
Abstract
When radioactive materials are released into the environment due to nuclear power plant accidents, they may enter into the body, and exposing it to internal radiation for long periods of time. Although several agents have been developed that help excrete radioactive elements from the digestive tract, only one type of radioactive element can be removed using a single agent. Therefore, we considered the simultaneous removal of caesium (Cs) and strontium (Sr) by utilising the multiple metal removal mechanisms of probiotic bacteria. In this study, the Cs and Sr removal capacities of lactobacilli and bifidobacteria were investigated. Observation using an electron probe micro analyser suggested that Cs was accumulated within the bacterial cells. Since Sr was removed non metabolically, it is likely that it was removed by a mechanism different from that of Cs. The amount of Cs and Sr that the cells could simultaneously retain decreased when compared to that for each element alone, but some strains showed only a slight reduction in removal. For example, Bifidobacterium adolescentis JCM1275 could simultaneously retain 55.7 mg-Cs/g-dry cell and 8.1 mg-Sr/g-dry cell. These results demonstrated the potentials of utilizing complex biological system in simultaneous removal of multiple metal species.
Similar content being viewed by others


Introduction
The Chernobyl and Fukushima nuclear power plant accidents released large amounts of radioactive elements into the environment, which remained there for a long time1. Caesium (Cs) and strontium (Sr) have a high affinity for living organisms; therefore, when absorbed by the body, they cause prolonged internal exposure2,3. Since Cs and Sr are elements in the same group as potassium (K) and calcium (Ca), respectively, they behave similarly to these elements in the body, increasing the probability of cancer in the surrounding tissues4. Thus, a method for preventing these metals from being absorbed from the digestive tract is required.
Many of the methods used for environmental purification of metals5 cannot be used for removal of metals from the gastrointestinal tract. However, it may be possible to remove them using adsorbents by selecting appropriate edible materials. The ingestion of Prussian blue (iron ferrocyanide (III)) has been proposed as a means of facilitating Cs excretion4,6,7. This agent has high Cs-scavenging ability and low toxicity8. However, it also has several disadvantages, such as causing abdominal discomfort when ingested and requiring a high cost and large amount of time for collection after excretion9. In addition, despite the combined presence of Cs and Sr in many cases, Prussian blue can only remove Cs. Besides, alginic acid is known as an agent used to eliminate Sr from the body10. This reagent is found in edible seaweed and can be used without any major disadvantages. Eun et al9. fabricated a Prussian blue immobilized alginate aerogel and demonstrated it to be an effective method for removing Cs and Sr simultaneously from seawater. However, to the best of our knowledge, no method has been proposed for removing Cs and Sr simultaneously from the gastrointestinal tract.
In terms of edible removers, it is worth considering microorganisms. Many microorganisms that have been reported to have metal-removal capacities, such as lactobacilli11,12,13,14,15,16,17 and Saccharomyces cerevisiae18, have long been used as food products and can be easily consumed. Several mechanisms for metal removal by microorganisms have been reported19,20. In a previous study, we demonstrated that Lacticaseibacillus casei JCM1134 could adsorb metals, such as Sr, mainly through capsular polysaccharides present on the cell surface21. Therefore, if bacteria that adsorb Sr with their cell surface components can remove Cs by another mechanism such as bioaccumulation, it is possible to remove Cs and Sr simultaneously. In this study, we focused on lactobacilli, and bifidobacteria and investigated their potential to remove Cs and Sr.
Results and discussion
Distribution of Cs
The elemental distribution of L. casei JCM1134 and Bifidobacterium adolescentis JCM1275 cultured for 24 h in a medium supplemented with 100 mM Cs was analysed using an electron probe micro analyser. Cs was distributed in a granular form in both strains (Fig. 1). Moreover, phosphorus (P) and K were distributed in high amounts at sites with a high Cs content.

Caesium (Cs), phosphorus (P), and potassium (K) distribution and backscattered electron images (COMPO) of bacterial cells. Element maps show that the concentration of each element is higher at the point where the measurement level is higher.
Metal removal by microorganisms can be broadly classified into two categories: bioaccumulation, in which metals are taken up by living bacteria, and biosorption, in which metals are incorporated independently of microbial metabolism, such as adsorption to the surface layer of bacteria22. When Cs is removed by biosorption, it should be present evenly in the bacterial body; however, in Fig. 1, Cs was observed to be localized in both bacteria. On the other hand, Cs localization can be explained by considering that it is removed due to bioaccumulation. Some microorganisms store granular substances called polyphosphate inside their cells, which has a negative charge and can trap cations23. Therefore, considering that the cations K+ and Cs+ were trapped by polyphosphate, the results in Fig. 1 that P, K, and Cs were localized in similar locations are consistent. Kuwahara et al24. also mentioned the possibility that Cs could be trapped by polyphosphate in their study investigating Cs accumulation in Streptomyces sp. K202.
Hence, in L. casei JCM1134 and B. adolescentis JCM1275, Cs is likely removed via bioaccumulation. This suggests the possibility of simultaneously removing the two representative elements, Cs and Sr, using different removal mechanisms in one microorganism strain.
Growth inhibition by Cs
As the growth of microorganisms is inhibited when the Cs concentration is too high25,26, we compared the growth of microorganisms to determine the optimal amount of Cs that could support the growth of each microbe (Fig. 2). A Cs concentration of 50 mM had almost no effect on the growth of the lactobacilli strains (Fig. 2a–c); however, at 100 mM or higher, the growth rate and final cell concentration decreased. Three strains of Bifidobacterium (Fig. 2d–f) were more susceptible to Cs toxicity than were lactobacilli. For Bifidobacterium bifidum JCM1254 (Fig. 2d) and B. adolescentis JCM1275 (Fig. 2f), the cell concentrations after 24 h were generally the same at Cs concentrations of 50 mM or less. In contrast, in Bifidobacterium longum subsp. infantis JCM1260 (Fig. 2e), both the growth rate and final cell concentration decreased at 50 mM. Based on these results, Cs concentration of 50 mM was used for later experiments to minimize growth inhibition by Cs.

Growth curves of lactobacilli and bifidobacteria on Cs-supplemented medium. The vertical axis shows unit optical density at 680 nm (U.O.D.680). Each legend indicates the concentration of caesium supplemented to the medium. Error bars represent the standard error (n = 3).
It should be noted that a Cs concentration of 50 mM is not realistic, considering the actual gastrointestinal environment. For example, the concentration of 137Cs in food collected after the Chernobyl nuclear accident was as high as 1 × 104 Bq/kg1. When this was converted to a molar concentration assuming that 1 kg represents 1 L, it was approximately 2 × 10−11 M, which was far lower than the Cs concentration used in the present study. However, the K concentration has a large effect on the amount of Cs removed because they behave similarly27,28,29. The concentrations of K and other substances in the culture environment did not indicate an environment suitable for the gastrointestinal tract. We considered that there was no reason to set only the Cs concentration at a realistic value. Hence, an evaluation under practical conditions was not conducted in the present study, and the possibility of simultaneous removal of Cs and Sr was explored.
Cs removal capacity
To evaluate the amount of Cs that could be removed when only Cs was present (Fig. 3a), the Cs content of each strain was measured. As a result, 3.3–6.3 mg-Cs/g-dry cells were detected in lactobacilli and 23.8–70.5 mg-Cs/g-dry cells in Bifidobacterium (Fig. 4a). The accumulation of Cs was confirmed in all the bacteria used, and large differences were observed in the Cs accumulation capacity depending on the species.

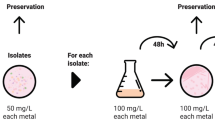
Experimental design for removal of Cs and Sr by bacteria.

Amount of Cs and Sr contained in each bacterium. Cs ( +) indicates the addition of 50 mM Cs to the culture medium, and Sr (+ ) indicates the suspension of cells in the solution containing 1 mM Sr. Error bars represent the standard error (n = 3).
We compared our current results on the Cs removal capacity with those previously reported for other Cs-removing bacteria. Kato et al30. investigated the Cs content per unit number of bacteria by Cs-resistant bacteria and reported 142.2 ± 7.1 mg-Cs/g-dry cell and 61.5 ± 6.9 mg-Cs/g-dry cell for Streptomyces sp. TOHO-2 and Streptomyces lividans TK24, respectively. Tomioka et al. reported 92 mg-Cs/g-dry cells for Rhodococcus erythropolis CS98 and 52 mg-Cs/g-dry cells for Rhodococcus sp. strain CS40226. The results of the present study showed that the three strains of lactobacilli had lower Cs content than the Bifidobacterium strains, with the highest removal capacity observed for B. adolescentis JCM1275 (70.5 mg-Cs/g-dry cells). The Cs removal capacity of B. adolescentis JCM1275 in the present study was similar to that previously reported for other Cs-removing bacteria.
Sr removal capacity
The Sr removal capacity was evaluated by suspending the cells cultured for 24 h in a buffer containing Sr and then measuring the amount of Sr contained in the cells (Fig. 3b). Sr was detected in the cells of all the strains used: 1.1–5.3 mg-Sr/g-dry cell for lactobacilli and 6.3–10.9 mg-Sr/g-dry cell for Bifidobacterium (Fig. 4b).
Based on our results, it is plausible that Sr is removed by biosorption, which occurs independently of bacterial metabolism, as Sr removal occurred without a nutrient source. In previous reports, the mechanism of Sr removal using microorganisms was considered to be through biosorption occurring on the surface of bacteria13,31. In addition, our previous study suggested that L. casei JCM1134 removed Sr via adsorption21.
Comparing the amount of Sr removed in the present study with that of other previously reported microbial adsorbents, 24.4 mg-Sr/g-dry cell was reported for Rhizopus nigricans32, 6.6 mg-Sr/g-dry cell for Rhizopus arrhizus33, and 10.0 mg-Sr/g-dry cell for S. cerevisiae18. Although making a direct comparison is difficult because the removal ability of microorganisms varies with culture and adsorption conditions, the Sr removal capacity of Bifidobacterium (6.3–10.9 mg-Sr/g-dry cell) in our study was similar to those previously reported for other microbial adsorbents. On the other hand, the removal ability of lactobacilli (1.1–5.3 mg-Sr/g-dry cell) was slightly lower than that of the previously reported microorganism adsorbents.
Simultaneous removal of Cs and Sr
To examine the amounts of Cs and Sr that one type of microbial strain can simultaneously retain, bacterial cells cultured in Cs-supplemented medium were suspended in Sr-containing buffer (Fig. 3c). As mentioned above, we expected the amounts of Cs and Sr to be irrelevant because they were thought to be removed by different mechanisms. However, contrary to expectations, all the strains showed a decrease in Cs content after Sr adsorption. The amount of released Cs differed depending on the strain, with B. adolescentis JCM1275 and B. longum subsp. infantis JCM1260 releasing 21% and 75%, respectively (Fig. 4a). In contrast, the amount of Sr removed was almost the same between cells cultured in the normal medium and those cultured in the Cs-supplemented medium (Fig. 4b).
The release of Cs after Sr adsorption can be explained by considering multiple mechanisms for Cs removal. As shown in Fig. 1, Cs is most likely retained in bacteria by trapping in polyphosphate; however, this does not mean that other mechanisms are not involved. Some previously published studies have reported Cs biosorption by the surface components of bacteria13,28,34,35, and it is possible that the microorganisms used in this study also adsorbed Cs on the surface. Therefore, the adsorbed Cs+ may have been replaced by the highly charged Sr2+ and released.
Although a certain amount of Cs was released, B. adolescentis JCM1275 showed high removal capacities of 55.7 mg-Cs/g-dry cell and 8.1 mg-Sr/g-dry cell. This indicates that with appropriate strain selection, it may be possible to remove multiple radioactive materials using a single type of microorganism.
Because some microbial functions change depending on the environment, it is necessary to investigate whether similar effects can be expected in an in vivo environment. Through these studies, it is expected that probiotic bacteria can be used as a highly accessible method for excreting radioactive elements from the gastrointestinal tract through fecal excretion.
Conclusion
Cs has been suggested to bioaccumulate in lactobacilli and Bifidobacterium. Therefore, we investigated whether Cs and Sr can be removed simultaneously using two mechanisms, bioaccumulation and biosorption, in combination with the metal-adsorption function of microorganisms. We showed that B. adolescentis JCM1275 simultaneously retained these two metals at high concentrations. If these functions were replicated in the gastrointestinal tract, they could be used to inhibit the absorption of multiple radionuclides into the human body.
Methods
Reagents
Glucose, K2HPO4, Polysorbate 80 (Tween 80; for biochemistry grade), sodium ascorbate (Wako special grade), l-cysteine hydrochloride, CsCl, H2O2, and SrCl2, were obtained from Fujifilm Wako Pure Chemical Co. (Osaka, Japan). These reagents without grade description were of guaranteed grade. Casein peptone and yeast extract were from Nihon Pharmaceutical Co. Ltd. (Osaka, Japan). MRS medium and beef extract were from Becton, Dickinson and Co. (Franklin Lakes, NJ, USA). Bis–Tris was obtained from MP Biomedicals (Santa Ana, CA, USA).
Culture conditions
Lactobacilli (Lactobacillus acidophilus JCM1028, Lactobacillus paragasseri JCM1130, and L. casei JCM1134) and bifidobacteria (B. bifidum JCM1254, B. longum subsp. infantis JCM1260, and B. adolescentis JCM1275) were provided by the Japan Collection of Microorganisms, RIKEN BRC which is participating in the National BioResource Project of the MEXT, Japan. Lactobacilli were cultured in MRS medium. The bifidobacterial medium contained 10 g/L casein peptone, 5.0 g/L beef extract, 5.0 g/L yeast extract, 10 g/L glucose, 3.0 g/L K2HPO4, and 1.0 mL/L Tween 80; 1.0% (w/v) sodium ascorbate and 0.05% (w/v) l-cysteine hydrochloride were added after autoclaving. Both bacteria were statically incubated at 37 °C. All experiments were performed after two pre-cultures with 1% (v/v) inoculum.
Microbial growth on Cs medium
CsCl was added to the medium at concentrations of 0, 10, 50, 100, and 200 mM, and each bacterium was cultured statically at 37 °C. Non-radioactive Cs was used in this study. The concentration of bacteria in the culture medium was monitored by measuring absorbance at 680 nm using a spectrometer UV-1200 (Shimadzu Co., Kyoto, Japan).
Element mapping
Bacteria cultured for 24 h in Cs-supplemented medium were washed with distilled water and diluted 20 times for lactobacilli and 10 times for Bifidobacterium. Thereafter, the bacterial culture solution was dropped onto a sample table, dried, and carbon deposition was performed using a vacuum deposition apparatus JFE-420 (JEOL Ltd., Tokyo, Japan). Backscattered electron and elemental mapping images of Cs, P, and K were obtained using an electron probe micro analyser JXA-8530F (JEOL Ltd.) at an accelerating voltage of 10.0 kV and a beam current of 5.0 × 10−8 A.
Determination of Cs contained in bacterial cells
The culture medium was incubated for 24 h with Cs concentration of 50 mM and centrifuged (6000 × g, 3 min) to collect the bacterial pellets, which were then suspended in distilled water and centrifuged twice to remove components derived from the culture medium. Wet digestion of bacterial cells was performed using a method modified from Tomioka et al26. The bacterial pellets were transferred to a conical beaker, 5 mL of nitric acid was added, and the watch glass was placed on the conical beaker and heated at 180 °C for 2–3 h. To decompose the sample, 0.5 mL of H2O2 was added until the colour of the solution became light pale yellow to colourless. The Cs concentration was determined using inductively coupled mass spectrometry (ICP-MS; ELAN DRC-e; PerkinElmer, Waltham, MA, USA) after dilution to the detectable concentration. The Cs content was converted to concentration per unit dry bacterial mass (mg-Cs/g-dry cell).
Sr adsorption amount
The experiment was conducted using a method modified from our previous study21. In brief, 2 mL of the culture medium was incubated for 24 h as described in section “Culture conditions” or “Determination of Cs contained in bacterial cells” was sampled and the bacteria were collected by centrifugation (6000 × g, 3 min). After washing, the bacteria were suspended in 2 mL of Bis–Tris buffer (1 mM, pH 6.8), SrCl2 (non-radioactive) was added to a final Sr concentration of 1 mM, and the bacterial suspension was incubated at 37 °C for 3 h (200 strokes/min, stroke length = 25 mm). After incubation, the bacterial cells were collected by centrifugation (6000 × g, 3 min), washed well, and subjected to acid decomposition as described in the “Determination of Cs contained in bacterial cells” section. The Sr content was then determined by ICP-MS.
Data availability
All data presented in this study are available from the corresponding author upon reasonable request.
References
Steinhauser, G., Brandl, A. & Johnson, T. E. Comparison of the Chernobyl and Fukushima nuclear accidents: A review of the environmental impacts. Sci. Total Environ. 470–471, 800–817 (2014).
Froidevaux, P., Bochud, F. & Haldimann, M. Retention half times in the skeleton of plutonium and 90Sr from above-ground nuclear tests: A retrospective study of the Swiss population. Chemosphere 80, 519–524 (2010).
Leggett, R. W., Williams, L. R., Melo, D. R. & Lipsztein, J. L. A physiologically based biokinetic model for cesium in the human body. Sci. Total Environ. 317, 235–255 (2003).
Aaseth, J., Nurchi, V. M. & Andersen, O. Medical therapy of patients contaminated with radioactive cesium or iodine. Biomolecules 9, 856 (2019).
Sheta, S. M. et al. Recent progress in high-performance environmental impacts of the removal of radionuclides from wastewater based on metal–organic frameworks: a review. RSC Adv. 13, 25182–25208 (2023).
Faustino, P. J. et al. Quantitative determination of cesium binding to ferric hexacyanoferrate: Prussian blue. J. Pharm. Biomed. Anal. 47, 114–125 (2008).
Le Gall, B., Taran, F., Renault, D., Wilk, J.-C. & Ansoborlo, E. Comparison of Prussian blue and apple-pectin efficacy on 137Cs decorporation in rats. Biochimie 88, 1837–1841 (2006).
Carniato, F. et al. More efficient Prussian blue nanoparticles for an improved caesium decontamination from aqueous solutions and biological fluids. Molecules 25, 3447 (2020).
Eun, S., Ryu, J., Kim, H., Hong, H.-J. & Kim, S. Simultaneous removal of radioactive cesium and strontium from seawater using a highly efficient Prussian blue-embedded alginate aerogel. J. Environ. Manag. 297, 113389 (2021).
Idota, Y. et al. Alginate enhances excretion and reduces absorption of strontium and cesium in rats. Biol. Pharm. Bull. 36, 485–491 (2013).
Arun, K. B. et al. Probiotics and gut microbiome−prospects and challenges in remediating heavy metal toxicity. J. Hazard. Mater. 420, 126676 (2021).
Feng, M., Chen, X., Li, C., Nurgul, R. & Dong, M. Isolation and identification of an exopolysaccharide-producing lactic acid bacterium strain from Chinese Paocai and biosorption of Pb(II) by its exopolysaccharide. J. Food Sci. 77, T111–T117 (2012).
Kinoshita, H. et al. In vitro mass-screening of lactic acid bacteria as potential biosorbents of cesium and strontium ions. J. Microbiol. Biotechnol. Food Sci. 4, 383–386 (2015).
Lin, Z., Zhou, C., Wu, J., Zhou, J. & Wang, L. A further insight into the mechanism of Ag+ biosorption by Lactobacillus sp. strain A09. Spectrochim. Acta A Mol. Biomol. Spectrosc. 61, 1195–1200 (2005).
Polak-Berecka, M., Szwajgier, D. & Waśko, A. Biosorption of Al(+3) and Cd(+2) by an exopolysaccharide from Lactobacillus rhamnosus. J. Food Sci. 79, T2404-2408 (2014).
Polak-Berecka, M. et al. Studies on the removal of Cd ions by gastrointestinal lactobacilli. Appl. Microbiol. Biotechnol. 101, 3415–3425 (2017).
Schut, S., Zauner, S., Hampel, G., König, H. & Claus, H. Biosorption of copper by wine-relevant lactobacilli. Int. J. Food Microbiol. 145, 126–131 (2011).
Chen, C. & Wang, J. Influence of metal ionic characteristics on their biosorption capacity by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 74, 911–917 (2007).
Fomina, M. & Gadd, G. M. Biosorption: current perspectives on concept, definition and application. Bioresour. Technol. 160, 3–14 (2014).
Yaashikaa, P. R., Kumar, P. S., Saravanan, A. & Vo, D.-V.N. Advances in biosorbents for removal of environmental pollutants: A review on pretreatment, removal mechanism and future outlook. J. Hazard. Mater. 420, 126596 (2021).
Endo, R. & Aoyagi, H. Adsorption preference for divalent metal ions by Lactobacillus casei JCM1134. Appl. Microbiol. Biotechnol. 102, 6155–6162 (2018).
Vijayaraghavan, K. & Yun, Y.-S. Bacterial biosorbents and biosorption. Biotechnol. Adv. 26, 266–291 (2008).
Günther, S. et al. Dynamics of polyphosphate-accumulating bacteria in wastewater treatment plant microbial communities detected via DAPI (4’,6’-diamidino-2-phenylindole) and tetracycline labeling. Appl. Environ. Microbiol. 75, 2111–2121 (2009).
Kuwahara, C. et al. Characteristics of cesium accumulation in the filamentous soil bacterium Streptomyces sp. K202. J. Environ. Radioact. 102, 138–144 (2011).
Kang, S.-M. et al. Cesium-induced inhibition of bacterial growth of Pseudomonas aeruginosa PAO1 and their possible potential applications for bioremediation of wastewater. J. Hazard. Mater. 338, 323–333 (2017).
Tomioka, N., Uchiyama, H. & Yagi, O. Cesium accumulation and growth characteristics of Rhodococcus erythropolis CS98 and Rhodococcus sp. strain CS402. Appl. Environ. Microbiol. 60, 2227–2231 (1994).
Kim, I. et al. Removal of radioactive cesium from an aqueous solution via bioaccumulation by microalgae and magnetic separation. Sci. Rep. 9, 10149 (2019).
Lan, T. et al. Biosorption behavior and mechanism of cesium-137 on Rhodosporidium fluviale strain UA2 isolated from cesium solution. J. Environ. Radioact. 134, 6–13 (2014).
Lee, K.-Y., Lee, S.-H., Lee, J. E. & Lee, S.-Y. Biosorption of radioactive cesium from contaminated water by microalgae Haematococcus pluvialis and Chlorella vulgaris. J. Environ. Manage. 233, 83–88 (2019).
Kato, F. et al. Accumulation and subcellular localization of Cesium in mycelia of Streptomyces lividans and a Cs tolerant strain, Streptomyces sp TOHO-2. J. Health Sci. 46, 259–262 (2000).
Ogawa, K. et al. Evaluation of Chlorella as a decorporation agent to enhance the elimination of radioactive Strontium from body. PLoS One 11, e0148080 (2016).
Kogej, A. & Pavko, A. Comparison of Rhizopus nigricans in a pelleted growth form with some other types of waste microbial biomass as biosorbents for metal ions. World J. Microbiol. Biotechnol. 17, 677–685 (2001).
Brady, J. M. & Tobin, J. M. Binding of hard and soft metal ions to Rhizopus arrhizus biomass. Enzyme Microb. Technol. 17, 791–796 (1995).
Sivaperumal, P., Kamala, K. & Rajaram, R. Adsorption of cesium ion by marine actinobacterium Nocardiopsis sp. 13H and their extracellular polymeric substances (EPS) role in bioremediation. Environ. Sci. Pollut. Res. 25, 4254–4267 (2018).
Yu, R. et al. Behavior and mechanism of cesium biosorption from aqueous solution by living Synechococcus PCC7002. Microorganisms 8, 491 (2020).
Acknowledgements
We thank Editage for English language editing.
Funding
This work was supported in part by Sumitomo Electric Industries Group Corporate Social Responsibility Foundation, 2023–2027 and Japan Society for the Promotion of Science, KAKENHI Grant-in-Aid for Scientific Research B (22H02474) [grants to H. A.].
Author information
Authors and Affiliations
Contributions
R.E.: Data curation, Formal analysis, Investigation, Validation, Visualization, Writing-original draft. S.K.: Conceptualization, Methodology, Validation. H.A.: Conception of research theme, Supervision, Project administration, Funding acquisition, Writing-review & editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Endo, R., Karasawa, S. & Aoyagi, H. Simultaneous removal of caesium and strontium using different removal mechanisms of probiotic bacteria. Sci Rep 14, 7630 (2024). https://doi.org/10.1038/s41598-024-57678-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-57678-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
