
Abstract
We aimed to determine SARS-CoV-2 antibody seropositivity among pregnant women and the transplacental transfer efficiency of SARS-CoV-2-specific antibodies relative to malaria antibodies among SARS-CoV-2 seropositive mother-cord pairs. This cross-sectional study was conducted in Accra, Ghana, from March to May 2022. Antigen- specific IgG antibodies against SARS-CoV-2 (nucleoprotein and spike-receptor binding domain) and malarial antigens (circumsporozoite protein and merozoite surface protein 3) in maternal and cord plasma were measured by ELISA. Plasma from both vaccinated and unvaccinated pregnant women were tested for neutralizing antibodies using commercial kit. Of the unvaccinated pregnant women tested, 58.12% at antenatal clinics and 55.56% at the delivery wards were seropositive for both SARS-CoV-2 nucleoprotein and RBD antibodies. Anti-SARS-CoV-2 antibodies in cord samples correlated with maternal antibody levels (N antigen rs = 0.7155, p < 0.001; RBD rs = 0.8693, p < 0.001). Transplacental transfer of SARS-CoV-2 nucleoprotein antibodies was comparable to circumsporozoite protein antibodies (p = 0.9999) but both were higher than transfer rates of merozoite surface protein 3 antibodies (p < 0.001). SARS-CoV-2 IgG seropositivity among pregnant women in Accra is high with a boost of SARS-CoV-2 RBD-specific IgG in vaccinated women. Transplacental transfer of anti-SARS-CoV-2 and malarial antibodies was efficient, supporting vaccination of mothers as a strategy to protect infants against SARS-CoV-2.
Similar content being viewed by others

Introduction
In May 2023, the World health organisation (WHO) declared coronavirus disease (COVID-19) pandemic which began in 2019 over. As at the time of declaration, there were an estimated 765 million confirmed cases of COVID-19 globally, with Africa accounting for less than 2% of the confirmed COVID-19 cases1. COVID-19-related morbidity and mortality in sub-Saharan Africa is comparatively lower than that in developed countries2. Significantly low testing rates in many African countries due to limited technical capacity and scarce resources mean that the true disease burden of COVID-19 is underestimated3. In addition, reduced testing leads to the challenge of identifying at-risk populations within each local context and targeting vulnerable populations for interventions.
SARS-CoV-2 infection during pregnancy, particularly in the third trimester, increases the risk of severe disease and results in poor obstetric outcomes4,5,6. Due to the vulnerability of pregnant women, many countries have prioritized pregnant women for interventions such as vaccination. Over 60% of global pregnancies occur in resource-limited settings, where the rates of maternal mortality and adverse pregnancy outcomes are disproportionately high in relation to developed parts of the world7. It is therefore critical to determine the burden of SARS-CoV-2 exposure among the pregnant population in Africa to inform population-based policy decisions in order to mitigate the impact of COVID-19 and any future disease threats on maternal and neonatal health. Several seroepidemiological studies from different African countries have reported significantly high SARS-CoV-2 seropositivity rates within a wider population, which confirms the underestimation of COVID-19 cases in Africa based on reported testing figures8,9,10. It is possible COVID-19 burden among the pregnant population in Africa might diverge from estimates in most developed countries. There are a few studies examining the prevalence of SARS-CoV-2 infection among African pregnant women10,11,12,13,14,15 which point to an increase in SARS-CoV-2 seropositive rates over the course of the pandemic. However, only one study examined effects of prenatal COVID-19 exposure on birth outcomes15 and transplacental efficiency of SARS-CoV-2 IgG alone but did not compare SARS-CoV2 transfer efficiency to other antigen-specific IgG.
Aside COVID-19 pandemic, sub-Saharan Africa remains endemic to other infectious diseases, such as malaria, tuberculosis, and HIV. Over 90% of global malaria-associated morbidity and mortality occur in sub-Saharan Africa, with pregnant women and children under five years having the greatest risk of severe disease. However, in such endemic populations, infants 0–6 months rarely develop malaria and are resistant to severe disease, a phenomenon attributed to the placental transfer of malaria-specific immunoglobulins (Igs) from mothers to fetuses16. The presence of infections can affect the efficiency of maternofetal transfer of IgG from mothers to newborns, as has been shown in HIV-infected pregnant women17. Studies conducted in developed countries have shown reduced placental transfer of SARS-CoV-2-specific IgG from mothers to infants18,19. In sub-Saharan Africa where infectious disease burden is high, it is therefore critical to understand how COVID-19 infection in pregnancy affects the efficiency of placental transfer of IgG against common infections such as malaria.
In the present study, we determined SARS-CoV-2 antibody seropositivity among pregnant women attending routine antenatal clinics and parturient women admitted to delivery wards. We further examined the transplacental transfer efficiency of SARS-CoV-2-specific antibodies in relation to the transfer of malaria-specific antibodies among mother-infant dyads.
Results
Participant characteristics
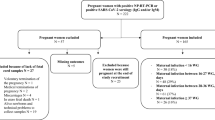
Pregnant women were recruited from Maamobi General Hospital in Accra, Ghana. Participants’ age ranged from 15 to 47 years (Table 1). Over 80% of the study population had at least some basic education with most of them employed in the informal sector. Few (5%) of our participants had existing comorbidities. Only 9.2%(55/599) of the participants had received at least a single dose of COVID-19 vaccines. All Naso- and oropharyngeal swabs from the pregnant women tested in RT-PCR were negative for SARS-CoV-2 indicating none of the pregnant women attending ANC or admitted to the delivery wards had an active SARS-CoV-2 infection during the study period.
SARS-CoV-2 IgG prevalence
At the time of conducting the study, vaccination was recommended for pregnant women in Ghana. In examining SARS-CoV-2 antibody seropositivity, individuals who had received at least one dose of COVID-19 vaccines were excluded from this analysis (Fig. 1). Of the 351 women at the antenatal clinics who were unvaccinated, 204 (58.12%, 95% CI 52.75–63.30%) were seropositive for both SARS-CoV-2 N antigen and RBD, 108 (30.77%, 95% CI 26.04–35.93%) and 13 (3.7%, 95% CI 2.07–6.41%) showed IgG reactivity to only N antigen and RBD respectively (Fig. 1a). The remaining 27 (7.69%, 95% CI 5.22–11.12) women were seronegative against both antigens. A similar patten was observed among the parturient women at the delivery wards (Fig. 1b). Out of the 180 women who were unvaccinated and were tested, 100 (55.56%, 95% CI 47.98–62.89) elicited a positive IgG response against both N antigen and RBD, 61 (33.89%, 95% CI 27.12–41.36) were IgG positive for N antigen alone and 6 (3.33, 95% CI 1.36–7.45) were IgG positive for RBD alone. In all, 13 (7.22%, 95% CI 4.06–12.30) women at the delivery wards did not have antibodies against neither N antigen or RBD.

Seroprevalence to SARS-CoV-2 is high in pregnant women. IgG antibody seropositivity of SARS-COV-2 N-antigen (a) and RBD antigen (b) in plasma from pregnant women at antenatal and delivery wards. Seropositivity against N antigen alone (blue), RBD alone (green) and both antigens (grey).
Transplacental transfer efficiency of SARS-CoV-2 antibodies
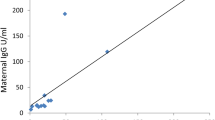
We next determined the transfer efficiency of SARS-CoV-2 antibodies between mothers who were seropositive (n = 170) and their matching cord samples (Fig. 2). SARS-CoV-2 N antigen and RBD-specific IgG antibody levels in cord plasma was significantly high than in mothers (p < 0.0001, Fig. 2A; p < 0.0032, Fig. 2B). We found positive correlations between seropositive mother plasma antigen-specific antibody levels and corresponding cord sample for both N antigen (r = 0.7155, p < 0.0001, Fig. 2C) and RBD (r = 0.8693, p < 0.0001, Fig. 2D).

Placental transfer of IgG between seropositive mothers to matching cord. SARS-CoV-2 antibodies against nucleoprotein (N) protein (A) and RBD (B) between mother:cord pairs. Spaghetti lines represent antibody levels between mother and matching cord sample. Box and whisker represent median with interquartile range. Correlation between maternal and cord IgG antibody levels against nucleoprotein (C) and RBD (D) presented in linear regression. Shaded are depicts 95% confidence interval. r represents the spearman’s rank coefficient .
As a comparator for transplacental efficiency, we estimated the cord:mother transfer ratio of antibodies against malaria pre-erythrocytic stage antigen CSP and merozoite antigen MSP3 among SARS-CoV-2 seropositive mothers and paired cord samples (Fig. 3a).There was no difference in the mean transfer ratio between N antibodies (mean = 1.08, 95% CI 0.94–1.24) and CSP antibodies (mean = 1.35, 95% CI 1.20–1.51). Both N antigen and CSP antibody transfer ratio was significantly high than RBD (mean = 0.89, 95% CI 0.89–1.06) and MSP3 antibody transfer ratio (mean = 0.733, 95% CI 0.73–0.84) with MSP3 being the least transferred. We subsequently compared the placental transfer ratios of CSP and MSP3 antibodies between SARS-CoV-2 seropositive mother:cord pairs and paired mother:cord plasma samples collected between 2016–2017 before the onset of COVID-19 pandemic. There was no difference in the cord:mother transfer ratios of CSP antibodies between SARS-CoV-2 seropositive individuals and the prepandemic samples (Fig. 3b). In contrast, MSP3-specific cord:mother transfer ratios were significantly high in the prepandemic samples than the SARS-CoV-2 seropositive individuals (Fig. 3c). Generally, CSP transfer ratio was high compared to MSP3.

Transfer efficiency of SARS-CoV-2 and malaria-specific antibodies. Cord:maternal antibody transfer ratio of antigens among SARS-CoV-2 seropositive individuals (a). Transfer ratio of CSP (b) and MSP3 (c) among SARS-CoV-2 seropositive paired mother:cord plasma samples and pre-pandemic paired mother:cord plasma samples. Transfer ratio was calculated as: (cord IgG concentration)/(maternal IgG concentration). Kruskal Wallis test was used to analyse comparisons in A. Mann–Whitney test was used to compare transfer ratios in b and c. Data is presented as geometric mean (thick horizontal lines) and 95% confidence interval (error bars).
Transfer of SARS-CoV-2 neutralizing antibodies
To examine the functionality of SARS-CoV-2 antibodies induced in individuals, we used plasma from a subset of the pregnant women who were seropositive for SARS-CoV-2 in a competitive ELISA (Fig. 4). Significantly high proportion of the seropositive pregnant women both at ANC (60/69) and delivery wards (n = 41/44) had neutralizing antibodies (Fig. 4a). Among the parturient women tested, the neutralizing antibodies were effectively transferred to cord (Fig. 4b) except between mothers who were seronegative for neutralizing antibodies and their matching cord.

Inhibition of ACE2 binding in SARS-CoV-2 seropositive pregnant. Percentage binding inhibition between seropositive pregnant women at antenatal clinics and at delivery (a). Inhibitory antibodies transferred from mothers to cord (b). Bars indicate median binding inhibition. Horizontal dashed lines represent 50% inhibition.
Vaccination boost spike- RBD antibodies
Our results show that a significantly high proportion of the participants had been naturally exposed to SARS-CoV-2 over the course of pandemic. Thus, we compared SARS-CoV-2 antibody levels between pregnant women naturally exposed to COVID-19 who were unvaccinated and naturally exposed women who had received at least a single dose of COVID-19 vaccine before or during pregnancy (Fig. 5). Median plasma N-antigen IgG levels were significantly increased among women who were unvaccinated compared to vaccinated women (p = 0.0192, Fig. 5a). Conversely, anti-RBD IgG levels were significantly boosted in the vaccinated women relative to the unvaccinated naturally exposed women (p = 0.0016, Fig. 5a). However, the percentage inhibition of ACE-2 binding was comparable (p = 0.2065) between vaccinated (median = 95.53%) and unvaccinated pregnant women (95.51%).

Vaccination against SARS-CoV-2 causes a boost in spike-RBD specific antibodies. Antibody levels against N antigen (a) and Spike-RBD (b) between pregnant women vaccinated against COVID-19 and unvaccinated pregnant women. ACE-2 binding inhibition rates between pregnant women vaccinated against COVID-19 and unvaccinated pregnant women (c). Mann–Whitney test was used to analyse comparisons between groups.
Discussion
Pregnancy predisposes pregnant women to severe disease outcomes when infected with SARS-CoV-2 compared to non-pregnant individuals. In Africa, COVID-19 testing capacities during the pandemic was limited in many countries particularly in sub-Saharan Africa. Levels of SARS-CoV-2 exposure among pregnant women in sub-Saharan is not fully known as data from such serosurveys is important in designing interventions targeting the vulnerable pregnant population to current and future disease threats. Our key findings suggest SARS-CoV-2 antibody prevalence among pregnant women in urban Accra is high (55–58%). COVID-19 vaccination rate was markedly low among our participants. We found that SARS-CoV-2 RBD-specifc IgG is significantly boosted in vaccinated women relative to unvaccinated women. SARS-CoV-2 specific antibodies are efficiently transferred between mother and neonates as there was strong correlation between mother and cord samples. The transplacental transfer rate of SARS-CoV-2 antibodies was comparable to malaria-specific antibodies among SARS-CoV-2 seropositive individuals.
Previous studies examining SARS-CoV-2 seropositivity among HIV-infected pregnant women in Mozambique revealed a 11.3% seroprevalence in 202114. Similar studies conducted in Ethiopia recorded a prevalence of 5.7% in pregnant women between April 2020 and March 202120. In Accra, Ghana, nucleoprotein seropositivity rates among the general non-pregnant population was found to have increased from 19.8% in November, 2020 to 42.7% in July, 20219. Our seroprevalence data is much higher than the seroconversion rates recorded in the earlier study in Ghana and other African countries. At the time of conducting the present study, Ghana had experienced 4 major waves21. The high seroprevalence rates we observed is suggestive of ongoing transmission which increased with each subsequent wave as demonstrated in a study conducted among pregnant women in Gambia between 2020 and 202110. Although majority of the seropositive individuals had detectable antibodies against both the nucleoprotein and RBD antigens, some individuals showed seropositivity against only one antigen. Similar observations have been made in earlier serosurveys among the wider non-pregnant population in Ghana9 and other LMICs22. It is thought that in some individuals, there is preferential seroconversion of antibodies against one antigen over the other. This was reported in comparative antibody profile studies in children and adults where children were observed to have mainly anti-spike antibodies but not anti-nucleoprotein antibodies23.
Based on the physical examination and interviews, none of our study participants were symptomatic and seropositivity was not associated with any adverse birth outcomes. Studies in other populations in both developed and resource-limited settings have shown that PCR-confirmed COVID-19 diagnosis in pregnant women increased risk of adverse birth outcomes compared to undiagnosed pregnant women5,24,25.
Transplacental SARS-CoV-2 RBD IgG transfer was found to be impaired in symptomatic pregnant women relative to asymptomatic pregnant women18,19,26. In the present study, we observed efficient transfer of SARS-CoV-2 nucleoprotein and RBD antibodies from mothers to neonates. Our results are in agreement with similar studies which found passive transfer of SARS-CoV-2 antibodies between PCR-positive pregnant women who were asymptomatic27. Since all our participants are asymptomatic, it is plausible some women were infected early on in their pregnancies allowing enough time for mothers to develop antibodies which are transferred to neonates. Antibodies in cord samples strongly correlated with maternal antibody levels in line with studies both in natural infections and vaccination27,28,29. Transplacental rates of malaria-specific antibodies (anti-CSP and MSP3) were similar between SARS-CoV-2 seropositive mother:cord pairs and pre-pandemic paired mother:cord samples in contrast to reports by others who found reduced transfer rates of influenza and diphtheria antibodies in COVID-19 infected pregnancies compared to uninfected pregnancies19. These observations are reassuring considering that malaria remains an important public health disease in sub-Saharan Africa and the passive transfer of malaria-specific antibodies are important in immunity against malaria in the first six months of life.
At the time of conducting the study, COVID-19 vaccination was recommended for pregnant women by the Ghana Health Service. Despite our participants knowledge about the importance of vaccination in protection against COVID-1930, vaccine uptake was significantly low. Vaccination significantly increased SARS-CoV-2 RBD antibodies in the vaccinated women compared to natural infections. Our finding is unsurprising given that the COVID-19 vaccines target the spike RBD. Regardless of the marked increase in the RBD levels, we did not find any difference in the levels of neutralizing antibody between the vaccinated and unvaccinated pregnant women. From the vaccination records, majority of the respondents had received only a single dose of the viral-vectored Oxford-AstraZeneca vaccine and these vaccinations for most of the women were before the current pregnancies. Thus, the number of doses and timing of these immunizations may impact the durability and function of vaccine-induced antibody responses.
Our study had some limitations as discussed below: First, the cross sectional study design did not allow us to follow pregnant women and infants up to determine durability of SARS-CoV-2 specific antibodies. Prior work has shown that vaccination in pregnant women induce more durable SARS-CoV-2 protective antibodies compared to infection-induced antibodies31. Secondly, in comparing the neutralizing antibodies for vaccinated and non-vaccinated pregnant women, we used a kit that has recently been shown to have reduced sensitivity to emerging variants particularly post-omicron era22,32. Molecular surveillance data from Ghana published after the present study was conducted showed that the predominant circulating strain during the study period was the omicron variant21. This may explain why the elevated anti-RBD antibodies seen in vaccinated mothers versus unvaccinated mothers did not translate to an increased percent of inhibition of ACE-2 binding in our study.
Thirdly, the absence of RT-PCR positive cases in our study population limited our ability to investigate fully, the effects of active prenatal COVID-19 on birth outcomes within our population. Additionally, due to the low vaccination rates among our participants, we were unable to compare transfer efficiencies between COVID-19 vaccinated and unvaccinated individuals. However, the efficient transfer of naturally acquired antibodies from mothers to neonates together with the strong correlation between maternal and infant antibody levels in our population supports the immunisation of pregnant women to ensure optimal protection of infants during the first few weeks of life.
In conclusion, we have shown higher SARS-CoV-2 antibody seropositivity in pregnant women in Accra. There is efficient transplacental transfer of SARS-CoV-2 antibodies comparable to malaria-specific antibodies in SARS-CoV-2 seropositive pregnancies. The study highlights the importance of maternal immunisation to enhance neonatal health.
Methods
Study participants
The study was a cross sectional prospective study conducted from March to May 2022 at the Maamobi General Hospital, Accra, Ghana. Pregnant women attending routine antenatal clinics (ANC) and parturient women admitted to the delivery wards were enrolled in the study. Pregnant women at ANC were sampled during the month of March 2022 while samples from parturient women were collected from March to May 2022. Naso- and oropharyngeal swabs were collected from all the pregnant women into virus transport media. Heparinized venous blood was drawn from participants at both ANC and delivery wards. At the delivery wards, matching cord blood was drawn immediately after clamping the cord. Plasma was separated by centrifugation and stored at -20°C until use. A structured questionnaire was administered to study participants to capture socio-demographic, clinical and vaccination status. In addition, participants were screened by the medical team for fever, cold, cough, loss of smell or taste, chest tightness and breathing difficulty within 48 h before enrolment. Anthropometric data including gestational age, birth weight, length of baby and head circumference were recorded after delivery and was used in assessing birth outcomes.
The study was approved by the Institutional Review Board of Noguchi Memorial Institute for Medical Research (NMIMR-IRB CPN 047/21-22) and the Ghana Health Service Ethics Review Committee (GHS-ERC 021/11/22). Participation in the study was voluntary and informed consent was obtained from all volunteers before taking part in the study. All methods were carried out in accordance with relevant guidelines and regulations.
RT-PCR for SARS-CoV-2 detection
Naso- and oropharyngeal swabs in virus transport medium were transported to the Virology lab at Noguchi Memorial Institute for Medical Research for RT-PCR detection of SARS-CoV-2. Viral ribonucleic acid (RNA) was extracted using the RADI PREP DNA/RNA kit (KH Medical) following the manufacturer's protocol. The extracted RNA was tested in RT-PCR using Veri-Q Prep M16 system (MiCoBiomed, Korea) with primer sets and probes targeting the open reading frame (ORF3a), envelope genes (E) and SARS-CoV-2nucleoprotein following the manufacturer’s protocol and has been described in detail previously33. Cycle threshold less than 40 was considered positive for SARS-CoV-2. All RT-PCR was run within 24 h after sample collection.
Protein expression
The SARS-CoV-2 nucleoprotein (N) was was expressed as detailed in34. Briefly, the protein was expressed from the plasmid pHYRSF53 (Addgene #64696), containing the SARS-CoV-2 nucleocapsid gene (NCBI Gene ID: 43740575) fused with a cleavable SUMO-His-tag nucleotide sequence in Escherichia coli BL21 (DE3) cells. After Ni-NTA purification, the SUMO-His-tag was cleaved and further purified using Ni-NTA beads to obtain a tag-free nucleocapsid antigen. The receptor binding domain (RBD) of the spike protein of the SARS-CoV-2 ancestral strain was expressed in 293 freestyle cell systems as described elsewhere35. Plasmodium falciparum circumsporozoite protein (CSP) and merozoite surface protein (MSP3) of the 3D7 clone were expressed in the Lactococcus lactis system and as described elsewhere in detail36.
Antigen-specific IgG reactivity
To measure antigen-specific IgG in plasma, an indirect ELISA was used as described previously37 with a few modifications. In summary, Nunc Maxisorp plates were coated with antigens at 0.5 µg/mL (N antigen), 0.5 µg/mL (RBD antigen), 1 µg/mL (CSP), 0.5 µg/mL (MSP3) and stored at 4°C overnight. Plates were washed with PBS/tween and blocked with BSA/PBS-Tween for 1 h. Following blocking, plasma was added to wells at a dilution of 1:100 for all antigens except MSP3 which was diluted at 1:200. Plasma pools from COVID-19 convalescent individuals and archived pre-pandemic samples from non-pregnant adults were used as positive and negative plasma control respectively to normalize plate-plate variation. After plasma incubation, plates were washed and IgG binding detected with horseradish peroxidase (HRP)-conjugated rabbit anti-human IgG. Substrate was developed with 3,3',5,5' tetramethylbenzidine (TMB) and absorbance was read at 450 nm. The cut-off for seropositivity was calculated as follows;
SARS-CoV-2 neutralizing antibody (NAbs) levels
To quantify neutralizing antibodies to SARS-CoV-2 virus in plasma, the WANTAI SARS-CoV-2 NAbs ELISA (Beijing Wantai Biological Pharmacy Enterprise; Beijing, China) was used according to manufacturer’s instructions. In summary, 100 μL of plasma and controls were added to SARS-CoV-2 spike pre-coated plates in duplicates and incubated for 60 min at 37 °C. After incubation, the plates were washed 5 × with supplied wash buffer. Following which, 100 μL of HRP-conjugated spike-RBD antibody was added. The plates were then incubated for 30 min at 37 °C and washed again 5x. Subsequently, the plates were developed by the addition of 50 μL of Chromogen Solution A and 50 μL of Chromogen Solution B per well and incubated in the dark for 15 min at 37 °C. The reaction was stopped with 50 μL of stop solution per well. The absorbance of 450 nm was measured using a Multiskan FC plate reader. Samples were analyzed in duplicates and the binding inhibition rate was calculated as:
Samples with a binding inhibition rate ≥ 50% were considered positive.
Data analysis
Data was analysed using Graph Pad Prism Software Version 10.0.0 (Boston, Massachusetts USA). Seropositivity rates were estimated as the proportion of individuals (and 95% confidence interval) reactive to SARS-CoV-2-specific antigens based on the seropositivity cut-off. Paired t- test was used to compare matching maternal and cord IgG levels. Transplacental efficiency was estimated as the ratio of cord IgG levels: maternal IgG levels. In all analysis, p value < 0.05 was considered statistically significant.
Data availability
All data generated or analysed during this study are included in this published article.
References
UN News. WHO chief declares end to COVID-19 as a global health emergency | UN News. United Nations News https://news.un.org/en/story/2023/05/1136367 (2023).
Kusi, K. A. et al. High infectious disease burden as a basis for the observed high frequency of asymptomatic SARS-CoV-2 infections in sub-Saharan Africa. AAS Open Res. 4, 2 (2021).
Kobia, F. & Gitaka, J. COVID-19: Are Africa’s diagnostic challenges blunting response effectiveness?. AAS Open Res. 3, 4 (2020).
Woodworth, K. R. et al. Birth and infant outcomes following laboratory-confirmed SARS-CoV-2 infection in pregnancy - SET-NET, 16 jurisdictions, March 29-October 14, 2020. MMWR Morb. Mortal. Wkly. Rep. 69, 1635–1640 (2020).
Villar, J. et al. Maternal and neonatal morbidity and mortality among pregnant women with and without COVID-19 infection: The INTERCOVID multinational cohort study. JAMA Pediatr. 175, 817–826 (2021).
Karasek, D. et al. The association of COVID-19 infection in pregnancy with preterm birth: A retrospective cohort study in California. Lancet Reg. Health Am. 2, 100027 (2021).
Sedgh, G., Singh, S. & Hussain, R. Intended and unintended pregnancies worldwide in 2012 and recent trends. Stud. Fam. Plann. 45, 301–314 (2014).
Usuf, E. & Roca, A. Seroprevalence surveys in sub-Saharan Africa: What do they tell us?. Lancet Glob. Health 9, e724–e725 (2021).
Mensah, B. A. et al. Population-based sero-epidemiological investigation of the dynamics of SARS-CoV-2 infections in the Greater Accra Region of Ghana. Sci. Rep. 12, 21582 (2022).
Janha, R. E. et al. SARS-CoV-2 seroprevalence in pregnant women during the first three COVID-19 waves in The Gambia. Int. J. Infect. Dis. 135, 109–117 (2023).
Koech, A. et al. SARS-CoV-2 seroprevalence in pregnant women in Kilifi, Kenya from March 2020 to March 2022. Front. Public Health https://doi.org/10.3389/fpubh.2023.1292932 (2023).
Sawry, S. et al. High prevalence of SARS-CoV-2 antibodies in pregnant women after the second wave of infections in the inner-city of Johannesburg, Gauteng Province, South Africa. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 125, 241–249 (2022).
Lucinde, R. K. et al. Sero-surveillance for IgG to SARS-CoV-2 at antenatal care clinics in three Kenyan referral hospitals: Repeated cross-sectional surveys 2020–21. PloS One 17, e0265478 (2022).
González, R. et al. SARS-CoV-2 seropositivity and HIV viral load among Mozambican pregnant women. J. Acquir. Immune Defic. Syndr. 1999(92), 115–121 (2023).
Hookham, L. et al. Seroepidemiology of COVID-19 in pregnant women and their infants in Uganda and Malawi across multiple waves 2020-2022. 2023.08.19.23294311. Preprint at https://doi.org/10.1101/2023.08.19.23294311 (2023).
Amaratunga, C. et al. A role for fetal hemoglobin and maternal immune IgG in infant resistance to Plasmodium falciparum Malaria. PLOS One 6, e14798 (2011).
Ray, J. E. et al. Reduced transplacental transfer of antimalarial antibodies in Kenyan HIV-exposed uninfected infants. Open Forum Infect. Dis. 6, ofz237 (2019).
Joseph, N. T. et al. Maternal antibody response, neutralizing potency, and placental antibody transfer after severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection. Obstet. Gynecol. 138, 189–197 (2021).
Edlow, A. G. et al. Assessment of maternal and neonatal SARS-CoV-2 viral load, transplacental antibody transfer, and placental pathology in pregnancies during the COVID-19 pandemic. JAMA Netw. Open 3, e2030455 (2020).
Assefa, N. et al. Seroprevalence of anti-SARS-CoV-2 antibodies in women attending antenatal care in eastern Ethiopia: A facility-based surveillance. BMJ Open 11, e055834 (2021).
Asante, I. A. et al. Repurposing an integrated national influenza platform for genomic surveillance of SARS-CoV-2 in Ghana: A molecular epidemiological analysis. Lancet Glob. Health 11, e1075–e1085 (2023).
Martínez-Barnetche, J. et al. Comparable diagnostic accuracy of SARS-CoV-2 Spike RBD and N-specific IgG tests to determine pre-vaccination nation-wide baseline seroprevalence in Mexico. Sci. Rep. 12, 18014 (2022).
Weisberg, S. P. et al. Distinct antibody responses to SARS-CoV-2 in children and adults across the COVID-19 clinical spectrum. Nat. Immunol. 22, 25–31 (2021).
Budhram, S. et al. Maternal characteristics and pregnancy outcomes of hospitalized pregnant women with SARS-CoV-2 infection in South Africa: An international network of obstetric survey systems-based cohort study. Int. J. Gynecol. Obstet. 155, 455–465 (2021).
Carrasco, I. et al. SARS-COV-2 infection in pregnant women and newborns in a Spanish cohort (GESNEO-COVID) during the first wave. BMC Pregnancy Childbirth 21, 326 (2021).
Rubio, R. et al. Maternal and neonatal immune response to SARS-CoV-2, IgG transplacental transfer and cytokine profile. Front. Immunol. https://doi.org/10.3389/fimmu.2022.999136 (2022).
Flannery, D. D. et al. Assessment of maternal and neonatal cord blood SARS-CoV-2 antibodies and placental transfer ratios. JAMA Pediatr. 175, 594–600 (2021).
Boelig, R. C. et al. Comprehensive serologic profile and specificity of maternal and neonatal cord blood SARS-CoV-2 antibodies. AJOG Glob. Rep. 2, 100046 (2022).
Helguera-Repetto, A. C. et al. Cord blood SARS-CoV-2 IgG antibodies and their association with maternal immunity and neonatal outcomes. Front. Pediatr. https://doi.org/10.3389/fped.2022.883185 (2022).
Partey, D. et al. COVID-19 Risk perception and vaccine uptake among pregnant women in Ghana. https://doi.org/10.21203/rs.3.rs-3585205/v1 (2023).
Otero, S. et al. Maternal antibody response and transplacental transfer following severe acute respiratory syndrome coronavirus 2 infection or vaccination in pregnancy. Clin. Infect. Dis. 76, 220–228 (2023).
Bayart, J.-L. et al. Analytical sensitivity of six SARS-CoV-2 rapid antigen tests for omicron versus delta variant. Viruses 14, 654 (2022).
Akowuah, K. A. et al. SARS-CoV-2 infections among asymptomatic individuals contributed to COVID-19 cases: A cross-sectional study among prospective air travelers from Ghana. Front. Public Health 10, 1035763 (2022).
Tapela, K. et al. Development of an Affordable ELISA Targeting the SARS-CoV-2 Nucleocapsid and Its Application to Samples from the Ongoing COVID-19 Epidemic in Ghana. Mol Diagn Ther 27, 583–592 (2023).
Wrapp, D. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260–1263 (2020).
Singh, S. K. et al. Lactococcus lactis provides an efficient platform for production of disulfide-rich recombinant proteins from Plasmodium falciparum. Microb. Cell Factories 17, 55 (2018).
Partey, F. D. et al. Kinetics of antibody responses to PfRH5-complex antigens in Ghanaian children with Plasmodium falciparum malaria. PLOS ONE 13, e0198371 (2018).
Acknowledgements
We are grateful to the pregnant women who volunteered to participate in the study for their time and effort and the midwives at the Maamobi General hospital. We thank Professor Michael Theisen (Staten’s Serum Institute, Denmark) for the generous gifts of CSP and MSP3 antigens. This work was supported by the Noguchi Memorial Institute for Medical Research through the Office for Research Support Fund (Fund ID- EC/P26965/01). F.D.P, DO. B.A.M and E.Y.B were grant recipients.
Funding
Funding was provided by Noguchi Memorial Institute for Medical Research (Grant number: EC/P26965/01).
Author information
Authors and Affiliations
Contributions
F.D.P: conception, design of the work; acquisition, analysis, interpretation of data; writing—original draft, review & editing, funding acquisition. D.O: conception, design of the work; acquisition, analysis, interpretation of data; writing—original draft, review & editing, funding acquisition. E.Y.B.: conception, design of the work; review & editing, funding acquisition. A.N.A.P: acquisition, analysis, interpretation of data; writing—review & editing. I.F.D: acquisition, analysis, interpretation of data; writing—review & editing. J.A: acquisition, analysis, interpretation of data; writing—review & editing. K.E: acquisition, analysis, interpretation of data; writing—review & editing. F.Y.N: acquisition, analysis, interpretation of data; writing—review & editing. K.A.K: acquisition, writing—review & editing. B.A.M.: conception, design of the work; acquisition, analysis, interpretation of data; review & editing, funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Partey, F.D., Obiri, D., Bonney, E.Y. et al. Efficient transplacental transfer of SARS-CoV-2 antibodies between naturally exposed mothers and infants in Accra, Ghana. Sci Rep 14, 10772 (2024). https://doi.org/10.1038/s41598-024-61496-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-61496-3
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
