
Abstract
Solanum nigrum L. (Black nightshade), is one of the most troublesome weeds of summer crops such as corn, soybean, sunflower, etc. To study the effect of combined Castor oil as an adjuvant with different doses of Lumax (Mesotrion + S-metolacholor + Terbuthylazine) on the physiological behavior of Solanum nigrum L., a greenhouse experiment was conducted in randomized complete block design with four replications in agricultural faculty of the University of Tabriz in 2021. A foliar application of Lumax increased proline, malondialdehyde, and hydrogen peroxide concentrations and superoxide dismutase, catalase, and peroxidase activity. The content of protein and photosynthetic pigments (Chlorophyll a, b, and carotenoids) also decreased significantly by using Lumax herbicide. Applying castor oil in combination with Lumax intensifies oxidative stress and lipid peroxidation. Results showed that by increasing the herbicide doses in comparison with control (non-herbicide), Area, Fm, Fv, Fv/Fm, Fv/F0, Sm, Sm/Tfm, and Fv/F0 decreased 48.32%, 19.52%, 27.95%, 10.47%, 50.90%, 28.34%, 79.38%, and 50.90%, respectively and F0, F0/Fm increased 46.76% and 82.38%, respectively. Castor oil showed a synergistic effect on Lumax herbicide and enhanced its efficacy on Solanum nigrum. The presented results supported the view that by evaluating chlorophyll a fluorescence parameters, we would realize herbicide (alone or mixed with any adjacent) efficacy before the visual symptoms appear in the plant.
Similar content being viewed by others

Introduction
Solanum nigrum L. (Black nightshade) is a common and troublesome annual weed in most parts of the world1. It is incredibly vigorous, persistent, and hard to control after the seedling stage, dominating the crop2. It has been reported in many studies that Solanum nigrum L. causes serious damage to important crops, such as corn3,4,5,6,7. Therefore, effectively controlling this weed is vital for successful crop production. Within the range of available weed control practices, mechanical and chemical controls are the most commonly used methods under field conditions8.
Herbicides are one of the most effective means of eliminating weeds in agriculture due to their easy application and effectiveness. Their consumption has increased significantly in recent years, especially in countries with advanced agricultural systems. In the agricultural industry, about fifty percent of herbicides used on crops cause adverse effects on chloroplasts. Among the new pre-emergence herbicides known as photosystem II (PSII) inhibitors, Lumax is one of the most recently developed products9. The Lumax (537.5 SE) controls weeds such as annual grasses and broadleaf weeds by preventing them from germination and establishing new plants. Lumax is a combination of three herbicides, including mesotrione, s-metolachlor, and terbuthylazine, which act differently on different sites of the plant, inhibiting cell division, pigment synthesis, and photosynthesis, respectively10. Due to the potential for resistance to herbicides and pollution associated with the continuous use of these chemical compounds, researchers have developed auxiliary methods, including adjuvants, to reduce herbicide application dosages while increasing the effectiveness of these chemicals11.
Adjuvants are compounds added to the herbicide formulation or sprayer tank to enhance their efficacy12. Adjuvants increase the activity of herbicides by reducing the surface tension of spray solutions, increasing the contact surface and promoting wetting, increasing the penetration of herbicides by increasing the solubility of cuticle wax and increasing the retention of herbicide drops on plant surfaces13. Adjuvants such as surfactants and oil soften or dissolve cuticle wax and improve herbicide penetration by transferring the active ingredient from the surface of the target plant to its internal tissues14. Adjuvants through improving the performance of foliar-applied herbicides aim to optimize herbicide rates and minimize their cost and adverse effects. Enhancement in the efficacy of herbicides for effective weed control is of great importance, especially at a lower dosage and it could be considered as an approach to reducing application rates of herbicides15. Vegetable oils, as one of the adjuvants, have aroused a point of interest because of their less toxicity and risk, eco-compatibility, natural renewability, and biodegradability. The effect of adjuvant types to increase or decrease herbicides efficacy depends on the type, characteristics, and formulation of herbicides, weed species, and environmental conditions; therefore, to detect appropriate adjuvant for each herbicide, field experiments are required16.
Treatment of plants with herbicides induces physiological changes and leads to stress reactions. Visible damages do not always accompany these changes. The physiological responses to different stress factors such as herbicides are similar. The changes in pigment content17, proline quantity18, products of lipid peroxidation19, and chlorophyll a fluorescence20 are considered as possible stress-induced markers. However, which physiological-biochemical indexes of Solanum nigrum were changed under Lumax herbicide stress alone and in combination with castor oil? How did they change? Solving these problems is very important for the chemical control of Solanum nigrum, and there were seldom reports about those changes in weeds after herbicide treatment. So this study was done to survey the effect of Lumax herbicide alone and in combination with Castor oil on the physiological responses of Solanum nigrum during the early period after treatment with Lumax.
Materials and methods
Experimental condition
A study was undertaken to assess the physiological parameters of Solanum nigrum plants treated with different doses of Lumax (0, 25, 50, 75, and 100% of the recommended dosage (4.5 L ha−1)) with or without Castor oil at 0.3 (v/v). In 2021, an experiment utilizing a randomized complete block design with four replications was carried out in the glass greenhouse of the University of Tabriz under 14 h of natural light (27–32 °C). All accessions were obtained under national and international guidelines and the plant was collected under the supervision and permission of Tabriz University and all authors comply with all the local and national guidelines. Seeds of Solanum nigrum were collected from mature plants in field research at the University of Tabriz and placed in the open air for three months during the winter to break seed dormancy. In plastic pots (20 × 20 cm × 25 cm) filled with 1.0 kg of perlite, 15 Solanum nigrum seeds were planted at a depth of 3 cm, and then tap water (0.6 dS m-1) was added to achieve 100% field capacity. At the three-leaf stage, plants were thinned to five plants per pot. This study used Hoagland solution (Electrical conductivity = 1.3 ds m−1 and pH = 6.5–7) to compensate for the pots' losses21. Every 20 days, the perlite in the pots was washed to prevent a further increase in electrical conductivity due to the addition of the Hoagland solution21. After Solanum nigrum seedlings reached the 4–5 leaf stage, herbicide was applied once using the sprayer equipped with an XR11002-VP TeeJet nozzle, operating at a pressure of 3 bar and a velocity of 4 km h−1, delivering a spray volume of 200 L ha−1 on the upper side of the seedlings' leaves21. For measurement of proline, soluble protein contents, antioxidant enzymes, MDA, H2O2 and photosynthesis pigment three plants were destructively harvested from each repetition and then combined during the extraction stage (to reduce costs) and finally one sample was obtained for each repetition. However, two plants were considered for non-destructively chlorophyll fluorescence measurement and 3 clips were used in each plant (6 data in total for each repetition) and the average data of each parameter was considered for each repetition.
Measurement of proline and soluble protein contents
Three days after herbicide treatment, proline content was determined spectrophotometrically using ninhydrin, adopting the method of Bates et al.22. Fresh leaf tissues (300 mg) were homogenized in 3 mL of 3% sulphosalicylic acid. One milliliter each of acid ninhydrin and glacial acetic acid was added to the homogenate filtrate, and the reaction was carried out for one hour in a test tube placed in a water bath at 100 °C. A spectrophotometer at 520 nm was used to measure the absorbance of the mixture after it was extracted with toluene.
The total soluble protein contents were measured three days after herbicide treatment using the method proposed by Bradford23. Fresh leaf samples (1000 mg) were homogenized with 4 mL Na phosphate buffer (pH 7.2), transferred to cold centrifuge tubes (2 mL Eppendorf tubes), and centrifuged at 12,000 g for 10 min at four °C. Supernatants and dye were pipetted in spectrophotometer cuvettes, and the absorbance was recorded using a UV–Vis spectrophotometer at 595 nm.
Measurement of antioxidant enzymes
A sample of three Solanum nigrum plants was collected three days after herbicide treatment and kept in an ice container. The leaves were rinsed in distilled water to eliminate any remaining moisture. A mortar and pestle were chilled to crush approximately 0.5 g of fresh leaf pieces from each plot in an ice-cold 0.1 M phosphate buffer (pH 7.5) containing 0.5 mmol L EDTA. In a subsequent step, the homogenized material was centrifuged for 15 min at 15,000 g at four °C (Hettich, MIKRO 200, Germany). The mixture was used for enzyme assay.
The superoxide dismutase (SOD) activity was assayed according to Beyer and Fridovich (1987)24. One enzyme unit of SOD was defined as the amount of enzyme required to cause 50% inhibition of the rate of NBT (Nitroblue tetrazolium) reduction measured spectrometrically at 560 nm (Hitachi model U- 2000).
The activity of catalase (CAT) was determined according to Aebi25. The mixture reaction included 5.2 mL of 50 mM phosphate buffer (pH = 7.0), with 0.2 mL hydrogen peroxide (1%) and 0.3 mL of enzyme extract. The catalase activity was estimated by the decrease of absorbance at 240 nm for 1 min due to H2O2 consumption26.
The peroxidase (POD) activity was measured using Rao et al.27 technique. At 4 C, the extraction was performed. One gram of tissues was blended with 50 mmol/L potassium phosphate buffer (pH 7.0) in an ice-cold mortar before being centrifuged at 15,000 g for 15 min to obtain the filtrate for the detection of POD. A 50 µL enzyme extract solution was added to a 50 µL reaction mixture containing 20 mmol/L guaiacols, 2.8 mL of potassium phosphate buffer (pH 7.0), and 50 L of guaiacol (20 mmol/L) at 25º C. The reaction was initiated by adding 20 µL of H2O2 at a 40 mmol/L concentration. Every 30 s, changes in absorbance at 470 nm were observed. POD activity was expressed as 1 mol of tetraguaiacol per minute, and enzyme activity was reported in units per minute and mg of protein.
Measurement of MDA and H2O2
According to Jana and Choudhuri28 approach, the Hydrogen peroxide (H2O2) content was measured colorimetrically. In a 3 mL phosphate buffer (50 mM, pH = 6.5), homogenize 0.5 g of plant material (fresh leaves) for 25 min at 6000 g. The supernatant was then combined with 1 ml of titanium sulfate in 20 percent H2SO4 and centrifuged for 15 min at 6000 g. A UV–VIS spectrophotometer measured the absorbance at 410 nm (T80, PG Instruments, United Kingdom).
The decomposition product of lipid degradation, malondialdehyde (MDA), is a marker of lipid peroxidation status. About 0.5 g of fresh leaf samples were homogenized in a solution containing 20% (w/v) trichloroacetic acid and 0.5% 2-thiobarbituric acid to extract thiobarbituric acid-reactive substances responsible for lipid peroxidation. Immediately after heating for 30 min at 95 °C, this mixture was cooled in an ice bath to prevent further reaction. The absorbance of the supernatant was measured at 532 and 600 nm after 10 min of centrifugation at 5,000*g and 25 °C. Following the removal of the unwanted discoloration at 600 nm with a spectrophotometer, the molar extinction coefficient of MDA was calculated at 155 mmol/mol (L cm)29.
Measurement of photosynthesis pigment
Arnon's30 method was used to determine the chlorophyll a, b, and carotenoid content. Three days after herbicide treatment, fresh leaf samples (0.2 g) were ground to a powder in 10 mL of acetone at an 80% concentration and centrifuged at 10,000 rpm for five minutes. The chlorophyll a, b, and carotenoid content was measured with a spectrophotometer (Model Analytik Jena Spekol 1500 Germany) at three wavelengths: 663, 645, and 470 nm for chlorophyll a, chlorophyll b, and carotenoids, respectively.
Chlorophyll a fluorescence analysis
The chlorophyll fluorescence (ChlF) was monitored from the upper surface of fully expanded Solanum nigrum leaves using a Handy PEA fluorimeter (Hansatech Instruments Ltd., England) three days after herbicide application. Prior to measuring ChlF signals, the dark-adapted leaves (30–40 min) were exposed to solid actinic light (8000 μmol m−2 s−1), and fluorescence was recorded for a period of 20 µs to 2 s31. A PEA plus (1.10) program was then used to export ChlF signals32.
Statistical analysis
Based on the experimental design, the recorded data were ANOVA analyzed using SAS 9.1.3 software and Microsoft Office Excel 2020 software. Duncan's multiple range test was applied to compare the means of each trait at p ≤ 0.05.
Result
Proline and soluble protein contents
Analyses of variance related to Lumax herbicide alone or in combination with Castor oil on Solanum nigrum plants showed that the proline and soluble protein content, antioxidant enzymes activities, malondialdehyde, H2O2, and photosynthetic pigments were significantly affected by these treatments (Table 1).
Increasing herbicide dosage enhances the leaf proline content (Fig. 1a). Castor oil intensified proline content, increasing compared to herbicide application alone (Fig. 1a). The highest proline content in Solanum nigrum leaf was 2.10 (unit mol/gr FW), which belonged to 4.5 L ha−1 dose Lumax with Castor oil (Fig. 1a).

Changes in (a) proline and (b) soluble protein content of Solanum nigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil application. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.01).
The soluble protein content of Lumax-treated plants was lower than control (Fig. 1b). Use of Castor oil did not alter soluble protein content of leaf under control conditions (Fig. 1b). However, in plants treated with different doses of Lumax, application of castor oil caused more reduction in leaf soluble protein content (Fig. 1b). Under 4.5 L ha−1 with Castor oil, the lowest of soluble protein content in leaves observed (Fig. 1b).
Antioxidant enzyme activities
Data for the CAT, SOD, and POD activity of different treatments are illustrated in Fig. 2. As a result of Lumax treatment, the amount of CAT, SOD, and POD activity in the leaves increased (Fig. 2). CAT, POD, and SOD activities were significantly increased in Castor oil-treated leaves compared to non-Castor oil-treated leaves (Fig. 2). Compared to control, Lumax (4.5 L ha−1) + Castor oil increased CAT, POD, and SOD activities by 46.51%, 119.51%, and 77.77%, respectively (Fig. 2). In addition, POD and SOD activity did not significantly differ under control conditions when Castor oil was used (Fig. 2b,c).

Changes in (a) CAT, (b) POD, and (c) SOD activity of Solanum nigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.01).
MDA content and H2O2 concentration
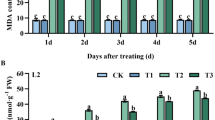
As shown in Fig. 3, Lumax had adverse effects on MDA content and H2O2 concentration of Solanum nigrum plants. The MDA content increased with increasing Lumax dosages, although no significant difference was observed between 1.125 and 2.25 L ha−1 (Fig. 3a). Furthermore, MDA content was significantly increased when Castor oil was added to Lumax (Fig. 3a). Generally, the highest MDA content was 3.12 umol/g FW in leaf, which belonged to Castor oil treatment under 4.5 L ha−1 Lumax (Fig. 3a).

Changes in (a) MAD and (b) H2O2 of Solanum nigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.01).
As Lumax dosages were increased, H2O2 concentration also increased (Fig. 3b). Castor oil did not alter the H2O2 concentration of leaf in the control treatment (Fig. 3b). However, combining Castor oil with herbicide increased the H2O2 concentration of Solanum nigrum plants (Fig. 3b). As compared to the control, the H2O2 concentration in 1.125, 2.25, 3.375, and 4.5 L ha−1 of Lumax + Castor oil increased 57.56%, 38.04%, 34.14%, and 25.36%, respectively (Fig. 3b).
Chlorophyll and carotenoids contents
The chlorophyll a, b, carotenoid, and total chlorophyll contents of Solanum nigrum leaves decreased while the ratio of chlorophyll a/b was augmented by increasing Lumax dosages (Table 2). Compared with non-castor oil control, at the highest dose of Lumax (4.5 L ha−1), chlorophyll a, b, carotenoid, and total chlorophyll decreased by 35.36%, 52.45%, 42.85%, and 40.29%, respectively (Table 2). In addition, mixing Castor oil with herbicide also decreased the content of chlorophyll a, b, carotenoid, and total chlorophyll and increased the ratio of chlorophyll a/b in leaves (Table 2). Irrespective of Lumax herbicide, chlorophyll a, carotenoid, and total chlorophyll is decreased using Castor oil (Table 2). The sharp decrease in chlorophyll b content compared to chlorophyll a caused the chlorophyll a/b ratio to increase with increasing herbicide doses (Table 2).
Chlorophyll a fluorescence
Analyses of variance showed significant effects of Lumax herbicide alone or in combination with Castor oil on chlorophyll a fluorescence parameters such as Tfm, Area, F0, Fm, Fv, F0/Fm, Fv/Fm, Fv/F0, and PI (Table 3).
Tfm and area
The results have indicated that increased dosages of Lumax led to a boost in the time required to reach maximum fluorescence (Tfm) (Fig. 4a). Although Tfm did not change by Castor oil application under non- lumax treatment, Castor oil usage increased Tfm under all dosages of Lumax herbicide (Fig. 4a). Under 4.5 L ha−1 Lumax with Castor oil, the highest of Tfm in leaves observed (Fig. 4a).

Changes in Tfm (a) and Area (b) of Solanum gigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.01).
The results presented in Fig. 4b showed that the PS II pool size or Area parameter differed markedly in Lumax and Castor oil-treated plants. Averaged over Castor oil, a decrease of 29% and 48.32% in Area were observed in the plants grown under 3.375 and 4.5 L ha−1 Lumax herbicide (Fig. 4b). Although the addition of Castor oil to the herbicide caused a further reduction in the area, this parameter did not change by Castor oil in the control condition (Fig. 4b).
F0 and Fm
Three days after herbicide application, the minimum fluorescence (F0) was increased by 46.76 and 82.05% at the 4.5 doses of Lumax alone and Lumax + Castor oil, respectively, compared with the control (Fig. 5a). All reaction centers in the photosystem II (PSII) at F0 are fully oxidized and ready to accept electrons4. Therefore, by increasing F0 following the mixing of Lumax herbicide with Castor oil, it can be said that most of the reaction centers remain closed, and the efficiency of PSII in electron transfer is diminished.

Changes in F0 (a) and Fm (b) of Solanum gigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.05).
The maximum fluorescence (Fm) also decreased with increasing herbicide doses (Fig. 5b). The addition of Castor oil to the herbicide tank increased the intensity of this decrease (Fig. 5b). Compared with the control, the Fm value in 1.125, 2.25, 3.375, and 4.5 L ha−1 of Lumax + Castor oil had declined 10.45%, 20.51%, 31.30%, and 38.08%, respectively (Fig. 5b).
Fv and F0/Fm
As Lumax doses increased, the variable fluorescence (Fv) decreased (Fig. 6a). The maximum dosage of Lumax (4.5 L ha−1) reduced the Fv parameter by 27.95 percent compared with the control (Fig. 6a). Castor oil reduced this parameter in minimal herbicide dosage (1.125 L ha−1) by 54.21% compared to control due to its synergistic effects with Lumax (Fig. 6a). Independently of Lumax, when Castor oil was sprayed to the Solanum nigrum plant, Fv decreased (Fig. 6a).

Changes in Fv (a) and F0/Fm (b) of Solanum gigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.05).
The fraction of sunlight absorbed by PSII that cannot be used photochemically and dissipated as heat (F0/Fm) demonstrated a significant increase by increasing herbicide dosages (Fig. 6b). Castor oil did not alter the F0/Fm value of leaf under non-herbicide conditions (Fig. 6b). However, under all Lumax treatments, the value of F0/Fm increased by application of Castor oil (Fig. 6b).
Fv/Fm, Fv/F0, and PI
In response to increasing Lumax dosage, maximum PSII quantum efficiency (Fv/Fm) decreased (from 0.88 at control to 0.79 at 4.5 L ha−1 of Lumax). Castor oil application with herbicide significantly changed Fv/Fm (Fig. 7a). Fv/Fm was only reduced by 23.75% at the highest dosage of Lumax (4.5 L ha−1) than the control (Fig. 7a). In this experiment, Fv/Fm also significantly decreased after adding Castor oil to the Lumax tank (Fig. 7a). Castor oil had more significant effects on the Fv/Fm parameter at high doses of herbicide than 25% of the recommended doses (Fig. 7a).

Changes in Fv/Fm (a), Fv/F0 (b), and PI (c) of Solanum nigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.01).
The efficiency of the water-splitting complex as an electron donor to PSII (Fv/F0) decreased with increasing herbicide dosage (Fig. 7b). This decrease was intensified by adding Castor oil to the herbicide composition (Fig. 7b). The reduction intensity of Fv/F0 at reduced doses of Lumax (2.25 and 3.375 L ha−1) was also significant compared to the recommended dosage (4.5 L ha−1) (Fig. 7b). At the maximum dosage of Lumax with and without Castor oil application, Fv/F0 decreased by 50.89% and 74.93%, respectively (Fig. 7b).
PI value, PSII performance index on an absorption basis, of Solanum nigrum leaves was significantly reduced by applying different dosages of Lumax herbicide (Fig. 7c). This reduction was more pronounced by Castor oil treatment (Fig. 7c). PI values decreased by 28.99% in control plants treated with Castor oil but untreated with Lumax (Fig. 7c). Lumax dose-dependently reduced this parameter (97.39%) when sprayed at 4.5 L ha−1 onto plants treated with Castor oil (Fig. 7c).
Sm and Sm/Tfm
The energy required to close all reaction centers (Sm) decreased with increasing dosage of Lumax, although there was no significant difference between 1.125, 2.25, and 3.375 L ha−1 (Fig. 8a). At 100% recommended dose of herbicide (4.5 L ha−1) in the absence of castor oil, Sm was reduced by 28.40% compared to the control (Fig. 8a). Castor oil with a synergistic effect on Lumax increased the intensity of Sm reduction. It caused a 92.20% reduction in this parameter compared to the control (Fig. 8a).

Changes in Sm (a) and Sm/Tfm (b) of Solanum nigrum in response to different doses of Mesotrion + S-metolacholor + Terbuthylazine with and without Castor oil. Each value is the mean of four replicates. Different letters represent significant differences (P < 0.01).
The Sm/Tfm ratio represents the intermediate redox state of Quinone A (QA) in the period from 0 to Tfm and, concomitantly, the average fraction of open reaction centers during the time needed to complete their closure. Increasing the herbicide dose caused a sharp reduction in this parameter of Solanum nigrum plants (Fig. 8b). Castor oil in combination with Lumax increased the reduction of oxidation and Quinone A in the mentioned time interval, so this reduction reached 78.57% in 4.5 L ha−1 of Lumax and 98.33% in4.5 L ha−1 of Lumax + Castor oil (Fig. 8b).
Discussion
In the present study, the soluble protein and photosynthetic pigments decreased; however, the activity of CAT, POX, and SOD and the content of proline, H2O2, and MDA increased by Lumax and Lumax + Castor oil treatments. As a result of environmental stress, proline is one of the first amino acids to increase33. Besides its role in protein structure, proline is an essential scavenger of reactive oxygen species (ROS) and is often discussed as a stress indicator34. Generally, the proline amount increases in glyphosate-treated plants35,36,37. These observations are also supported by the results presented here. Enhancing proline content in Solanum nigrum leaves under Lumax and Castor oil stress (Fig. 1a) is correlated with the enhancement of the activities of some enzymes such as Pyrroline-5-carboxylate synthase (P5CS), resulting from the activation of some gene expressions38. The soluble protein content is an essential indicator of the physiological health of plants39. Our results showed that soluble protein content was reduced in response to Lumax treatment in a dose-dependent pattern and Castor oil application. The decrease in the amount of protein under Lumax with and without Castor oil treatment could be justified as follows as reported by Souahi et al. (2016)40, some herbicides show detrimental effects on protein biosynthesis, amino acid absorption, and translocation or increasing the rate of protein degradation.
According to the results of adding Castor oil to the herbicide in this experiment and the research of other researchers, it can be said that oils are a group of adjuvants that increase the effectiveness of the herbicide by dissolving the cuticle and improving the penetration of the poison13. Rashed Mohassel et al. (2010)41 pointed out the increased effectiveness of diclofop-methyl and cycloxydim herbicides after mixing with olive and Castor oils in controlling Avena fatua L. and Phalaris minor L. weeds. Adding olive oil to the diclofop-methyl and cycloxydim herbicides to control Phalaris minor L. improved the efficiency of these two herbicides due to the faster dissolution of the cuticle wax of the weed leaves by olive oil41. Crop and mineral oils increase the control rate of Avena fatua L. and Lolium perenne L. by the herbicide clodinafop-propargyl42. According to Kargar et al. (2013)43, Cytogate and Castor oil can increase the effectiveness of mesosulfuron and iodosulfuron herbicides in controlling Stellaria media L.
In plant tissues, ROS are regulated by a system of antioxidant enzymes such as superoxide dismutase, catalase, peroxidase, and glutathione reductase, as well as non-enzymatic low molecular weight antioxidants such as glutathione, carotenoids, and tocopherols, and osmoprotectants such as proline44,45. Oxidative damage of herbicides is a common effect generated by ROS46. Previously, Jiang and Yang47 reported that herbicides at high concentrations increase CAT and POD activities due to the accumulation of H2O2 and O−2. The increase in SOD activity is correlated with the increase in oxidative stress, so activation of SOD in Solanum nigrum plants treated with Lumax + Castor oil indicates oxidative stress.
MDA content and H2O2 concentration increased with the increased peroxidation of structural lipids after applying Lumax + Castor oil (Fig. 3). MDA and H2O2 responses to oxidative stress are complex and may vary depending on factors such as stress intensity, duration, and plant species. MDA, the final product of lipid peroxidation, has increased due to glyphosate application in Zea mays L., Salvinia natans (L.), Hordeum vulgare L., Vallisneria natans (Lour.) Hara, and Solanum Lycopersicum L. plants36,37,48. In other studies, an inverse trend was found in which high herbicide concentrations led to a reduction and a tissue-specific decline of MDA36. After applying glyphosate, both reduction and elevation of H2O2 content were observed37,49,50,51.
Our results demonstrate that increasing osmotic and oxidative stresses in Solanum nigrum leave due to Lumax and Castor oil application reduced chlorophyll synthesis (Table 2). Increasing chlorophyll a/b ratio in these conditions indicated that chlorophyll b in Solanum nigrum plants is more sensitive to herbicide toxicity than chlorophyll a (Table 2). With the increase in glyphosate ratio, the accumulation of macroelements and microelements in plant tissues decreased, and photosynthetic parameters decreased52,53,54. Inhibitory effects of herbicides appear to be intensified with increasing concentrations. The reduction in the dry matter may be due to herbicides inhibiting chlorophyll synthesis55,56.
The fluorescence of chlorophyll a is a valid physiological indicator to determine the changes induced in the photosynthetic apparatus57. Several studies show the herbicides Asulam, Bifenax, 2,4-D, Glyphosate, Diclofop-methyl, Imazapir, Dicamba + 2,4-D, and Cetoxydim have no direct effect on photosynthesis, but they affect the fluorescence of chlorophyll a 58,59. The results of our study show that by increasing Lumax dosage, the plastoquinone pool size (Fig. 4b), the variable fluorescence (Fig. 6a), the maximum quantum efficiency of PSII (Fig. 7a), the water splitting efficiency (Fig. 7b), and the PSII performance index (Fig. 7c), the energy required to close all reaction centers (Fig. 8a) and the intermediate redox state of Quinone A in the period from 0 to Tfm (Fig. 8b) drastically decreased, and the PSII electron transport rate (Fig. 4a), the minimum fluorescence (Fig. 5a), and the fraction of sunlight absorbed by PSII that cannot be used photochemically and dissipated as heat (Fig. 6b) increased. After adding Castor oil to the herbicide composition, the intensity of these changes increased.
In most plants, the value of Fv/Fm is close to 0.83–0.85, and it is believed that under controlled conditions, this index is proportional to the rate of photosynthesis60. Changes in the Fv/Fm ratio may also be triggered by non-photochemical quenching61, protein D1 degradation, and the inactivation of PSII Reaction centers62. Fm reduction indicates that stress induces and decreases the activity of PSII57. High values of F0 under a high dosage of Lumax indicate distorted transport of excitation energy in PSII antennae and lower efficiency of energy trapping by the PSII reaction centers, which is most probably caused by the dissociation of LHCII (light-harvesting complex II) from PSII20. In investigating the effects of glyphosate on soybean (Glycine max L. Merr) in greenhouse conditions, it was observed that F0 and Fm were affected 48 h after the application of this herbicide35.
Stress conditions cause a decrease in the value of Area, which characterizes the transport of electrons to the pool of plastoquinones63. In plants growing under high doses of herbicide, the Fv/F0 ratio decreases; this points to a drop in the efficiency of the water-splitting reaction and weaker photosynthetic electron transport64,65. A prolonged Tfm period under herbicide stress might be due to the evident decrease in the value of the PSII performance index (PI) and disrupted water photolysis reaction on the donor side of PSII. Lumax ± Castor oil reduced PSI ends electron acceptors (Fig. 7c). The values of PI describe the energy flow efficiency of the photosynthetic transport chain beyond PSII66. According to Force et al.66 and our results in this research, PI, which describes the overall vitality of PSII, was the most sensitive indicator for enabling researchers to capture the effect of PAR fluctuations within a short time interval. The increase in Sm after Lumax and Castor oil treatment indicates an increase in the relative number of electron acceptors in the plastoquinone pool67.
Conclusion
It was shown that reactive oxygen species (ROS) damages rendered by Lumax herbicide decreased the protein and photosynthetic pigments and increased the MDA and proline contents, the concentration of H2O2, and the activity of antioxidant enzymes, i.e., CAT, POX, and SOD of Solanum nigrum plants. The actual mechanism behind ROS formation must be further clarified. On the other hand, the combination of Castor oil with Lumax herbicide disturbed the antioxidant system, increased the level of lipid peroxidation, and made the plants more sensitive to the adverse effects of Lumax. Using a mixture of Lumax and Castor oil disrupted the photosynthetic apparatus by interfering with water photolysis, reducing the size of the PQ pool, and slowing electron transport in PSII. This research proved the best control effect on Solanum nigrum when castor oil was mixed with Lumax at 75% (3.375 L/ha) of the recommended dose.
Data availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
References
Henriques, J., Lima, M., Rosa, S., Dias, S. & Dias, L. S. Allelopathic plants. XVIII. Solanum nigrum L.. Allelopathy J. 17, 1–16 (2006).
Kiran Kudesia, R., Rani, M. & Pal, A. Reclaiming degraded land in India through the cultivation of medicinal plants. Bot. Res. Int. 2, 174–181 (2009).
Sharma, N. & Rayamajhi, M. Different aspects of weed management in maize (Zea mays L.): A brief review. Adv. Agric. 2022, 1–10 (2022).
Porheidarghafarbi, S. Assessment of physiological and biochemical responses of corn (Zea mays L.) and its main weeds to mixing of herbicides with salicylic acid, solphat ammonium and citogit. PhD thesis of Weed Science. University of Tehran. Tehran, Iran. 2017 (In Persian with English abstract).
Soliman, I. E. & Gharib, H. S. Response of weeds and maize (Zea mays L.) to some weed control treatments under different nitrogen fertilizer rates. Zagazig J. Agri. Res. 38, 249–271 (2011).
Zand, E., Baghestani, M.A., Pourazar, R., Sabeti, P., Ghezeli, F., & Khayami, M. The evalution of nicosulfuron + rimsulfuron (ultima), mesotrion+S-Metolacholor+Terbuthlazine (Lumax 537.5SE) and Dynamic (Amicarbazon) new herbicides performance compared with common herbicides in Zea Mays L. field in Iran. J. Plant Prot. 23, 42–55 (2009).
Tanski, M. & Idziak, R. Influence of weed control regulation on herbicide efficacy and grain yield of maize. Prog. Plant Prot. 49, 349–352 (2009).
Jasinskas, A., Steponavičius, D., Šniauka, P. & Zinkevičius, R. Weed control by chemical and mechanical means. Weed Biol. Cont. https://doi.org/10.5772/60002 (2015).
Anorvey, V. Y., Asiedu, E. K. & Dapaah, H. K. Growth and yield of maize as influenced by using Lumax 5 SE for weed control in the transitional agro-ecological zone of Ghana. Inter. J. Plant Soil Sci. 21, 1–11 (2018).
Anorvey, V. Y., Dapaah, H. K. & Asiedu, E. K. Productivity and economic benefits of using different rates of Lumax 537.5 SE for weed control in maize in a transition agro-ecological zone of Ghana. J. Sci. Res. Rep. 17(6), 1–10 (2017).
Cobb, A. H. & Reade, J. P. H. Herbicides and Plant Physiology 2nd edn. (Wiley-Blackwell, West Sussex, 2010).
Rashed-Mohassel, M. H., Rastgo, M., Mousavi, S. K., Valiollahpor, R. & Hagigi, A. Principle of Weed Management 367 (University of Mashhad Press, 2006).
Hazen, J. L. Adjuvants terminology, classification and chemistry. Weed Tech. 14, 773–784 (2000).
Hall, F. R., Chapple, A. C., Downer, R. A., Kirchner, L. M. & Thacker, J. R. M. Pesticide application as affected by spray modifiers. Pest Manag. Sci. 38, 123–133 (1995).
Mousavi, S. K., Zand, E. & Saremi, H. Physiological function and application 378 (University of Zanjan Press, 2005).
Johnson, A. K., Roeth, F. W., Martin, A. R. & Klein, R. N. Glyphosate spray drift management with drift-reducing nozzles and adjuvants. Weed Tech. 20, 893–897 (2006).
Yordanov, I. Response of photosynthetic apparatus to temperature stress and molecular mechanisms of its adaptations. Photosynthetica 26(4), 517–531 (1992).
Reddy, P. & Veerajnegulu, K. Proline metabolism in senescing leaves of horse gram. J. Plant Physiol. 137, 381–383 (1991).
Sergiev, I., Alexieva, V., Yanev, S. & Karanov, E. Effect of atrazine and spermine on free proline and some antioxidants in pea (Pisum sativum L.) plants. Comp. Rend. Acad. Bulgari des Sci. 53(10), 63–66 (2000).
Hassannejad, S., Porheidarghafarbi, S. & Abbasvand, E. Herbicides and Plant Physiology 483 (University of Tabriz Press, 2017) (in Persian).
Hassannejad, S., Porheidar-Ghafarbi, S. & Lotfi, R. How photosynthetic light phase of spiny cocklebur (Xanthium spinosum L.) changes during times after herbicide application. J. Plant Physiol. Breed. 10(1), 61–70 (2020a).
Bates, L. S., Walden, R. P. & Teare, I. D. Rapid determination of proline for water stress studies. Plant Soil. 39, 205–207 (1973).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
Beyer, W. F. Jr. & Fridovich, I. Assaying for super oxide dismutase activity: Some large consequences of minor changes in conditions. Annual Bioch. 161, 559–566 (1987).
Aebi, H. Catalase in-vitro. Methods Enzym. 105, 121–126 (1984).
Havir, A. E. & McHale, N. A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 84, 450–455 (1987).
Rao, M. V., Paliyath, C. & Ormrod, D. P. Ultraviolet-B and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 110, 125–136 (1996).
Jana, S. & Choudhuri, M. A. Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquatic Bot. 12, 345–354 (1982).
Kosugi, H. & Kikugawa, K. Thiobarbituric acid-reactive substances in chicken fat and unsaturated fatty acids. J. Food Sci. 50, 1181–1182 (1985).
Arnon, D. I. Copper enzymes in isolated chloroplasts polyphenol oxidase in Beta vulgaris. Plant Physiol. 24, 1–15 (1949).
Abbasvand, E., Hassannejad, S., Zehtab Salmasi, S. & Alizadeh Salteh, S. Effects of seed priming with salicylic acid on chlorophyll a fluorescence parameters of basil (Ocimum basilicum L.) infested by field dodder (Cuscuta campestris Yunk). J. Plant Physiol. Breed. 9(2), 11–18 (2019).
Hassannejad, S. et al. Early Identification of herbicide modes of action by the use of chlorophyll fluorescence measurements. Plants 9, 529 (2020).
Ashraf, M. & Foolad, M. R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 59, 206–216 (2007).
Kishor, P. B. K. & Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homoeostasis a more critical issue?. Plant Cell Environ. 37, 300–311 (2014).
Huang, J., Silva, E. N., Shen, Z., Jiang, B. & Lu, H. Effects of glyphosate on photosynthesis, chlorophyll fluorescence and physico-chemical properties of cogongrass (Imperata cylindrical L.). Plant Omics. 5, 177–183 (2012).
Soares, C., Pereira, R., Spormann, S. & Fidalgo, F. Is soil contamination by a glyphosate commercial formulation truly harmless to non-target plants? Evaluation of oxidative damage and antioxidant responses in tomato. Environ. Poll. 247, 256–265 (2019).
Spormann, S., Soares, C. & Fidalgo, F. Salicylic acid alleviates glyphosate-induced oxidative stress in Hordeum vulgare L. J. Environ. Manag. 241, 226–234 (2019).
Szekely, G. et al. Duplicated P5CS genes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. Plant J. 53, 11–28. https://doi.org/10.1111/j.1365-313X.2007.03318.x (2008).
Doganlar, Z. B., Demir, K., Basak, H. & Gul, I. Effects of salt stress on pigment and total soluble protein contents of three different tomato cultivars. African J. Agri. Res. 5(15), 2056–2065 (2010).
Souahi, H., Meksem Amara, L. & RedaDjebar, M. Effects of sulfonylurea herbicides on protein content and antioxidants activity in wheat in semi-arid region. Int. J. Adv. Eng. Manag. Sci. 2, 1471–1476 (2016).
Rashed-Mohassel, M. H., Aliverdi, A., Hamami, H. & Zand, E. Optimizing the performance of diclofop-methyl, cycloxydim, and clodinafop-propargyl on littleseed canarygrass (Phalaris minor) and wild oat (Avena ludoviciana) control with adjuvants. Weed Biol. Manag. 10, 57–63 (2010).
Stagnari, F. & Onofri, A. Influence of vegetable and mineral oils on the efficacy of some post-emergence herbicides for grass weed control in wheat. Pest. Sci. Soc. Jap. 31, 339–343 (2006).
Kargar, M., Rashed-Mohassel, M. H., Nezami, A. & Darbandi, I. Evaluation effect of adjuvant on mesosulfuron+ iodosulfuron herbicide performance on littleseed canarygrass control. J. Plant Prot. 29(3), 35 (2015) (in Persian).
Bolkhania, O., Virolainen, E. & Fagerstedt, K. V. Antioxidant, oxidative damage and oxygen deprivation stress: A Review. Ann. Bot. 91, 179–194 (2003).
Hayat, Sh. et al. Role of proline under changing environments. Plant Sign. Behav. 7, 1456–1466 (2012).
Lukatkina, A. S., Garkova, A. N., Bochkarjova, A. S., Nushtaeva, O. V. & Teixeira da Silva, J. A. Treatment with the herbicide TOPIK induces oxidative stress in cereal leaves. Pest. Biochem. Physiol. 105, 44–49 (2013).
Jiang, L. & Yang, H. Prometryne-induced oxidative stress and impact on antioxidant enzymes in wheat. Ecot. Environ. Saf. 72, 1687–1693 (2009).
Zhong, G., Wu, Zh., Yin, J. & Chai, L. Responses of Hydrilla verticillata (L.f.) Royle and Vallisneria natans (Lour.) Hara to glyphosate exposure. Chemosphere. 193, 385–393 (2018).
Miteva, L., Tsoneva, J., Ivanov, S. & Alexieva, V. Alterations of the content of hydrogen peroxide and malondialdehyde and the activity of some antioxidant enzymes in the roots and leaves of pea and wheat plants exposed to glyphosate. C. R. Academic Bulgare des Sci. 58, 733–738 (2005).
Sergiev, I. G., Alexieva, V. S., Ivanov, S. V., Moskova, I. I. & Karanov, E. N. The phenylureacytokinin 4PU-30 protects maize plants against glyphosate action. Pest. Biochem. Physiol. 85, 139–146 (2006).
De Freitas-Silva, L. et al. Glyphosate-induced oxidative stress in Arabidopsis thaliana affecting peroxisomal metabolism and triggers activity in the oxidative phase of the pentose phosphate pathway (OXPPP) involved in NADPH generation. J. Plant Physiol. 218, 196–205 (2017).
Gomes, M. P. et al. Differential effects of glyphosate and aminomethylphosphonic acid (AMPA) on photosynthesis and chlorophyll metabolism in willow plants. Pest. Biochem. Physiol. 130, 65–70 (2016).
Singh, H., Singh, N. B., Singh, A., Hussain, I. & Yadav, V. Physiological and biochemical roles of nitric oxide against toxicity produced by glyphosate herbicide in Pisum sativum. Rus. J. Plant Physiol. 64, 518–524 (2017).
Harre, N. T., Nie, H., Jiang, Y. & Young, B. G. Differential antioxidant enzyme activity in rapid-response glyphosate resistant Ambrosia trifida. Pest Manag. Sci. 74(9), 2125–2132 (2018).
Caux, P. Y., Menard, L. & Kent, R. Comparative study of the effects of MCPA, butylate, atrazine and cyanazine on Selenastrum capricornutum. Environ. Poll. 92(2), 219–225 (1996).
Okmen, G., Turkcan, O. & Redal, P. Effect of herbicides on chlorophyll-a, β- caroten, phycocyanin and allophycocyanin content of Anabaena Sp.. J. App. Biol. Sci. 7(2), 20–27 (2013).
Mehta, P., Jajoo, A., Mathur, S. & Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol. Biochem. 48, 16–20 (2010).
Barbagallo, R. P., Oxborough, K., Pallett, K. E. & Baker, N. R. Rapid, non-invasive screening for perturbations of metabolism and plant growth using chlorophyll florescence imaging. Plant Physiol. 132, 485–493 (2003).
Hammami, H., Mohassel, M. H., Parsa, M., Bannayan-aval, M. & Zand, E. Behavior of sethoxydim alone or in combination with turnip oils on chlorophyll fluorescence parameter. Notulae Sci. Biol. 6(1), 112–118 (2014).
Björkman, O. & Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170, 489–504 (1987).
Maxwell, K. & Johnson, G. N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 51, 659–668. https://doi.org/10.1093/jexbot/51.345.659 (2000).
Kalaji, M. H. & Guo, P. Chlorophyll fluorescence: A useful tool in barley plant breeding programs. Photochem. Res. Progress 29, 439–463 (2008).
Murchie, E. H. & Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 64, 3983–3998 (2013).
Hassannejad, S. & Porheidarghafarbi, S. Assessment of some chlorophyll a fluorescence parameters of different corn cultivars in response to clodinafop-propagrgyl herbicide and salicylic acid. J. Plant Physiol. Breed. 8(1), 47–57 (2018).
Strasser, R. J., Tsimilli-Michael, M. & Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence. A Signature of Photosynthesis (eds Papageorgiou, G. C. & Govindjee) 321–362 (Springer, 2004).
Force, L., Critchley, C. & Van Rensen, J. J. S. New fluorescence parameters for monitoring photosynthesis in plants. Photo. Res. 78(1), 17–33 (2003).
Balabanova, D. A. et al. Photosynthetic performance of the imidazolinon resistant sunflower exposed to single and combined treatment by the herbicide imazamox and an amino acid extract. Front. Plant Sci. 5, 1–10 (2016).
Author information
Authors and Affiliations
Contributions
B.F., E.A., S.P.G. and Z.N. carried out the experiment and wrote the manuscript with support from S.H. S.H. supervised the project. All authors discussed the results and contributed to the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hassannejad, S., Fadaei, B., Abbasvand, E. et al. Early response of Solanum nigrum L. to Lumax and castor oil combination in relation to antioxidant activity, osmolyte concentration and chlorophyll a fluorescence. Sci Rep 13, 409 (2023). https://doi.org/10.1038/s41598-023-27428-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-27428-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
