


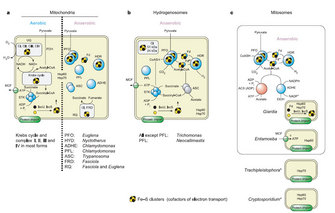
 Figure 1: Enzymes and pathways found in various manifestations of mitochondria
Figure 1: Enzymes and pathways found in various manifestations of mitochondria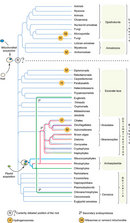
















« Prev Next »
What variety is there in mitochondria? Mitochondria occur in various forms across various eukaryotic groups, yet considerations on the origin of mitochondria sometimes neglect this understanding. Four main mitochondrial types can be distinguished on the basis of functional criteria concerning how or whether ATP is produced. These functional types do not correspond to natural groups, because they occur in an interleaved manner across the tree of eukaryotic life. Instead they correspond to ecological specializations.
Mitochondria: A Ubiquitous and Diverse Family of Organelles
© 2006 Nature Publishing Group Embley, T. M. & Martin. W. Eukaryotic evolution, changes and challenges. Nature 440, 623–630 (2006). All rights reserved. 
The mitochondria typical of mammalian cells respire O2 during the process of pyruvate breakdown and ATP synthesis, generating water and carbon dioxide as end products. The Krebs cycle and the electron transport chain in the inner mitochondrial membrane enable the cell to generate about 36 moles (mol) of ATP per mole of glucose, with the help of O2–respiring mitochondria. Such typical mitochondria also occur in plants and various groups of unicellular eukaryotes (protists) that, like mammals, are dependent on oxygen and specialized to life in oxic environments.
In contrast, the mitochondria of many invertebrates (worms like Fasciola hepatica and mollusks like Mytilus edulis being well–studied cases) do not use O2 as the terminal acceptor during prolonged phases of the life cycle. These mitochondria allow the anaerobically growing cell to glean about 5 mol of ATP per mole of glucose, as opposed to about 36 with O2. The typical excreted end products are carbon dioxide, acetate, propionate, and succinate, which are generated mostly through the rearrangement of Krebs cycle reactions and the help of the mitochondrial electron transport chain. These organelles are commonly called anaerobic mitochondria.
Mitochondria of yet another kind yield even less ATP per molecule of glucose. These are mitochondria of several distantly related unicellular eukaryotes (protists) that lack an electron transport chain altogether. They synthesize ATP from pyruvate breakdown via simple fermentations that typically involve the production of molecular hydrogen as a major metabolic end product. These mitochondria are called hydrogenosomes and allow the cell to gain about 4 mol of ATP per mole of glucose. Hydrogenosomes were discovered in 1973 in trichomonads, a group of unicellular eukaryotes. They were later found in chytridiomycete fungi that inhabit the rumen of cattle, as well as some ciliates, and they continue to be found in other groups. The enzymes of hydrogenosomes are not unique to these anaerobes. They are found also in the mitochondria, the cytosol, or even the plastids of other eukaryotes (Figure 1).
A fourth category of eukaryotes possesses small, inconspicuous mitochondria that are not involved in ATP synthesis at all. These eukaryotes synthesize their ATP in the cytosol with the help of enzymes that are otherwise typically found in hydrogenosomes. They obtain 2-4 mol of ATP per mole of glucose. Their typical end products are carbon dioxide, acetate, and ethanol, and their mitochondria are called mitosomes. Mitosomes were discovered in the human intestinal parasite Entamoeba histolytica in 1999, and were subsequently found in many additional eukaryotes, including Giardia lamblia in 2003.
Knowledge about these different forms of mitochondria comes from decades of biochemical and physiological investigations of eukaryotic anaerobes, many of which are important pathogens or parasites of humans and livestock. Well into the 1990s it was widely thought that several anaerobic eukaryotes, such as Giardia lamblia, lack mitochondria altogether and had never possessed them in the evolutionary past. Newer work, however, has shown that mitochondria are just as defining and ubiquitous among eukaryotes as is the nucleus itself. That realization has had considerable impact on current views about the origin of mitochondria.
The Endosymbiotic Origin of Mitochondria
There are currently two main, competing theories about the origin of mitochondria. They differ with regard to their assumptions concerning the nature of the host, the physiological capabilities of the mitochondrial endosymbiont, and the kinds of ecological interactions that led to physical association of the two partners at the onset of symbiosis.

© 2006 Nature Publishing Group Embley, T. M. & Martin. W. Eukaryotic evolution, changes and challenges. Nature 440, 623–630 (2006). All rights reserved.
An alternative theory posits that the host that acquired the mitochondrion was a prokaryote, an archaebacterium outright. This view is linked to the idea that the ancestral mitochondrion was a metabolically versatile, facultative anaerobe (able to live with or without oxygen), perhaps similar in physiology and lifestyle to modern Rhodobacteriales. The initial benefit of the symbiosis could have been the production of H2 by the endosymbiont as a source of energy and electrons for the archaebacterial host, which is posited to have been H2 dependent. This kind of physiological interaction (H2 transfer or anaerobic syntrophy) is commonly observed in modern microbial communities. The mechanism by which the endosymbiont came to reside within the host is unspecified in this view, but in some known examples in nature prokaryotes live as endosymbionts within other prokaryotes. In this view, various aerobic and anaerobic forms of mitochondria are seen as independent, lineage-specific ecological specializations, all stemming from a facultatively anaerobic ancestral state. Because it posits that eukaryotes evolved from the mitochondrial endosymbiosis in a prokaryotic host, this theory directly accounts for the ubiquity of mitochondria among all eukaryotic lineages.
Eukaryotes are genetic chimeras. They possess genes that they inherited vertically from their archaebacterially related host. Genes for cytosolic ribosomes in eukaryotes, for example, reflect that origin. But eukaryotes also possess genes that they inherited vertically from the endosymbiont - for example, mitochondrially encoded genes for mitochondrial ribosomes. But even the largest mitochondrial genomes possess only about sixty protein-coding genes, while typical mitochondria harbor up to a thousand proteins or more that are encoded in the nucleus. During the course of mitochondrial genesis, many genes were transferred from the genome of the mitochondrial endosymbiont to the genome of the host. This kind of endosymbiotic gene transfer is nothing unusual; endosymbiosis very often entails gene transfers from the endosymbiont to the host. It happened during the origin of plastids too, and it is still ongoing in our own genome: Mitochondrial DNA constantly escapes from the organelle and becomes integrated as copies into nuclear DNA. The vast majority of mitochondrial proteins are encoded by nuclear genes, and many of these are endosymbiotic acquisitions from the mitochondrial ancestor.
When and How Often Did Mitochondria Arise?
The oldest undisputedly eukaryotic microfossils go back
1.45 billion years in the fossil record. Given the coincidence of mitochondria
with the eukaryotic state, this can also be seen as a minimum age for
mitochondria and a rough best-guess starting date for eukaryotic evolution.
According to newer geochemical views, this date of origin corresponds to a
protracted phase in Earth history when the oceans were mostly anoxic — from 1.8
billion years ago until about 580 million years ago — because of the workings
of marine, H2S-producing bacteria. Eukaryotes thus arose and
diversified in an environment where anoxia was commonplace. Accordingly it is
hardly surprising that many independent eukaryotic lineages have preserved
anaerobic energy-producing pathways in their mitochondria (Figure 3).
Like eukaryotes themselves, mitochondria appear to have arisen only once in all of evolution. The best evidence for the single origin of mitochondria comes from a conserved set of clearly homologous and commonly inherited genes preserved in the mitochondrial DNA across all known eukaryotic groups. In the case of hydrogenosomes (which usually lack DNA) and mitosomes (which so far always lack DNA), the strongest evidence for their common ancestry with mitochondria is twofold. First, aspects and components of the mitochondrial protein import process are conserved in hydrogenosomes and mitosomes, arguing strongly for common ancestry with mitochondria. Second, all known lineages of eukaryotes that possess hydrogenosomes or mitosomes branch as sisters to mitochondrion-bearing lineages.
Summary
Mitochondria arose once in evolution, and their origin entailed an endosymbiosis accompanied by gene transfers from the endosymbiont to the host. Anaerobic mitochondria pose a puzzle for traditional views on mitochondrial origins but fit nicely in newer theories on mitochondrial evolution that were formulated specifically to take the common ancestry of mitochondria and hydrogenosomes into account. The presence of mitochondria in the eukaryote common ancestor continues to change the way we look at eukaryote origins, with endosymbiosis playing a more central role in considerations on the matter now than it did twenty years ago. The integral part that mitochondria play in many aspects of eukaryote biology might well reflect their role in the origin of eukaryotes themselves.
References and Recommended Reading
Boxma, B., et al. An anaerobic mitochondrion that produces hydrogen. Nature 434, 74–79 (2005) doi:10.1038/nature03343.
Cox, C. J., et al. The archaebacterial origin of eukaryotes. PNAS 105, 20356–20361 (2008) doi:10.1073/pnas.0810647105.
Dolezal, P., et al. Evolution of the molecular machines for protein import into mitochondria. Science 313, 314–318 (2006) doi:10.1126/science.1127895.
Dyall, S. D., et al. Ancient invasions: From endosymbionts to organelles. Science 304, 253–257 (2004) doi:10.1126/science.1094884.
Embley, T. M., & Martin, W. Eukaryotic evolution, changes and challenges. Nature 440, 623–630 (2006) doi:10.1038/nature04546.
Hazkani-Covo, E., et al. Molecular poltergeists: Mitochondrial DNA copies (numts) in sequenced nuclear genomes. PLoS Genetics 6, e1000834 (2010) doi:10.1371/journal.pgen.1000834.
John, P., & Whatley, F. R. Paracoccus denitrificans and the evolutionary origin of the mitochondrion. Nature 254, 495–498 (1975) doi:10.1038/254495a0.
Lane, N. Life Ascending: The Ten Greatest Inventions of Evolution. London: Profile Books, 2009.
Lane, N. Power, Sex, Suicide: Mitochondria and the Meaning of Life. Oxford: Oxford University Press, 2006.
Martin, W., & Müller, M. The hydrogen hypothesis for the first eukaryote. Nature 392, 37–41 (1998) doi:10.1038/32096.
Mentel, M., & Martin, W. Energy metabolism among eukaryotic anaerobes in light of Proterozoic ocean chemistry. Philosophical Transactions of the Royal Society of London B: Biological Sciences 363, 2717–2729 (2008) doi:10.1098/rstb.2008.0031.
Müller, M. The hydrogenosome. Journal of General Microbiology 139, 2879-2889 (1993).
Tielens A. G. M., et al. Mitochondria as we don't know them. Trends in Biochemical Sciences 27, 564–572 (2002) doi:10.1016/S0968-0004(02)02193–X.
Timmis, J. N., et al. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nature Reviews Genetics 5, 123–135 (2004) doi:10.1038/nrg1271.
Van Valen, L. M., & Maiorana V. C. The Archaebacteria and eukaryotic origins. Nature 287, 248–250 (1980) doi:10.1038/287248a0.










