
Abstract
Fertilizers application are widely used to get a higher yield in agricultural fields. Nutrient management can be improved by cultivating leguminous species in order to obtain a better understanding of the mechanisms that increase the amount of available phosphorus (P) and potassium (K) through fertilizer treatments. A pot experiment was conducted to identify the leguminous species (i.e., chickpea and pea) under various fertilizer treatments. Experimental design is as follows: T0 (control: no fertilizer was applied), T1: P applied at the level of (90 kg ha−1), T2: (K applied at the level of 90 kg ha−1), and T3: (PK applied both at 90 kg ha−1). All fertilizer treatments significantly (p < 0.05) improved the nutrient accumulation abilities and enzymes activities. The T3 treatment showed highest N uptake in chickpea was 37.0%, compared to T0. While T3 developed greater N uptake in pea by 151.4% than the control. However, T3 treatment also increased microbial biomass phosphorus in both species i.e., 95.7% and 81.5% in chickpeas and peas, respectively, compared to T0 treatment. In chickpeas, T1 treatment stimulated NAGase activities by 52.4%, and T2 developed URase activities by 50.1% higher than control. In contrast, T3 treatment enhanced both BGase and Phase enzyme activities, i.e., 55.8% and 33.9%, respectively, compared to the T0 treatment. Only the T3 treatment improved the activities of enzymes in the pea species (i.e., BGase was 149.7%, URase was 111.9%, Phase was 81.1%, and NAGase was 70.0%) compared to the control. Therefore, adding combined P and K fertilizer applications to the soil can increase the activity of enzymes in both legume species, and changes in microbial biomass P and soil nutrient availability make it easier for plants to uptake the nutrients.
Similar content being viewed by others
Introduction
Nutrient management is an important strategy for achieving high plant yield and maintaining soil fertility status1. Nitrogen (N) Phosphorus (P) and potassium (K) are the primary macronutrients that regulate the plant growth and development2. These essential nutrients play a significant role in physiological processes, such as N, which is directly related to the photosynthesis process, which is important for the healthy vegetative growth of a plant. Phosphorus and K are involved in protein synthesis, enzyme activation, glycolysis, and sugar transport redox reactions3,4. Macronutrient deficiencies in agriculture are widespread, affecting plant production around the world5. Phosphorus and K fertilizers will continue to play a vital role in agricultural systems. It turns out that any disturbance, including fertilization, crop rotation, tillage, or pollution, primarily affects this organic layer of soil and affects the microbial communities6,7. Pakistan is situated in an arid to semi-arid area of the world, and deficiency of P is mainly noted in soils that have lower moisture contents and dry lands8. About 80–90% of Pakistani soils are P-deficient due to their high carbonate content (CaCO3 > 3.0%) and alkaline pH > 7.0. There is a need to improve its range to supply external P content9.
The application of fertilizer could increase several processes; the first would modify the properties of the soil10. However, diminishing natural resources in P and K are a serious concern due to reduced availability and increased costs11,12. Fertilizer application, tillage practices, and other conventional agricultural methods alter soil properties, requiring the improvement and examination of soil quality13,14. Therefore, the application of extensive fertilizer not only increases the P fixation capacity but also causes environmental pollution. In advance agriculture, inorganic and organic fertilizer, or a combination of both, were used to get higher production. On the other hand, biological processes have significance for the functioning of agricultural systems15. Soil microbes regulate P cycling through organic phosphate mobilization and the breakdown of organic matter. Soil microbial biomass, also known as microbiological biomass P (MBP), can accumulate released P. Soil microbes and plants may benefit from MBP's multiple functions as a P sink and the accumulation of potentially bioavailable P. Microbes in the soil compete with plants for plant-available P16. Green manures are a fast-growing crop that is one of the most effective methods for improving soil quality in a variety of ways. They have the ability to enhance soil quality, organic matter, and improve nutrient availability, such as P and K. In this regard, the present study cultivates leguminous species to find a better understanding of the mechanisms that improve plant nutrients and fertilizer treatments. Leguminous green manure crops have the potential to improve the nitrogen cycle of the soil and highly absorb nutrients from the soil, and high nutrient uptake capacity is mainly related to plant root morphological traits to achieve high yields under low nutrient conditions because a plant may use different strategies to take up P and K from low-nutrient soils17. Leguminous plants can produce pod-shaped fruits that contain fleshy seeds that are known as beans.
In Pakistan, peas and chickpeas are important food legume crops. These are significant nutritional legume crops that provide a low-cost, necessary supply of protein. Chickpeas are a valuable source of vitamins, carbohydrates, and minerals. Both chickpeas and peas are multi-functional legume crops that play an important role in small-scale economics for farmers18. Leguminous plants can easily acquire greater amounts of P and K from the soil for their growth and the associated root morphological mechanisms19,20. Plant root characteristics can improve soil aggregates, decrease soil bulk density, and maintain soil pH; these factors can possibly increase soil quality21. The root systems of leguminous species are mainly occupied by a large number of microorganisms that also have the ability to release enzymes, which contribute significantly to the various processes that are associated with plant growth22,23. Soil microorganisms perform the basic role in soil organic P transformation, and microbial biomass is also essential for nutrient cycling in soil status24. It is assumed that the microbial biomass P is directly released from cells when microbes die, is certainly decomposed in soil, and can be easily taken by plants12,25. Soil enzymatic activities are an indicator of microbial activity and function. They can modify soil chemical processes, and soil organic matter (SOM) dynamics contribute to changes in abiotic and biotic factors in soil26,27. In order to examine the soil microbial biomass,28 reported that fertilizer treatments also had an impact on a number of enzyme activities29. Each enzyme plays a specific role in increasing nutrient availability, while some enzymes related to the carbon cycle (C), for example, β-xylosidase and β-glucosidase are famous for their quick responses30. However, urease (URase) and N-acetylglucosaminidase (NAGase) enzymes that regulate the N cycle hydrolyze urea into ammonia and carbon dioxide, and NAGase breakdown amino acids into sugar, which are the main sources of N mineralization31. Some enzymes are famous for their quick responses, such as β-xylosidase and β-glucosidase. These enzymes break down complex molecules into simple ones for plant uptake32. It was previously reported that phosphatase activity catalyzes the cleavage of P minerals from organic phosphates in acidic and alkaline soils and enhances organic matter with the help of mycorrhizal species33,34.
An estimation of the amount of soil available as nutrients for crops would help in the design of fertilizer application regimes to optimize the P and K fertilization rates. By measuring the amount of P and K present in the soil, it is possible to estimate the fertilizer application rate35,36,37. Many researchers have been emphasizing the N fertilizer treatment to evaluate the leguminous species, further demonstrating the activities of enzymes affected by N fertilization38,39. An earlier study used mineral fertilizer applications with manures and N fertilizer applications to investigate the responses of enzymatic activities to legume crops40. Keeping this view in mind, the present research focuses on appropriate P and K fertilizer strategies and responses to improve nutrient uptake and nutrient use efficiency in legume species and their impact on soil nutrient availability. In addition, there is limited research on how soil enzymes and microbial biomass phosphorus vary in response to the cultivation of leguminous species under different P and K fertilizer treatments. It is hypothesized that specific fertilizer treatments could potentially improve the mechanisms involved in plant nutrient uptake and plant nutrient use efficiency subsequently increasing the dry biomass yield of leguminous crops. This study aims to investigate the responses of soil enzymes (such as β-glucosidase phosphatase, Urease, and N-acetylglucosaminidase) and microbial biomass phosphorus to variations in PK fertilization treatments. Further, explore how these responses showed a correlation with the nutrient uptake abilities of legume crops.
Materials and methodology
Experimental design
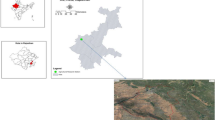
The pot experiment was carried out in Sindh province, located between coordinates 25°42 34″ N and 68°54′ 08″ E in southern Pakistan, to determine the responses of two different legume species under P and K fertilizer techniques. We collected the experimental soil from the field of the Pulses Research Sub-station, Tandojam Sindh, using an auger with an internal diameter of 5 cm and a depth of 0–20 cm. The basic soil properties are the following: soil organic matter (SOM) 0.65%; total N (TN) 1.5 g kg−1; available P 10.8 mg kg−1 and K 112.3 mg kg−1; soil pH 7.8 (1:2.5), EC was 0.35 cmol kg−1 and CaCO3 was 8.3% with clay loam texture class. While the seeds of legume species were collected from the Agriculture Research Institute in Tandojam. An experiment design was planned as a completely randomized design with factorial arrangements; each treatment had four replications, and a total of 16 pots were used for each legume species. Every pot was manually packed with 7 kg of dry soil with upper 28 cm diameters and lower 18 cm and a depth of 26 cm with a surface area of 0.049 m2. However, each pot was properly homogenized with P (P2O5), and K (K2O) fertilizers.
The fertilizer sources used in the experiment were superphosphate (P2O5, 12%) and potassium chloride (K2O, 60%). Treatments fallow as T0 (no fertilizer was applied (control), T1 (applied P at a level of 90 kg ha−1), T2 (applied K at the level of 90 kg ha−1), and T3 (applied both PK together at the level of 90 kg ha−1), respectively. The leguminous species seed was soaked in water for one day before sowing. After the preparation of soil for the pot experiment, ten seeds were sown in each pot and covered with dry soil. After plant development, five vigorous plants in every pot allowed to be examined were selected. While distilled water was commonly used for irrigation purposes. Leguminous species Chickpeas (Cicer arietinum L) and Peas (Cicer arietinum L), fallowed as cultivars (NIAB Channa-2016) and (Sarsabz) were seeded on 1st November 2020. Figure 1 shows the geographical location of the surveyed region.

Location map of study area.
Plant sampling and analysis
At the full bloom stage, on 5th March 2021 both leguminous species were harvested, and the plant shoot samples were separated from the pots. After that, plant root samples were removed from the pots and washed thoroughly to remove any remaining soil. Leguminous fresh shoot and roots biomass samples were weighed, dried for 72 h at 65 °C in an oven, crushed, and stored for nutrient analyses. Samples of plant shoots and root were burned at high temperatures to determine the N, P, and K contents using a solution of sulphuric acid (H2SO4) and hydrogen peroxide (H2O2) diluted. Further, a previously described Kjeldahl digestion method, was used to assess the amount of N in plant shoot and root biomass; the molybdovanadate method was used to determine the amount of P in plants41, while determination of K content in plants flame photometer was used to analysis42.
Soil determination
Following plant harvesting, soil samples were collected from each pot, packed, and then transported to the laboratory for analysis. After that, the samples were separated into two main parts, and immediately kept one portion at − 4 °C for soil moisture content, inorganic nitrogen, and enzyme analysis.
The second portion of samples was air-dried and passed through a 2-mm sieve for the determination of pH range, available P, K contents. Additionally, we passed the sub-samples through a 0.25-mm sieve to examine soil total nitrogen (TN) and SOM. Analysis of soil inorganic nitrogen contents by the Kjeldhal method of steam distillation, as described in an earlier study43. Oven drying at 105 °C for 48 h determined soil moisture. Soil TN content was examined by the Kjeldhal method described in previous research44. An earlier Walkley–Black method, was used for the determination of SOM44. Soil electrical conductivity (EC) and pH were determined by preparing a ratio of 1:2.5 soil/water in order to use an EC meter (Hanna Model-8733, Germany) and a pH meter (Jenway, Model-3510, Gransmore Green, Felsted, Dunmow, Essex, CM6 3LB, UK), respectively. According to45,46, microbial biomass phosphorus (MBP) was examined in fresh, wet soil using a chloroform-fumigation extraction method. To determine the total P in the soil, samples are digested in a solution of perchloric acid (HClO4) and nitric acid (HNO3) in a ratio of 1:3. According to the procedure described in47, a 0.5 M NaHCO3 extract solution was used to determine soil Olsen P. The contents of total P and available P were analyzed using a spectrophotometer with visible blue light (Model UV-2100, Shimadzu, Tokyo, Japan). For available K content, 5 g of dry soil was extracted with one molarity of NH4OAc and determined on a flame photometer48.
Soil enzyme activities
All enzyme activities were determined in fresh soil samples collected after harvesting the legume species. Before the enzyme activity assay, a portion of the subsamples were oven dried at 105 °C for 24 h to determine soil moisture. The activities of the urease enzyme were investigated by the previously described colorimetric method49. Firstly, 5 g of fresh soil was incubated for 2 h at 36 °C, and urea solution was used as a substrate. An altered Berthelot reaction was used to release NH4+ through the potassium chloride (KCl) solution. The β-glucosidase activity was assessed in 1 g of moist soil (< 2 mm) kept in a tube, treated with 0.25 ml of toluene, and 4 ml of adjusted universal buffer (pH 6), added 1 ml of PNG solution (25 mM) and placed in an incubator for 1 h at 37 °C50. After incubation, 1 ml of CaCl2 solution and 4 ml of Tris buffer (pH 12) were added, and absorbance at 400 nm was measured using a spectrophotometer (RIGOL, USA). As described by51. Phosphatase activity has been investigated using the substrate p-nitrophenyl phosphate (r-NPP). After that, 5 g of wet soil was mixed with 20 ml of acetate buffer (pH 5.2) and 100 mMof r-NPP and kept for 30 min in an incubator at 30 °C. To determinate the reaction, 1 ml of CaCl2 and 4 ml of 0.2 MNaOH were added after incubation. The absorbance was measured with a spectrophotometer set to 405 nm (RIGOL, United States). The p-nitrophenol produced method after the soil was incubated with added substrate and acetic buffer (pH 5.5) for an hour at 37 °C was used to evaluate the N-acetyl-D-glucosaminidase enzyme. At 400 nm, the filtrate's intensity of its yellow colour was measured52. The activities of enzyme units have been calculated in Table 1.
Statistical analyses
IBM SPSS Statistics version 20.0 (Corp., Armonk, NY, USA) was used to evaluate a one-way analysis of variance (ANOVA) to see if there were any significant effects of fertilizer treatments on legume nutrient accumulation. Investigate the fertilizer application by using Tukey’s multiple variable levels (p < 0.05). The significant correlation pattern of the study has been examined using IBM SPSS by applying Pearson’s correlation and indicating the significant range at ** p < 0.01 and * p < 0.05. Origin software, using Origin Pro 9.0, created the graphs for the enzymatic activity determination. The principal component analysis (PCA) was performed to determine the general relationship between various cultivars, enzymatic activities, and soil characteristics using CONOCO (version 5) at a p < 0.05 level.
Ethical approval and consent to participate
This study does not include human or animal subjects.
Results
Shoot biomass yield
Application of fertilizers has significant effects on chickpea and pea shoot dry biomass yield and root biomass (Table 2). Chickpeas presented the highest dry shoot biomass yield in T3 treatment by 33.8% as compared with T0 treatment. A lower amount of biomass yield increased in T1 treatments by 12.0% compared to T0 treatments. The T3 treatment recorded the highest root biomass of chickpeas at 21.2%, while the T2 treatment recorded the lowest root biomass at 12.1%, compared to the control. However, chickpea species decreased root biomass in T1 by 48.1% compared to the control. In the case of pea species, the maximum shoot dry biomass yield in the T1 treatment increased by 19.9% compared to the control. Despite that, T3 treatment also increased the dry biomass yield of pea species by 15.3%, higher than control. Root dry biomass and nutrient uptake capability of pea species showed significant changes under fertilization treatments. The pea species root dry biomass increased in the following pattern: T3, and T2, by 148.9% and 65.9%, respectively, compared to the T0 treatment.
Chickpea shoot nutrient uptake
Significant changes were observed in shoot nutrient uptake during different fertilizer treatments, as presented in Table 3. However, T1 treatment promoted the N uptake ability by 37.0%, compared to the control. While the minimum shoot N uptake (11.0%) was noted in T2 treatment compared to the T0. Whereas P and K uptake of chickpeas increased significantly in T2, i.e., 52.2% and 44.2%, respectively, compared to the control. The least shoot P uptake in chickpea was noted in T1 by 18.1% compared to the control. Shoot K uptake of chickpea was decreased in T3 treatment, which was 7.7% compared to T0.
Chickpea root nutrient uptake
Table 4 showed the variation in root nutrient (N, P, and K) uptake ability of chickpea species during fertilization. Therefore, compared to the control, chickpea species reduced their N uptake abilities under fertilizer treatments. In contrast, fertilizers treatment enhanced root P and K uptake, the maximum P and K uptake of root was recorded in the T3 treatment, i.e., 26.2 and 13.8%, respectively, as compare with T0 treatment. The minimum root P and K uptakes of chickpea cultivars were found in T2 at 15.6% and 8.9%, respectively, compared to no fertilizer applied.
Pea shoot nutrient uptake
Fertilizer treatments have significant effects on nutrients such as N, P, and K uptake in pea shoots as presented in (Table 5). The greater N uptake ability was observed in T3 by 151.4% compared to the control. The lowest N uptake was recorded for T1 treatment by 6.5% compared to the control. On the other hand, pea species had a high amount of P uptake in their shoot under T3 (49.2%), and the minimum shoot P uptake was detected in the T1 by 27.2% compared to the T0 treatment. The maximum K uptake ability was noted in T1 (50.3%), compared to the control. While shoot K uptake was reduced in T3 treatment in comparison with control.
Pea root nutrient uptake
Significant modifications were recorded in the nutrient uptake of pea species roots during fertilizer treatments (see Table 6). The maximum root N uptake increased in T3 by 151.4%, while root N uptake decreased by 32.3% in T1 compared to T0 treatment. In comparison to the control, the T3 treatment increased root P uptake by 100.1%, and the lower root P uptake was 40.0% observed in the T1 treatment. However, the greater root K uptake of pea species was noted in the T3 treatment by 13.0%, and the root K uptake was reduced in the T1 treatment by 3.1% compared to the T0 treatment.
Soil nitrogen contents
The soil mineral nitrogen content affected by P and K fertilizer treatments during the planation of legume species is shown in Table 7. The T2 treatment increased the NH4+ and NO3- content, i.e., 15.3% and 16.6% in chickpea species respectively, higher than control. While the T3 treatment reduced the NH4+ content by 2% and T1 decreased the NO3− content by 15% in chickpeas as compared to the T0 treatment. In the case of pea species, the T2 improved the NH4+ and NO3− content of the soil, i.e., 21.1% and 34.4%, respectively, compared to the control.
Soil MBP and available P and K contents
Soil microbial biomass phosphorus (MBP) and available P and K contents of soil changed with fertilizer treatments when leguminous species were grown (Table 8). The percentages of MBP in T3 (95.7%), T1 (90.3%), and T2 (79.2%) in chickpea species were greater than control. The Olsen P of chickpea species was increased in T2 and T3, i.e., 36.3% and 28.9%, respectively, compared to T0 treatments. While the P content of soil was decreased in T1 treatment by 32.9% compared to control. Soil available K content showed an increasing trend in the following directions: 16.7%, 32.8%, and 42.9% for T1, T2, and T3, respectively, compared to T0 treatment. In the case of pea species, T3 treatment increased the highest range of MBP by 81.5%, and the T1 recorded the lowest MBP at 9.31%, compared to the T0 treatment. The maximum soil available P content observed in T2 was 34.8%, and T3 also improved P content by 31.0% as compared to T treatment. The greater K content of soil was in T3 by 28.6%, when compared with control. Soil available K content decreased in T1 by 5.2% compared to control.
Influences of fertilization treatments on Enzymes activities
Fertilizer treatments showed a significant effect on soil enzymatic (i.e., N-acetyl-β-D-glucosaminidase (NAGase), β-glucosidase (BGase), phosphates (Phase), and urease URase) activities after harvesting the chickpea species, as shown in Fig. 2. T1 showed the highest NAG activity at 52.4%, while T2 and T3 also increased, i.e., 21.6% and 15.5% activity, respectively, in chickpeas compared to T0. However, both BGase and Phases enzyme activities were higher in T3, i.e., 55.8% and 33.9%, respectively, as compared to the control. While T2 treatment improved soil URase enzyme activities by 50.1% compared to the control.

Effects of fertilization on soil enzyme activities after harvesting the chickpea species: (a) N-acetylglucosaminidase (NAGase), (b) β-glucosidase (BGase), (c) phosphatase (Phase), and (d) urease (URase). Bars with different small letters indicate significant variation at the (p < 0.05) level based on Tukey’s multiple test (repeats n = 4).
All fertilizer treatments enhanced the activity of extracellular enzymes in soil, such as NAGase, BGase, Phase, and URase after the harvesting of pea species (Fig. 3). Maximum NAGase activities were found in T3 treatments (70.0%), and the lowest activity recorded in T2 was 9.6% as compared to the T0 treatment. The T3 treatment significantly increased BGase enzyme activities by 149.7% compared to the control. However, the maximum Phase and URease activities were also prominent in T3, i.e., 81.1% and 111.9%, respectively, which is higher than in T0 treatment.

Impact of fertilizer treatments on soil enzyme activities after harvesting the pea species: (a) N-acetylglucosaminidase (NAG), (b) β-glucosidase (BG), (c), phosphatase (Phase), and (d) urease (Urease). Bars with different small letters indicate significant variation at the (p < 0.05) level based on Tukey’s multiple test (repeats n = 4).
Correlation between soil enzymes and soil properties
However, analysis of Pearson’s correlation (r) shows soil enzymes NAGase, BGase, Phase, and URase activities are significantly correlated with soil properties after harvesting the chickpea species during fertilization (Table 9). The NAG was significantly positively correlated with SOM and MBP (r = 0.502* and 0.543*, respectively). While the BG enzyme found a significant positive correlation with TN and AK (r = 0.517*, and 0.613*).The Phase activities indicated a strong and significant relationship with MBP and AK (r = 0.508*, 0.648**). However, the URase activities, also significantly related to MBP (r = 0.770**) and AK (r = 0.779**).
Based on Pearson’s correlation (r) analysis, the soil enzymes such as NAGase, BGase, Phase, and URase activities are considerably correlated with soil properties once harvested the pea species under PK fertilization (Table 10). The NAG enzyme was significantly positively related to SOM and K content (r = 0.505* and 0.611**, respectively), and BG showed a significant negative interaction only with soil TN (r = 0.517*). However, phase enzymes showed a significant negative relationship with MBP and AK, r = − 0.831** and − 0.822**, respectively. In contrast, a positive relationship is presented with AP r = 0.498*. Urease is significantly positively related to MBP and AP (r = 0.743**, 0.498*), and a negative relationship is seen with available K (r = − 0.686**). Non-significant associations were observed between soil enzymatic activities and other soil properties.
Correlation among legume nutrient accumulation, soil properties, and enzymatic activities
Based on principle component analyses, an overall correlation was seen among shoot and root PK uptake, soil properties, and enzymatic activities under PK fertilizer treatments (Fig. 4). During plantation chickpea cultivation, the first axis attributed 24.23% and the second axis 42.2% of the changes in plant PK uptake abilities, soil properties, and soil enzymes for chickpea species. T1 and T0 treatments clustered with chickpea root N, P, and uptakes, soil TN and soil pH. Soil characteristics such as available P and K content, MBP, and soil enzymes, Phase and URase presented a significant relationship followed by T3 treatment. While T2 cluster with shoot P uptake and NH4+and NO3–N.., The first and second axes contributed 25.2% and 46.2%, respectively. An opposite trend was noted among treatments, soil enzymes, and properties for pea species The results of the PCA analysis showed various fertilizer treatments presented interactions with PK uptake, soil properties, and soil enzymes. The T2 treatment linked soil properties (NO3 − , available P) and soil enzymes (phase and urease). Soil available K content, MBP and BGase enzymes, and root N uptake were all grouped together in the T3 treatments. While T0 and T1 treatments clustered with shoot and root N, P, and K uptake and soil properties, i.e., SOM, pH, TN, and NH4+., only NAGase soil enzymes were related to the T0 treatment.

PCA analysis showed an overall relationship between shoot and root PK uptake of plants, soil enzymes, and soil properties in two different legumes: (A) Chickpeas and (B) Peas. The position of variables showed relationships to each other. Note NAGase (N-acetylglucosaminidase), BGase (β-glucosidase), Phase (phosphatase), URase (urease) properties, TN (total nitrogen), NH4+ (ammonia) and NO3− (nitrate) SOM (soil organic matter), and AP, AK (available phosphorus and potassium).
Discussion
Effects of fertilizer on plant nutrient abilities
This study investigated the shoot and root biomass and phosphorus and potassium uptake abilities of two leguminous species, such as chickpea and pea species. The accumulation of biomass yield and the ability to acquire nutrients such as N, P, and K from the soil depend on fertilization techniques. However, in this study, fertilizer treatments increased biomass production and N, P, and K uptake abilities in both legumes. The PK fertilizer effects of N-uptake legumes are famous for their root nodule nitrogen fixation ability and nitrogen transfer capacity to the soil, which decrease competition with crops for nutrient uptake53. There is an extensive amount of researches are present about the quantity of nitrogen that legumes transfer to related crops. It fluctuates according to agronomic parameters like weather patterns and symbiotic effectiveness as well as conditions that affect legume N-fixation, such as species of legumes. In general, because the soil’s available nutrients change rapidly in the soil, they will be affected by many factors, including farming methods, fertilizer types, fertilizer application methods, and crop growth54. The previous finding showed that fertilizer input can improve plant biomass yield and is essential for enhancing agronomic efficiency, while a higher rate of fertilizer can have an adverse effect on plant growth55. Studies from the past showed that the levels of P and K in soils that had been treated with fertilizer were much higher. Another study56 demonstrated that legumes take up K from K-fertilized soils57. Phosphorus and K fertilization can improve nutrient uptake for legume crop growth. Plants with higher P and K uptake from the soil develop utilization in processes that promote quicker development and supply higher nutrient percentages to the above-ground plant parts58. Similarities were seen in other studies; for example, the P fertilizer is a parameter that reflects the plant nutrient (P) uptake capacity from the rhizosphere soil59. The excess P may accumulate in the inorganic and organic reservoirs, resulting in highly saturated PO43− pools in the soil. Phosphorus uptake does not depend on the present soil P content but is relative to the available (Olsen) P that plants may use60. The uptake of nutrients such as P and K during plant growth and biomass production depends on the root length, density, and root depth of a plant during its growing period61.
Impact of fertilization on soil nutrients
Our results showed significant differences in soil MBP under fertilization treatments. Generally, variations in soil MBP may depend on the soil amendment. Previous research supports the current study; results showed that oats were grown as green manure with chemical fertilizer applications such as N, P, and K and without organic fertilizer. The soil biomass P might have improved without the incorporation of organic fertilizer. This may happen because some limiting factors can help to easily decompose accessible P in soil, which was required to enhance biomass P in the soil. This development of microbial biomass P could increase when only inorganic fertilizer was applied62. During the P fertilization, when plant roots and microbes interacted with each other, microorganisms could release organic acids, phosphatases, enzymes, and other substances to stimulate soil insoluble phosphorus in the form of microbial biomass phosphorus, thereby improving soil phosphorus bioavailability46. In the present study, fertilization strategies could improve soil P and K contents. Similarly, it has been demonstrated that soil available P and K levels increased after receiving PK fertilization and were higher than in the non-P and K applied plots and other treatments57. Multiple studies have proposed that chemical fertilizer can improve soil physical and chemical properties as well as increase soil Olsen-P. Because the soil’s available nutrients change quickly in the soil, they will be affected by many factors, including farming methods, fertilizer types, fertilizer methods, and plant development37,63.
The current study showed T2 treatments where K fertilizer applied were more effective for soil P content. However,34 say that increasing the amount of P in the soil may involve a number of reactions, such as the pH of the soil, its sorption capacity, and the root exudation of legume species. Also, the properties of the soil affect how much P is available. In calcareous soils, P treatment often makes P less available because Ca-P minerals form and stick to the soil. The immobilized P will be released slowly; therefore, the released P comes from physico-chemical adsorption reactions. If the immobilized phosphorus is passed from one generation of microbes to the next, then there will be constant competition between microbes and plants for phosphorus64. This implies that the slow release of immobilized phosphorus contributes to the soil's available phosphorus. Measuring soil characteristics is crucial when fertilizer is extremely concentrated, as P and K fertilizer sources play a significant role When fertilizer is used, the K level in the soil goes up because some of the K in the fertilizer is fixed as non-exchangeable K and then slowly released to the soil in the form of available K content65.
Soil extracellular enzymatic activities
Plants and microbes can produce enzymes, which play an extensive role in increasing macronutrients in the soil66. Our results have indicated that application of P and K fertilizer affect enzyme activities during the cultivation of legume crops (Figs. 2, 3). The previous finding supports the current study, which demonstrates that the activities of enzymes showed increasing trends in response to applied chemical fertilization in soil67. The (T1) P fertilizer increased NAGase in chickpea species, while theT3 treatment improved NAGase activity in pea species. The NAGase enzyme activity is important for plant development and plays an important part in the N cycle, and the breakdown of amino acids into sugars can mineralize N in soil. On the other hand, excessive amounts of chemical fertilizer decreased soil enzyme activities68.
This study found that BGase and Phase activities increased when fertilizer was applied. According to an earlier study, BGase activities developed with inorganic fertilization treatment are commonly known as a sensitive factor in soil fertility and quality69,70. The PK addition resulted in a considerable increase in soil respiration which suggests that higher microbial activity in the presence of addition P allows for faster transformation of soil organic matter. Previous research indicates improved acid phosphate (AP) activity in the soils under fertilizer application71. Phosphatase activities are a key indicator of the soil's P cycle, and more activity can lead to a rise in the number of microorganisms and change the soil's properties30. The resulting P and K addition could proliferate the required P in the soil and restrict the availability of P, which can increase and maximize soil microbes’ activity. Soil microorganisms capable of secretion release large amounts of phosphorus activity into the soil31. Additionally, the results of the experiments showed that P and K treatments upgraded maximum urease activity. Different soil characteristics, such as soil pH, nutrient availability in the soil, and fertilizers, have an impact on urease enzyme activity72. Urease can regulate N availability for plant growth and could hydrolyze urea into ammonia and CO273.
Interaction between plant nutrient enzymatic activities and soil properties
The current research examines the enzymatic activities of soil, which can have integrated effects on soil properties under PK fertilizer treatments. These results could support a part of the current study's hypothesis that soil enzymes and soil microbial biomass P can increase nutrient availability. The Pearson correlation analysis showed significant relationships between enzyme activities, and soil properties under fertilization when two legumes were planted. However, PCA analysis showed an overall relationship between legume nutrient uptake and soil characteristics, MBP, and soil enzymes. While PCA analysis indicated a 24% and 25% total modification in the uptake of N, P and K, enzymes, and soil properties for both species after fertilization. The PCA graph showed that there was an interaction between nutrient uptake and MBP. Although there has been competition between plants and microbes, the presence of increases in soil microbial biomass phosphorus (MBP) could enhance the uptake of phosphorus by plants74. The MBP pool provides an important supply of available phosphorus (P) in soils. Even when Olsen P levels are high, the MBP pool continues to provide an adequate quantity of inorganic P, which plants can utilize more efficiently. However, the NAGase and urease N-related enzymes are positively related to SOM and MBP while also showing a significant negative interaction with TN during legume cultivation. Similarly, an earlier study demonstrates that legume species, such as alfalfa, is able to participate in N-cycling due to their atmospheric N-fixing capacity through interactions with Rhizobia bacteria75. Previously, it was observed that NAGase was essential for N mineralization. There may be a strong negative interaction because there are higher amounts of NH4+ and NO3− and the mineral N stops the production of the NAG67,76. However, NAGase, Phase, and urease also showed a significant relationship with MBP. Enzymatic activities in the soil may act as an indicator for the presence of organic substances. Enzymatic processes that interact with the community of microbes have the potential to break down SOM and improve nutrient availability. BG enzyme activities have a significant and also show a significant negative relationship with TN. The correspondence with soil C was strongest for the N-acquire enzyme (NAG) and lower for C-acquire enzymes. Applying fertilizer can change the microbial population system by greatly increasing the number of absular mycorrhizal fungi. These fungi take carbon from the host plants and provide inorganic nutrients to the plants.
Table 9 demonstrates that the planting of chickpea cultivars did not significantly interact with Olsen P and Phase activity, but there was a relationship between soil P and MBP. A greater amount of soil MBP leads to an increase in the available P content of the soil; hence, soil microbes’ microorganisms efficiently uptake orthophosphate from the soil solution, and P can also be secreted from the pool of MBP due to microbial turnover77. According to78 both factors exhibited major negative interactions with each other in their studies where P fertilizers were applied and in naturally fertile soil. Positive associations between soil P and phase activities play a great role in P cycling; however, the relations among soil P and phase activities are more complex, which is not seen in the present. A massive group of soil microbes that can solubilize organic P and microbial biomass is a vital factor for measuring the activity of the enzyme in an agricultural environment, which is mainly related to increased metabolic activities79.
There was a strong relationship between phase enzyme and soil P and a significant negative relationship between MBP and soil K during pea plantations. Previous research supports the current study by showing that soil respiration developed after long-term P addition, suggesting that microbial activities are higher with the occurrence of P addition, which allows quick transformation of the soil available. It is observed that phosphate activities are essential for soil P metabolism, P legume species from root secretions, and plant P enhancement as well80. It is considered that P and K fertilizer amendments can alter the microbial community, and influences on the relative abundance of abuscular mycorrhizae fungi, which acquire C from their host plants, result in a negative relationship with soil K and MBP81. Soil microbial population, enzymatic activities, and MBP are essential factors in soil fertility status.
Conclusion
The current study results indicated that P and K fertilization increased the nutrient accumulation capabilities of legumes. The microbial biomass phosphorus and soil potassium content increased in the T3 treatment (applied PK fertilizer together). While the K fertilization treatment is more effective for soil nutrient status and increases soil P availability. The application of fertilizer could influence soil properties and enzymatic activity. Current results suggest that P and K fertilization could be applied in different treatments, while combined PK fertilization can increase soil enzymatic activities and increase microorganisms' activities in soil, resulting in improvements to soil fertility and nutrient supply to the plant that could help to improve plant production. Future research needs to explain the role of different species of legumes in the structure of the community of soil microbes, directly measure the soil microorganism’s role which interacted with soil nutrient and the mechanisms underlying their correlations. Additionally, incorporating organic and inorganic fertilizers should be suggested as a superior nutrient management solution for intensive and sustainable green manure crop yields, which subsequently improves main crop yield. Furthermore, research needs to determine the MBP and microbe’s status in adding specific microorganisms (e.g., biological fertilizers).
Data availability
The datasets analyzed during this study are included in this manuscript.
References
Solangi, F. et al. The global dilemma of soil legacy phosphorus and its improvement strategies under recent changes in agro-ecosystem sustainability. ACS Omega https://doi.org/10.1021/acsomega.3c00823 (2023).
Solangi, F. et al. Nutrient uptake potential of nonleguminous species and its interaction with soil characteristics and enzyme activities in the agro-ecosystem. ACS Omega https://doi.org/10.1021/acsomega.3c08794 (2023).
Oosterhuis, D. M., Loka, D. A., Kawakami, E. M. & Pettigrew, W. The physiology of potassium in crop production. Adv. Agron. 126, 203–233 (2014).
Zheng, Y. et al. Effects of water and fertilizer control on soil phosphorus content of Panax notoginseng under micro-sprinkler irrigation. Paiguan Jixie Gongcheng Xuebao/J. Drain Irrig. Mach. Eng. 40, 626–634 (2022).
Bibi, S. et al. Phosphorus extractability in relation to soil properties in different fields of fruit orchards under similar ecological conditions of Pakistan. Front. Ecol. Evol. 10, 1077270 (2023).
Huang, H. et al. The improved winter wheat yield estimation by assimilating GLASS LAI into a crop growth model with the proposed Bayesian posterior-based ensemble kalman filter. IEEE Trans. Geosci. Remote Sens. 61, 1–18 (2023).
Chu, H. et al. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem. 39, 2971–2976 (2007).
Bhuiyan, M., Rahman, M., Afroze, F., Sutradhar, G. & Bhuiyan, M. Effect of phosphorus, molybdenum and Rhizobium inoculation on growth and nodulation of mungbean. J. Soil. Nat. 2, 25–30 (2008).
Amanullah, et al. Compost and nitrogen management influence productivity of spring maize (Zea mays L.) under deep and conventional tillage systems in semi-arid regions. Commun. Soil Sci. Plant Anal. 46, 1566–1578 (2015).
Blanchet, G., Gavazov, K., Bragazza, L. & Sinaj, S. Responses of soil properties and crop yields to different inorganic and organic amendments in a Swiss conventional farming system. Agric. Ecosyst. Environ. 230, 116–126 (2016).
Tang, H., Niu, L., Wei, J., Chen, X. & Chen, Y. Phosphorus limitation improved salt tolerance in maize through tissue mass density increase, osmolytes accumulation, and Na+ uptake inhibition. Front. Plant Sci. 10, 450728 (2019).
Sugito, T., Yoshida, K., Takebe, M., Shinano, T. & Toyota, K. Soil microbial biomass phosphorus as an indicator of phosphorus availability in a Gleyic Andosol. Soil Sci. Plant Nutr. 56, 390–398 (2010).
Yi, J. et al. Assessing soil water balance to optimize irrigation schedules of flood-irrigated maize fields with different cultivation histories in the arid region. Agric. Water Manag. 265, 107543 (2022).
Qiu, S. et al. Carbon storage in an arable soil combining field measurements, aggregate turnover modeling and climate scenarios. Catena 220, 106708 (2023).
Almeida, D. S., Menezes-Blackburn, D., Zhang, H., Haygarth, P. M. & Rosolem, C. A. Phosphorus availability and dynamics in soil affected by long-term ruzigrass cover crop. Geoderma 337, 434–443 (2019).
Zhang, L. et al. Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 210, 1022–1032 (2016).
Dai, H., Wang, X., Deng, Y. & Li, X. Effect of nitrogen supply levels on root morphology and nutrient absorption of rice plants at heading stage. Paiguan Jixie Gongcheng Xuebao/J. Drain Irrig. Mach. Eng. 39, 727–732 (2021).
Soomro, A., Shaikh, A. N., Shaikh, A. N., Ata, A. U. R. & Malik, S. R. Evaluation of different varieties of Chickpea (Cicer Arietinum L.) under agro-ecological conditions of Naudero (District Larkana). Pak-Euro J. Med. Life Sciences 4, 327–336 (2021).
Fageria, N. K., Baligar, V. C. & Bailey, B. A. Role of cover crops in improving soil and row crop productivity. Commun. Soil Sci. Plant Anal. 36, 2733–2757 (2005).
Sulieman, S., Ha, C. V., Schulze, J. & Tran, L. P. Growth and nodulation of symbiotic Medicago truncatula at different levels of phosphorus availability. J. Exp. Bot. 64, 2701–2712 (2013).
Fageria, N. K., Gheyi, H. R., Carvalho, M. C. S. & Moreira, A. Root growth, nutrient uptake and use efficiency by roots of tropical legume cover crops as influenced by phosphorus fertilization. J. Plant Nutr. 39, 781–792 (2016).
Victoria O, Idorenyin U, Asana M, Jia L, Shuoshuo L, Yang S, Okoi IM, Ping A and Egrinya EA. Seed treatment with 24-epibrassinolide improves wheat germination under salinity stress. Asian J. Agric. Biol. 2023(3), https://doi.org/10.35495/ajab.2022.076(2023).
Zhou, Y., Berruti, F., Greenhalf, C. & Henry, H. A. L. Combined effects of biochar amendment, leguminous cover crop addition and snow removal on nitrogen leaching losses and nitrogen retention over winter and subsequent yield of a test crop (Eruca sativa L.). Soil Biol. Biochem. 114, 220–228 (2017).
Meena, A. & Rao, K. S. Assessment of soil microbial and enzyme activity in the rhizosphere zone under different land use/cover of a semiarid region. India. Ecol. Process. 10, 1–12 (2021).
Li, H. et al. Microbial inoculants and struvite improved organic matter humification and stabilized phosphorus during swine manure composting: Multivariate and multiscale investigations. Bioresour. Technol. 351, 126976 (2022).
Zhang, T. et al. Organic matter stabilization and phosphorus activation during vegetable waste composting: Multivariate and multiscale investigation. Sci. Total Environ. 891, 164608 (2023).
Trasar-Cepeda, C., Leirós, M. C. & Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 40, 2146–2155 (2008).
Idris I and Yuliar Y. Potential application of Bacillus amyloliquefaciens EB13 inoculant for improving soil fertility and Citrus sinensis growth. Asian J. Agric. Biol. 2022(1), 202102069. https://doi.org/10.35495/ajab.2021.02.069 (2022)
Zhang, T., Li, P., Fang, C. & Jiang, R. Phosphate recovery from animal manure wastewater by struvite crystallization and CO2 degasification reactor. Ecol. Chem. Eng. S 21, 89–99 (2014).
Adetunji, A. T., Lewu, F. B., Mulidzi, R. & Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 17, 794–807 (2017).
Burns, R. G. et al. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 58, 216–234 (2013).
Jian, S. et al. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 101, 32–43 (2016).
Smith, S. E., Anderson, I. C. & Smith, F. Mycorrhizal associations and phosphorus acquisition: from cells to ecosystems. Ann. Plant Rev. Vol. 48: Phosphorus Metab. Plants 48, 409–440 (2015).
Solangi, F. et al. Improved accumulation capabilities of phosphorus and potassium in green manures and its relationship with soil properties and enzymatic activities. Agronomy 9, 708 (2019).
Thornburg, T. E. et al. Potassium deficiency significantly affected plant growth and development as well as microRNA-mediated mechanism in wheat (Triticum aestivum L.). Front. Plant Sci. 11, 1219 (2020).
Congreves, K. A. et al. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 12, 637108 (2021).
Rawal, N., Pande, K. R., Shrestha, R. & Vista, S. P. Nutrient use efficiency (NUE) of wheat (Triticum aestivum L.) as affected by NPK fertilization. PLoS ONE 17, e0262771 (2022).
Jodaugiene, D. et al. The influence of organic mulches on soil biological properties. Zemdirbyste 97, 33–40 (2010).
Yang, Y. J. et al. Effect of organic mulches on soil bacterial communities one year after application. Biol. Fertil. Soils 38, 273–281 (2003).
Wang, C. et al. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 121, 103–112 (2018).
Soon, Y. K. & Kalra, Y. P. Short communication: A comparison of plant tissue digestion methods for nitrogen and phosphorus analyses. Can. J. Soil Sci. 75, 243–245 (1995).
Walker, J. M. & Barber, S. A. Absorption of potassium and rubidium from the soil by corn roots. Plant Soil 17, 243–259 (1962).
Antweiler, R. C., Patton, C. J. & Taylor, H. E. Automated, Colorimetric methods for determination of nitrate plus nitrite, nitrite, ammonium and orthophosphate ions in natural water samples. U.S.G.S. Rep. 93, 1–28 (1996).
Nelson, D. & Sommers, L. Chemical methods Soil Sci. Soc. Am. 961–1010 (Madison, 1996).
Brookes, P. C., Powlson, D. S. & Jenkinson, D. S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 14, 319–329 (1982).
Chafai W, EL Gabardi S, Douira A and Khalid A. Diversity and mycorrhizal potential of arbuscular mycorrhizal fungi in two natural soils in the eastern region of Morocco. Asian J. Agric. Biol. 2022(2), 202102101 https://doi.org/10.35495/ajab.2021.02.101 (2022)
Murphy, J. & Riley, J. P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 27, 31–36 (1962).
Zelazny, L. W., He, L. & Vanwormhoudt, A. M. In Methods of Soil Analysis, Part 3: Chemical Methods 1231–1253 (2018). https://doi.org/10.2136/sssabookser5.3.c41
Kandeler, E. & Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 6, 68–72 (1988).
Eivazi, F. & Tabatabai, M. A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 20, 601–606 (1988).
Schneider, K., Turrion, M. B. & Gallardo, J. F. Modified method for measuring acid phosphatase activities in forest soils with high organic matter content. Commun. Soil Sci. Plant Anal. 31, 3077–3088 (2000).
Weaver, R. W., Angle, J. S. & Bottomley, P. J. Methods of Soil Analysis, Part 2: Microbiological and Biochemical Properties. Methods of Soil Analysis, Part 2: Microbiological and Biochemical Properties (2018). https://doi.org/10.2136/sssabookser5.2
Melander, B., Rasmussen, I. A. & Olesen, J. E. Legacy effects of leguminous green manure crops on the weed seed bank in organic crop rotations. Agric. Ecosyst. Environ. 302, 107078 (2020).
Youming, M., Fangfang, M. & Pengnian, W. U. Effects of nitrogen application rates on soil water, temperature, carbon nitrogen, and maize yield in Yanghuang irrigation area. J. Drain. Irrig. Mach. Eng. 39, 950–958 (2021).
Ros, M. B. H. et al. Towards optimal use of phosphorus fertiliser. Sci. Rep. 10, 17804 (2020).
Bindraban, P. S., Dimkpa, C. O. & Pandey, R. Exploring phosphorus fertilizers and fertilization strategies for improved human and environmental health. Biol. Fertil. Soils 56, 299–317 (2020).
Belay, A., Claassens, A. S. & Wehner, F. C. Effect of direct nitrogen and potassium and residual phosphorus fertilizers on soil chemical properties, microbial components and maize yield under long-term crop rotation. Biol. Fertil. Soils 35, 420–427 (2002).
Irfan, M., Shah, J. A. & Abbas, M. Evaluating the performance of mungbean genotypes for grain yield, phosphorus accumulation and utilization efficiency. J. Plant Nutr. 40, 2709–2720 (2017).
Bindraban, P. S., Dimkpa, C., Nagarajan, L., Roy, A. & Rabbinge, R. Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 51, 897–911 (2015).
Simpson, R. J. et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 349, 89–120 (2011).
Barłóg, P., Grzebisz, W. & Łukowiak, R. Fertilizers and fertilization strategies mitigating soil factors constraining efficiency of nitrogen in plant production. Plants 11, 1855 (2022).
Wu, J. et al. Dynamics in microbial immobilization and transformations of phosphorus in highly weathered subtropical soil following organic amendments. Plant Soil 290, 333–342 (2007).
Heidari, G., Mohammadi, K. & Sohrabi, Y. Responses of soil microbial biomass and enzyme activities to tillage and fertilization systems in soybean (Glycine max L.) Production. Front. Plant Sci. 7, 207995 (2016).
Jindo, K. et al. Biotic and abiotic effects of soil organic matter on the phytoavailable phosphorus in soils: A review. Chem. Biol. Technol. Agricult. 10, 29 (2023).
Xu, X. et al. Effects of potassium levels on plant growth, accumulation and distribution of carbon, and nitrate metabolism in apple dwarf rootstock seedlings. Front. Plant Sci. 11, 534048 (2020).
Bowles, T. M., Acosta-Martínez, V., Calderón, F. & Jackson, L. E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 68, 252–262 (2014).
Olander, L. P. & Vitousek, P. M. Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 49, 175–190 (2000).
Ning, C. C. et al. Impacts of chemical fertilizer reduction and organic amendments supplementation on soil nutrient, enzyme activity and heavy metal content. J. Integr. Agric. 16, 1819–1831 (2017).
Piotrowska, A. & Koper, J. Soil β-glucosidase activity under winter wheat cultivated in crop rotation systems depleting and enriching the soil in organic matter. J. Elem. 15, 593–600 (2010).
Gu, Y., Zhang, X., Tu, S. & Lindström, K. Soil microbial biomass, crop yields, and bacterial community structure as affected by long-term fertilizer treatments under wheat-rice cropping. Eur. J. Soil Biol. 45, 239–246 (2009).
Tian, W. et al. Responses of microbial activity, abundance, and community in wheat soil after three years of heavy fertilization with manure-based compost and inorganic nitrogen. Agric. Ecosyst. Environ. 213, 219–227 (2015).
Moghimian, N., Hosseini, S. M., Kooch, Y. & Darki, B. Z. Impacts of changes in land use/cover on soil microbial and enzyme activities. Catena 157, 407–414 (2017).
Kooch, Y., Ehsani, S. & Akbarinia, M. Stoichiometry of microbial indicators shows clearly more soil responses to land cover changes than absolute microbial activities. Ecol. Eng. 131, 99–106 (2019).
Malik, M. A., Khan, K. S. & Marschner, P. Microbial biomass, nutrient availability and nutrient uptake by wheat in two soils with organic amendments. J. Soil Sci. Plant Nutr. 13, 955–966 (2013).
Mahmud, K., Makaju, S., Ibrahim, R. & Missaoui, A. Current progress in nitrogen fixing plants and microbiome research. Plants 9, 97 (2020).
Wardah, L. Y., Fuakubun, F. & Sopandi, T. Valorization of chicken feather into organic liquid fertilizer through two species of Bacillus bacteria fermentation. Asian J. Agric. Biol. 2023(4), 2022148. https://doi.org/10.35495/ajab.2022.148 (2023).
Schneider, K. D. et al. Microbially-mediated P fluxes in calcareous soils as a function of water-extractable phosphate. Soil Biol. Biochem. 106, 51–60 (2017).
Feng, X. et al. Soil ionomic and enzymatic responses and correlations to fertilizations amended with and without organic fertilizer in long-term experiments. Sci. Rep. 6, 24559 (2016).
Balota, E. L. & Chaves, J. C. D. Enzymatic activity and mineralization of carbon and nitrogen in soil cultivated with coffee and green manures. Rev. Bras. Cienc. do Solo 34, 1573–1583 (2010).
Abd El-Fattah, D. A., Hashem, F. A. & Abd-Elrahman, S. H. Impact of applying organic fertilizers on nutrient content of soil and lettuce plants, yield quality and benefit-cost ratio under water stress conditions. Asian J. Agric. Biol. 2, 202102086. https://doi.org/10.35495/ajab.2021.02.086 (2022).
Liu, L. et al. Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest. PLoS ONE 8, e61188 (2013).
Acknowledgements
The authors extend their appreciation to the Researchers Supporting Project number (RSP2024R185), King Saud University, Riyadh, Saudi Arabia.
Statement on guidelines
All experimental studies and experimental materials involved in this research are in full compliance with relevant institutional, national and international guidelines and legislation.
Funding
Open Access funding enabled and organized by Projekt DEAL. The authors are greatly indebted to the Key R&D Project of Jiangsu Province (Modern Agriculture) (No. BE2022351), and Project of Faculty of Agricultural Equipment of Jiangsu University (No. NZXB20210101). The authors also extend their appreciation to the Researchers Supporting Project number (RSP2024R185), King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
Conceptualization, investigation, data collection, and writing—original draft, F.S., X.Z., K.A.S and R.I. Software, script evaluation, and editing, M.S.E., K.M.A, and H.H.E. Writing—review and editing, F.S., X.Z., K.A.S and R.I. Data curation and formal analysis, M.S.E., K.M.A, and H.H.E. Visualization and validation, M.S.E., R.I. Methodology and references, F.S., X.Z., K.A.S and R.I. Supervision, F.S., X.Z., K.A.S. Funding Acquisition, H.H.E., and R.I. All authors reviewed the manuscript and agreed for final publication.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Solangi, F., Zhu, X., Solangi, K.A. et al. Responses of soil enzymatic activities and microbial biomass phosphorus to improve nutrient accumulation abilities in leguminous species. Sci Rep 14, 11139 (2024). https://doi.org/10.1038/s41598-024-61446-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-61446-z
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
