not limited to transcription factor targets. An enzyme involved in polyamine synthesis, ornithine decarboxylase, is also degraded by the proteasome in corticosteroid-treated cells, thus counterbalancing the induction of this enzyme by IL-3.

The ‘Yin and Yang’ of glucocorticoid-induced apoptosis. Cytokines and Bcl-2 favor the expression of a series of factors important for cell proliferation and cell survival, whereas glucocorticoids have the opposite effect. It appears that the cell death decision is based on the balance of these opposing actions
Although the preceding arguments suggest a mechanism by which corticosteroids may counterbalance the proliferative effects of growth factors, they do not readily explain how the decision to die is translated into caspase activation and cell death execution. One clue is that proteins that function to inhibit caspases (i.e., c-IAP1, XIAP) are degraded by the proteasome during corticosteroid-induced apoptosis. It is likely that other regulatory molecules that serve to keep either proapoptotic Bcl-2 family members or caspases in check will also be found to be targets of the proteasome during corticosteroid-induced apoptosis. Thus, although somewhat speculative at this point, it seems possible that corticosteroid-induced apoptosis is mediated by targeting a variety of prosurvival factors for degradation by the multicatalytic proteasome. Interestingly, the antiapoptotic protein Bcl-2 has been reported to stabilize several of the proteasome targets implicated in corticosteroid-induced apoptosis. Thus, it is possible that proteasome-mediated degradation of prosurvival factors may be a central, Bcl-2 regulated step in corticosteroid-induced apoptosis.
Abbreviations
- IL:
-
interleukin
- AP-1:
-
activating protein-1
- CTLL:
-
cytotoxic T lymphocytic leukemia
- NFAT:
-
nuclear factor of activated T lymphocytes
References
Wyllie AH . 1980 Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonuclease activation Nature 284: 555–556
Helmberg A, Auphan N, Caelles C, Karin M . 1995 Glucocorticoid-induced apoptosis of human leukemic cells is caused by the repressive function of the glucocorticoid receptor EMBO J. 14: 452–460
Karin M, Liu Z-G, Zandi E . 1997 AP-1 function and regulation Curr. Opin. Cell. Biol. 9: 240–246
Wisdom R . 1999 AP-1: One switch for many signals J. Exp. Med. 253: 180–185
Walker PR, Kwast-Welfeld J, Gourdeau H, Leblanc J, Neugebauer W, Sikorska M . 1993 Relationship between apoptosis and the cell cycle in lymphocytes: roles of protein kinase C, tyrosine phosphorylation, and AP1 Exp. Cell. Res. 207: 142–151
Wisniewska M, Stanczyk M, Grzelakowska-Sztabert B, Kaminska B . 1997 Nuclear factor of activated T cells (NFAT) is a possible target for dexamethasone in thymocyte apoptosis Cell. Biol. Int. 21: 127–132
Thompson EB . 1998 The many roles of c-Myc in apoptosis Annu. Rev. Physiol. 60: 575–600
Chapman MS, Askew DJ, Kuscuoglu U, Miesfeld RL . 1996 Transcriptional control of steroid-regulated apoptosis in murine thymoma cells Mol. Endocrinol. 10: 967–978
Ramdas J, Harmon JM . 1998 Glucocorticoid-induced apoptosis and regulation of NF-kappaB activity in human leukemic T cells Endocrinology 139: 3813–3821
Reichardt HM, Kaestner KH, Tuckermann J, Katz O, Wessely O, Bock R, Gass P, Schmid W, Herrlich P, Angel P, Schutz G . 1998 DNA binding of the glucocorticoid receptor is not essential for survival Cell 93: 531–541
Kullmann M, Schneikert J, Moll J, Heck S, Zeiner M, Gehring U et al. 1998 RAP46 is a negative regulator of glucocorticoid receptor action and hormone-induced apoptosis J. Biol. Chem. 273: 14620–14625
Hulkko SM, Wakui H, Zilliacus J . 2000 The pro-apoptotic protein death-associated protein 3 (DAP3) interacts with the glucocorticoid receptor and affects the receptor function Biochem. J. 349: 885–893
Potter M, Bernstein A, Lee JM . 1998 The wst gene regulates multiple forms of thymocyte apoptosis Cell. Immunol. 188: 111–117
Ramdas J, Liu W, Harmon JM . 1999 Glucocorticoid-induced cell death requires autoinduction of glucocorticoid receptor expression in human leukemic T cells Cancer Res. 59: 1378–1385
Diamond MI, Miner JN, Yoshinaga SK, Yamamoto KR . 1990 Transcription factor interactions: selectors of positive or negative regulation from a single DNA element Science 249: 1266–1272
Miner JN, Yamamoto KR . 1992 The basic region of AP-1 specifies glucocorticoid receptor activity at a composite response element Genes & Development 6: 2491–2501
Goldberg AL, Rock KL . 1992 Proteolysis, proteasomes and antigen presentation Nature 357: 375–379
Rechsteiner M, Hoffman L, Dubiel W . 1993 The multicatalytic and 26 S proteases J. Biol. Chem. 268: 6065–6068
Ciechanover A . 1994 The ubiquitin-proteasome proteolytic pathway Cell 79: 13–21
Schwartz LM, Myer A, Kosz L, Engelstein M, Maier C . 1990 Activation of polyubiquitin gene expression during developmentally programmed cell death Neuron 5: 411–419
Jones MEE, Haire MF, Kloetzel PM, Mykles DL, Schwartz LM . 1995 Changes in the structure and function of the multicatalytic proteinase (proteasome) during programmed cell death in the intersegmental muscles of the hawkmoth, Manduca sexta Develop. Biol. 169: 436–447
Dawson SP, Arnold JE, Mayer NJ, Reynolds SE, Billett MA, Gordon C, Colleaux L, Kloetzel PM, Tanaka K, Mayer RJ . 1995 Developmental changes of the 26S proteasome in abdominal intersegmental muscles of Manduca sexta during programmed cell death J. Biol. Chem. 270: 1850–1858
Lesche R, Peetz A, van der Hoeven F, Ruther U . 1997 Ft1, a novel gene related to ubiquitin-conjugating enzymes, is deleted in the Fused toes mouse mutation Mamm. Genome 8: 879–883
Grimm LM, Goldberg AL, Poirier GG, Schwartz LM, Osborne BA . 1996 Proteasomes play an essential role in thymocyte apoptosis EMBO J. 15: 3835–3844
Beyette J, Mason GG, Murray RZ, Cohen GM, Rivett AJ . 1998 Proteasome activities decrease during dexamethasone-induced apoptosis of thymocytes Biochem. J. 332: 315–320
Hirsch T, Dallaporta B, Zamzami N, Susin SA, Ravagnan L, Marzo I, Brenner C, Kroemer G . 1998 Proteasome activation occurs at an early, premitochondrial step of thymocyte apoptosis J. Immunol. 161: 35–40
Dallaporta B, Pablo M, Maisse C, Daugas E, Loeffler M, Zamzami N, Kroemer G . 2000 Proteasome activation as a critical event of thymocyte apoptosis Cell Death Differ. 7: 368–373
He H, Qi X-M, Grossman J, Distelhorst CW . 1998 c-Fos degradation by the proteasome: An early, Bcl-2 regulated step in apoptosis J. Biol. Chem. 273: 25015–25019
Ivanov VN, Nikolic-Zugic J . 1998 Biochemical and kinetic characterization of the glucocorticoid-induced apoptosis of immature CD4+CD8+ thymocytes Int. Immunol. 10: 1807–1817
Grassilli E, Benatti F, Dansi P, Giammarioli AM, Malorni W, Franceschi C, Desiderio MA . 1998 Inhibition of proteasome function prevents thymocyte apoptosis: involvement of ornithine decarboxylase Biochem. Biophys. Res. Commun. 250: 293–297
Gil-Gomez G, Berns A, Brady HJ . 1998 A link between cell cycle and cell death: Bax and Bcl-2 modulate Cdk2 activation during thymocyte apoptosis EMBO J. 17: 7209–7218
Yang Y, Fang S, Jensen JP, Weissman AM, Ashwell JD . 2000 Ubiquitin protein ligase activity of IAPs and their degradation in proteasomes in response to apoptotic stimuli Science 288: 874–877
Green DR . 2000 Apoptotic pathways: paper wraps stone blunts scissors Cell 102: 1–4
Di Croce L, Okret S, Kersten S, Gustafsson J, Parker M, Wahli W, Beato MS . 1999 Steroid and nuclear receptors. Villefranche-sur-Mer, France, May 25–27, 1999 EMBO J. 18: 6201–6210
Gao Y, Kinoshita Y, Hato F, Tominaga K, Tsuji Y . 1996 Suppression of glucocorticoid-induced thymocyte apoptosis by co-culture with thymic epithelial cells Cell. Mol. Biol. Noisy-le-grand 42: 227–234
Ferlin-Bezombes M, Jourdan M, Liautard J, Brochier J, Rossi JF, Klein B . 1998 IFN-alpha is a survival factor for human myeloma cells and reduces dexamethasone-induced apoptosis J. Immunol. 161: 2692–2699
Chauhan D, Pandey P, Ogata A, Teoh G, Treon S, Urashima M, Kharbanda S, Anderson KC . 1997 Dexamethasone induces apoptosis of multiple myeloma cells in a JNK/SAP kinase independent mechanism Oncogene 15: 837–843
Xu FH, Sharma S, Gardner A, Tu Y, Raitano A, Sawyers C, Lichtenstein A . 1998 Interleukin-6-induced inhibition of multiple myeloma cell apoptosis: support for the hypothesis that protection is mediated via inhibition of the JNK/SAPK pathway Blood 92: 241–251
Van Snick J, Houssiau F, Proost P, Van Damme J, Renauld JC . 1996 I-309/T cell activation gene-3 chemokine protects murine T cell lymphomas against dexamethasone-induced apoptosis J. Immunol. 157: 2570–2576
Bauer JH, Liu KD, You Y, Lai SY, Goldsmith MA . 1998 Heteromerization of the gammac chain with the interleukin-9 receptor alpha subunit leads to STAT activation and prevention of apoptosis J. Biol. Chem. 273: 9255–9260
Bulfone-Paus S, Ungureanu D, Pohl T, Lindner G, Paus R, Ruckert R, Krause H, Kunzendorf U . 1997 Interleukin-15 protects from lethal apoptosis in vivo Nat. Med. 3: 1124–1128
Xu F, Gardner A, Tu Y, Michl P, Prager D, Lichtenstein A . 1997 Multiple myeloma cells are protected against dexamethasone-induced apoptosis by insulin-like growth factors Br. J. Haematol. 97: 429–440
Guizani L, Perrin-Wolff M, Breard J, Binetruy B, Bertoglio J . 1996 Mechanisms in interleukin-2 protection against glucocorticoid-induced apoptosis: regulation of AP-1 and glucocorticoid receptor transcriptional activities J. Interferon Cytokine Res. 16: 601–609
Ogawa M, Nishiura T, Oritani K, Yoshida H, Yoshimura M, Okajima Y, Ishikawa J, Hashimoto I, Matsumura I, Tomiyama Y, Matsuzawa Y . 2000 Cytokines prevent dexamethasone-induced apoptosis via the activation of mitogen-activated protein kinase and phosphatidylinositol 3-kinase pathways in a new multiple myeloma cell line Cancer Res. 60: 4262–4269
Wang W, Wykrzykowska J, Johnson T, Sen R, Sen J . 1999 A NF-kappa B/c-myc-dependent survival pathway is targeted by corticosteroids in immature thymocytes J. Immunol. 162: 314–322
Xie H, Seward RJ, Huber BT . 1997 Cytokine rescue from glucocorticoid induced apoptosis in T cells is mediated through inhibition of IkappaBalpha Mol. Immunol. 34: 987–994
Auphan N, DiDonato JA, Rosette C, Helmberg A, Karin M . 1995 Immunosuppression of I kappa B synthesis [see comments] Science 270: 286–290
Brostjan C, Anrather J, Csizmadia V, Stroka D, Soares M, Bach FH et al. 1996 Glucocorticoid-mediated repression of NFkB activity in endothelial cells does not involve induction of IkBa synthesis J. Biol. Chem. 271: 19612–19616
De Bosscher K, Schmitz ML, Vanden Berghe W, Plaisance S, Fiers W, Haegeman G . 1997 Glucocorticoid-mediated repression of nuclear factor-kB-dependent transcription involves direct interference with transactivation Proc. Natl. Acad. Sci. USA 94: 13504–13509
Heck S, Bender K, Kullmann M, Gottlicher M, Herrlich P, Cato AC . 1997 I kappaB alpha-independent downregulation of NF-kappaB activity by glucocorticoid receptor EMBO J. 16: 4698–4707
Adcock IM, Nasuhara Y, Stevens DA, Barnes PJ . 1999 Ligand-induced differentiation of glucocorticoid receptor (GR) trans-repression and transactivation: preferential targeting of NF-kappaB and lack of I-kappaB involvement Br. J. Pharmacol. 127: 1003–1011
Asada A, Zhao Y, Kondo S, Iwata M . 1998 Induction of thymocyte apoptosis by Ca2+-independent protein kinase C (nPKC) activation and its regulation by calcineurin activation J. Biol. Chem. 273: 28392–28398
Stefanelli C, Stanic I, Bonavita F, Flamigni F, Pignatti C, Guarnieri C, Caldarera CM . 1998 Inhibition of glucocorticoid-induced apoptosis with 5-aminoimidazole-4-carboxamide ribonucleoside, a cell-permeable activator of AMP-activated protein kinase Biochem. Biophys. Res. Commun. 243: 821–826
Walach D, Varfolomeev E, Malinin N, Goltsev Y, Kovalenko A, Boldin M . 1999 Tumor necrosis factor receptor and Fas signaling mechanisms Annu. Rev. Immunol. 17: 331–367
Jamieson CA, Yamamoto KR . 2000 Crosstalk pathway for inhibition of glucocorticoid-induced apoptosis by T cell receptor signaling [In Process Citation] Proc. Natl. Acad. Sci. USA 97: 7319–7324
Sears R, Leone G, DeGregori J, Nevins JR . 1999 Ras enhances Myc protein stability Mol. Cell 3: 169–179
Wakui H, Wright AP, Gustafsson J, Zilliacus J . 1997 Interaction of the ligand-activated glucocorticoid receptor with the 14-3-3 eta protein J. Biol. Chem. 272: 8153–8156
Widen C, Zilliacus J, Gustafsson JA, Wikstrom AC . 2000 Glucocorticoid receptor Interaction with 14-3-3 and Raf-1, a proposed mechanism for crosstalk of two signal transduction pathways J. Biol. Chem
Veis DJ, Sorenson CM, Shutter JR, Korsmeyer SJ . 1993 Bcl-2-deficient mice demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair Cell 75: 229–240
Bian X, Hughes FM, Huang Y, Cidlowski JA, Putney JW . 1997 Roles of cytoplasmic Ca2+ and intracellular Ca2+ stores in induction and suppression of apoptosis in S49 cells Am. J. Physiol. 272 (Cell Physiol. 41): C1241–C1249
McColl KS, He H, Zhong H, Whitacre CM, Berger NA, Distelhorst CW . 1998 Apoptosis induction by the glucocorticoid hormone dexamethasone and the calcium-ATPase inhibitor thapsigargin involves Bcl-2 regulated caspase activation Mol. Cell. Endocrin. 139: 229–238
Camilleri-Broet S, Vanderwerff H, Caldwell E, Hockenbery D . 1998 Distinct alterations in mitochondrial mass and function characterize different models of apoptosis Exp. Cell. Res. 239: 277–292
Gross A, McDonnell JM, Korsmeyer SJ . 1999 BCL-2 family members and the mitochondria in apoptosis Genes Dev. 13: 1899–1911
Kroemer G, Reed JC . 2000 Mitochondrial control of cell death Nature Medicine 6: 513–519
Brunet CL, Gunby RH, Benson RS, Hickman JA, Watson AJ, Brady G . 1998 Commitment to cell death measured by loss of clonogenicity is separable from the appearance of apoptotic markers Cell Death Differ. 5: 107–115
Hakem A, Sasaki T, Kozieradzki I, Penninger JM . 1999 The cyclin-dependent kinase Cdk2 regulates thymocyte apoptosis J. Exp. Med. 189: 957–967
Feinman R, Koury J, Thames M, Barlogie B, Epstein J, Siegel DS . 1999 Role of NF-kappaB in the rescue of multiple myeloma cells from glucocorticoid-induced apoptosis by bcl-2 Blood 93: 3044–3052
Budijardjo I, Oliver H, Lutter M, Luo X, Wang X . 1999 Biochemical pathways of caspase activation during apoptosis Annu. Rev. Cell. Dev. Biol. 15: 269–290
Wang HG, Pathan N, Ethell IM, Krajewski S, Yamaguchi Y, Shibasaki F . 1999 Ca2+-induced apoptosis through calcineurin dephosphorylation of BAD Science 284: 339–343
Holinger EP, Chittenden T, Lutz RJ . 1999 Bak BH3 peptides antagonize Bcl-xL function and induce apoptosis through cytochrome c-independent activation of caspases J. Biol. Chem. 274: 13298–13304
Yin XM, Wang K, Gross A, Zhao Y, Zinkel S, Klocke B, Roth KA, Korsmeyer SJ . 1999 Bid-deficient mice are resistant to Fas-induced hepatocellular apoptosis Nature 400: 886–891
Knudson CM, Tung KS, Tourtellotte WG, Brown GA, Korsmeyer SJ . 1995 Bax-deficient mice with lymphoid hyperplasia and male germ cell death Science 270: 96–99
Hsu YT, Wolter KG, Youle RJ . 1997 Cytosol-to-membrane redistribution of Bax and Bcl-X(L) during apoptosis Proc. Natl. Acad. Sci. USA 94: 3668–3672
Griffiths GJ, Dubrez L, Morgan CP, Jones NA, Whitehouse J, Corfe BM et al. 1999 Cell damage-induced conformational changes of the pro-apoptotic protein Bak in vivo precede the onset of apoptosis J. Cell. Biol. 144: 903–914
Thomson JM, Waldrip HW, Compton MM . 1997 Identification of a differential display product associated with apoptosis in chicken thymocytes Dev. Comp. Immunol. 21: 413–424
Mok CL, Gil-Gomez G, Williams O, Coles M, Taga S, Tolaini M, Norton T, Kioussis D, Brady HJ . 1999 Bad can act as a key regulator of T cell apoptosis and T cell development J. Exp. Med. 189: 575–586
Bouillet P, Metcalf D, Huang DC, Tarlinton DM, Kay TW, Kontgen F, Adams JM, Strasser A . 1999 Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity Science 286: 1735–1738
Orrenius S, McConkey DJ, Nicotera P . 1991 Role of calcium in toxic and programmed cell death Adv. Exp. Med. Biol. 283: 419–425
Distelhorst CW, Dubyak G . 1998 Role of calcium in glucocorticosteroid-induced apoptosis of thymocytes and lymphoma cells: Resurrection of old theories by new findings Blood 91: 731–734
Squier MK, Cohen JJ . 1997 Calpain, an upstream regulator of thymocyte apoptosis J. Immunol. 158: 3690–3697
Lam M, Dubyak G, Distelhorst CW . 1993 Effect of glucocorticosteroid treatment on intracellular calcium homeostasis in mouse lymphoma cells Mol. Endocrinol. 7: 686–693
Hughes FM Jr, Bortner CD, Purdy GD, Cidlowski JA . 1997 Intracellular K+ suppresses the activation of apoptosis in lymphocytes J. Biol. Chem. 272: 30567–30576
Khan AA, Soloski MJ, Sharp AH, Schilling G, Sabatini DM, Li S-H, Ross CA, Snyder SH . 1996 Lymphocyte apoptosis: Mediation by increased type 3 inositol 1,4,5-trisphosphate receptor Science 273: 503–507
Jayaraman T, Marks AR . 1997 T cells deficient in inositol 1,4,5-trisphosphate receptor are resistant to apoptosis Mol. Cell. Biol. 17: 3005–3012
Chvatchko Y, Valera S, Aubry J-P, Renno T, Buell G, Bonnefoy J-Y . 1996 The involvement of an ATP-gated ion channel, P2X1, in thymocyte apoptosis Immunity 5: 275–283
Jiang S, Kull B, Fredholm BB, Orrenius S . 1996 P2x purinoceptor is not important in thymocyte apoptosis Immunol. Lett. 49: 197–201
Bortner CD, Cidlowski JA . 2000 Volume regulation and ion transport during apoptosis [In Process Citation] Methods Enzymol. 322: 421–433
Benson RS, Heer S, Dive C, Watson AJ . 1996 Characterization of cell volume loss in CEM-C7A cells during dexamethasone-induced apoptosis Am. J. Physiol. 270: 1190–1203
Bortner C, Hughes FM Jr, Cidlowski JA . 1997 A primary role for K+ and Na+ efflux in the activation of apoptosis J. Biol. Chem. 272: 32436–32442
Dallaporta B, Marchetti P, de Pablo MA, Maisse C, Duc HT, Metivier D et al. 1999 Plasma membrane potential in thymocyte apoptosis J. Immunol. 162: 6534–6542
Kolesnick RN, Kronke M . 1998 Regulation of ceramide production and apoptosis Annu. Rev. Physiol. 60: 643–665
Green DR . 2000 Apoptosis and sphingomyelin hydrolysis. The flip side [comment] J. Cell. Biol. 150: F5–7
Cifone MG, Migliorati G, Parroni R, Marchetti C, Millimaggi D, Santoni A, Tepper AD, Ruurs P, Weidmer T, Sims PJ, Borst J, van Blitterswijk WJ . 1999 Dexamethasone-induced thymocyte apoptosis: apoptotic signal involves the sequential activation of phosphoinositide-specific phospholipase C, acidic sphingomyelinase, and caspases Blood 93: 2282–2296
Tepper AD, Ruurs P, Wiedmer T, Sims PJ, Borst J, van Blitterswijk WJ . 2000 Sphingomyelin hydrolysis to ceramide during the execution phase of apoptosis results from phospholipid scrambling and alters cell-surface morphology [see comments] J. Cell. Biol. 150: 155–164
Askew DJ, Kuscuoglu U, Brunner T, Green DR, Miesfeld RL . 1999 Characterization of Apt-cell lines exhibiting cross-resistance to glucocorticoid- and Fas-mediated apoptosis Cell Death Differ. 6: 796–804
Yoshida H, Kong YY, Yoshida R, Elia AJ, Hakem A, Hakem R, Duncan GS, Henderson JT, Woo M, Soengas MS, Elia A, de la Pompa JL, Kagi D, Khoo W, Potter J, Yoshida R, Kaufman SA, Lowe SW, Penninger JM, Mak TW . 1998 Apaf1 is required for mitochondrial pathways of apoptosis and brain development Cell 94: 739–750
Hakem R, Hakem A, Duncan GS, Henderson JT, Woo M, Soengas MS, Kuida K, Haydar TF, Kuan CY, Gu Y, Taya C, Karasuyama H, Su MS, Rakic P, Flavell RA . 1998 Differential requirement for caspase 9 in apoptotic pathways in vivo Cell 94: 339–352
Kuida K, Haydar TF, Kuan CY, Gu Y, Taya C, Karasuyama H, Li P, Allen H, Banerjee S, Seshadri T . 1998 Reduced apoptosis and cytochrome c-mediated caspase activation in mice lacking caspase 9 Cell 94: 325–337
Li P, Allen H, Banerjee S, Seshadri T . 1997 Characterization of mice deficient in interleukin-1 beta converting enzyme J. Cell. Biochem. 64: 27–32
Kuida K, Zheng TS, Na S, Kuan C-y, Yang D, Karasuyama H, Sarin A, Wu ML, Henkart PA . 1996 Decreased apoptosis in the brain and premature lethality in CP32-deficient mice Nature 384: 368–372
Sarin A, Wu ML, Henkart PA . 1996 Different interleukin-1 beta converting enzyme (ICE) family protease requirements for the apoptotic death of T lymphocytes triggered by diverse stimuli J. Exp. Med. 184: 2445–2450
Bellosillo B, Dalmau M, Colomer D, Gil J . 1997 Involvement of CED-3/ICE proteases in the apoptosis of B-chronic lymphocytic leukemia cells Blood 89: 3378–3384
Hughes FM Jr, Cidlowski JA . 1998 Glucocorticoid-induced thymocyte apoptosis: protease-dependent activation of cell shrinkage and DNA degradation J. Steroid Biochem. Mol. Biol. 65: 207–217
Weimann E, Baixeras E, Zamzami N, Kelly P . 1999 Prolactin blocks glucocorticoid induced cell death by inhibiting the disruption of the mitochondrial membrane Leuk Res. 23: 751–762
Robertson NM, Zangrilli J, Fernandes-Alnemri T, Friesen PD, Litwak G, Alnemri ES . 1997 Baculovirus p35 inhibits the glucocorticoid-mediated pathway of cell death Cancer Res. 57: 43–47
Miyashita T, Nagao K, Krajewski S, Salvesen GS, Reed JC, Inoue T, Alam A, Braun MY, Hartgers F, Lesage S, Cohen L, Hugo P, Denis F, Sekaly RP . 1998 Investigation of glucocorticoid-induced apoptotic pathway: processing of caspase-6 but not caspase-3 Cell Death Differ. 5: 1034–1041
Alam A, Braun MY, Hartgers F, Lesage S, Cohen L, Hugo P, Komoriya A, Packard BZ, Brown MJ, Wu M-L, Henkart PA . 1997 Specific activation of the cysteine protease CPP32 during the negative selection of T cells in the thymus J. Exp. Med. 186: 1503–1512
Komoriya A, Packard BZ, Brown MJ, Wu M-L, Henkart PA . 2000 Assessment of caspase activities in intact apoptotic thymocytes using cell-permeable fluorogenic caspase substrates J. Exp. Med. 191: 1819–1828
Castedo M, Macho A, Zamzami N, Hirsch T, Marchetti P, Uriel J, Susin SA, Zamzami N, Marchetti P, Macho A, Kroemer G . 1995 Mitochondrial perturbations define lymphocytes undergoing apoptotic depletion in vivo Eur. J. Immunol. 25: 3277–3284
Castedo M, Hirsch T, Susin ZA, Zamzami N, Marchetti P, Macho A, Chauhan D, Pandey P, Ogata A, Teoh G, Krett N, Halgren R, Rosen S, Kufe D, Anderson K . 1996 Sequential acquisition of mitochondrial and plasma membrane alterations during early lymphocyte apoptosis J. Immunol. 157: 512–521
Chauhan D, Pandey P, Ogata A, Teoh G, Krett N, Halgren R, Gaynon PS, Carrel AL . 1997 Cytochrome c-dependent and -independent induction of apoptosis in multiple myeloma cells J. Biol. Chem. 272: 29995–29997
Gaynon PS, Carrel AL . 1999 Glucocorticosteroid therapy in childhood acute lymphoblastic leukemia Adv. Exp. Med. Biol. 457: 593–605
Kaspers GJ, Pieters R, Van Zantwijk CH, Van Wering ER, Van Der Does-Van Den Berg A, Veerman AJ . 1998 Prednisolone resistance in childhood acute lymphoblastic leukemia: vitro-vivo correlations and cross-resistance to other drugs Blood 92: 259–266
Hillmann AG, Ramdas J, Multanen K, Norman MR, Harmon JM . 2000 Glucocorticoid receptor gene mutations in leukemic cells acquired in vitro and in vivo Cancer Res. 60: 2056–2062
Lubbert M, Mirro J, Miller CW, Kahan J, Isaac G, Salomons GS, Brady HJ, Verwijs-Janssen M, Van Den Berg JD, Hart AA, Van Den Berg H, Behrendt H, Hahlen K, Smets LA . 1990 N-ras gene point mutations in childhood acute lymphocytic leukemia correlate with a poor prognosis Blood 75: 1163–1169
Salomons GS, Brady HJ, Verwijs-Janssen M, Van Den Berg JD, Hart AA, Van Den Berg H . 1997 The Bax alpha:Bcl-2 ratio modulates the response to dexamethasone in leukaemic cells and is highly variable in childhood acute leukaemia Int. J. Cancer 71: 959–965
Salomons GS, Smets LA, Verwijs-Janssen M, Hart AA, Haarman EG, Kaspers GJ, Pieters R, van Zantwijk CH, Broekema GJ, Hahlen K, Veerman AJ . 1999 Bcl-2 family members in childhood acute lymphoblastic leukemia: relationships with features at presentation, in vitro and in vivo drug response and long-term clinical outcome Leukemia 13: 1574–1580
Haarman EG, Kaspers GJ, Pieters R, van Zantwijk CH, Broekema GJ, Hahlen K, Veerman AJ . 1999 BCL-2 expression in childhood leukemia versus spontaneous apoptosis, drug induced apoptosis, and in vitro drug resistance Adv. Exp. Med. Biol. 457: 325–333
Acknowledgements
This work was supported by NIH grant CA42755.
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by G Nunez
Rights and permissions
About this article
Cite this article
Distelhorst, C. Recent insights into the mechanism of glucocorticosteroid-induced apoptosis. Cell Death Differ 9, 6–19 (2002). https://doi.org/10.1038/sj.cdd.4400969
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.cdd.4400969
Keywords
This article is cited by
-
Chronic stress decreases ornithine decarboxylase expression and protects against 1,2-dimethylhydrazine-induced colon carcinogenesis
Molecular Biology Reports (2020)
-
Metabolic reprogramming of acute lymphoblastic leukemia cells in response to glucocorticoid treatment
Cell Death & Disease (2018)
-
Mechanism of inhibition of Shiga-toxigenic Escherichia coli SubAB cytotoxicity by steroids and diacylglycerol analogues
Cell Death Discovery (2018)
-
Subchronic Glucocorticoid Receptor Inhibition Rescues Early Episodic Memory and Synaptic Plasticity Deficits in a Mouse Model of Alzheimer’s Disease
Neuropsychopharmacology (2015)
-
Paclitaxel-induced transcriptional regulation of Fas signaling pathway is antagonized by dexamethasone
Breast Cancer Research and Treatment (2015)
