


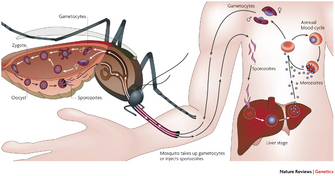
 Figure 1: Plasmodium falciparum life cycle
Figure 1: Plasmodium falciparum life cycle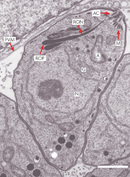
















« Prev Next »
In the 1970s, scientists discovered a novel organelle in apicomplexan parasites, which was named the apicoplast. Like mitochondria and chloroplasts, apicoplasts contain their own DNA. When scientists analyzed apicoplast DNA, they were surprised to learn that apicoplasts shared sequence similarities with plastids (organelles found in the cells of photosynthetic organisms like algae and plants). Could the apicoplast be a vestigial chloroplast? How could apicomplexans and plants be related? What follows is the amazing story of how apicoplasts were discovered and how they have become a promising target for drug development.
Apicomplexans
© 2007 Nature Publishing Group Su, X., Hayton, K., & Wellems, T. E. Genetic linkage and association analyses for trait mapping in Plasmodium falciparum. Nature Reviews Genetics 8, 497–506 (2007). All rights reserved. 
In contrast, human toxoplasmosis produces a mild disease, often without symptoms. It is acquired by ingesting infective Toxoplasma parasites, which can grow in all nucleated cells, including intestinal, muscle and brain cells. The parasite can also be transmitted from mother to fetus. The definitive hosts (in which sexual reproduction occurs) are cats and other felines, and humans are the intermediate hosts simply by accident (Figure 2). The disease is only serious (even fatal) in individuals with weakened immune systems (e.g., AIDS patients), and during pregnancy because it can cause severe developmental malformations. An estimated one third of the world's human population has been infected with Toxoplasma.
Eimeriosis is a disease caused by Eimeria parasites. Unlike Plasmodium and Toxoplasma, Eimeria requires a single host to complete its life cycle. Eimeria infections are a significant cause of disease in poultry, cattle and sheep.
At first glance, these three parasites seem to be very different in terms of their life cycles, hosts, and disease severity. However, despite their differences, the parasites all share a common structure known as the apical complex. This unique structure contains secretory organelles (rhoptries, micronemes, and dense granules), which sequentially secrete enzymes that allow the parasite to invade other cells. Because Plasmodium, Toxoplasma, and Eimeria all have apical complexes, they are members of the same club-the phylum Apicomplexa. All apicomplexans are obligate, intracellular, protozoan parasites, that is, they cannot live or reproduce without a host.
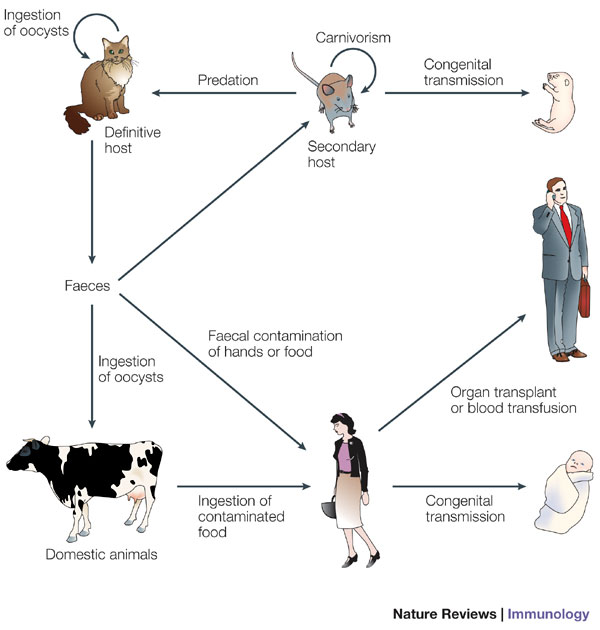
Figure 2: Toxoplasma gondii life cycle
© 2005 Nature Publishing Group Aliberti, J. Host persistence: exploitation of anti-inflammatory pathways by Toxoplasma GONDII. Nature Reviews Immunology 5, 162–170 (2005). All rights reserved.
Discovery of the Apicoplast

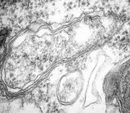
Figure 3: Separation of DNA in CsCl gradients
This photograph is an example of DNA separated in a cesium chloride density gradient. Two tubes containing cesium chloride gradients are arranged vertically next to each other. The gradients have higher densities toward the bottom of the tube and lower densities toward the top of the tube. The solutions contain ethidium bromide, which causes the DNA to appear as fluorescent bands. The tube on the left contains a single horizontal band of DNA, indicating that it has DNA of one density. The tube on the right has two horizontal bands: one band is at the same position in this tube as the band in the tube on the left, and the other band, which has a white arrow pointing to it, is lower in the tube. The lower band contains DNA of a higher density.
© 2000 Nature Publishing Group Radajewski, S., Ineson, P., Parekh, N. R., & Murrell, J. C. Stable-isotope probing as a tool in microbial ecology. Nature 403, 646–649 (2000). All rights reserved.
In the1970s, another clue was added to the puzzle by scientists who were characterizing the extrachromosomal DNA molecules in apicomplexans. According to the theory of endosymbiosis, they knew that some cellular organelles, like mitochondria and chloroplasts, contain their own independent genomes, which are separate from the nuclear genome and very similar to bacterial genomes. Using cesium chloride (CsCl) density gradients to separate DNA molecules by centrifugation (Figure 3), the scientists isolated what they believed to be mitochondrial DNAs from P. knowlesi (primate malaria parasites) and P. berghei (rodent malaria parasites). Araxie Kilejian, a post-doctoral scientist at Rockefeller University was intrigued by the results. She wondered why mitochondrial DNAs had not been detected in P. lophurae and P. gallinaceum, since both avian malaria parasites contained visible mitochondria. Kilejian also wanted to learn more about the nature of these extrachromosomal molecules. She soon realized that P. lophurae's extrachromosomal DNAs had the same density as the nuclear DNA, making it difficult to distinguish between them using regular CsCl gradients. She decided to perform careful sub-cellular fractionations and electron micrograph examinations. Using these methods, she succeeded in isolating a population of circular molecules that were estimated to be 10.3 µm (micrometers) in length and 27–kilobases in size (Kilejian 1975). A kilobase, or kb, is the unit of measure used by molecular biologists to measure the size of DNA or RNA fragments. One kb contains 1000 nucleotide base pairs.
At the time, the prevailing "rule of the ring" dogma was that most, if not all, mitochondrial DNAs were circular, so Kilejian mistook these circular structures for mitochondrial DNAs. Later, several groups confirmed her discovery by isolating equivalent extrachromosomal, circular DNA molecules from other apicomplexan species, including P. berghei (11.3 µm), T. gondii (13 µm), P. knowlesi (11.6 µm), and the human malarial parasite, P. falciparum (11.1 µm). Since the experimental data available did not suggest any alternative interpretation, scientists continued to believe that the circular, extrachromosomal DNAs were mitochondrial DNA until late in the 1980s.
In 1987, the acceptance of the "rule of the ring" dogma was called into question when a second extrachromosomal genome — a tandemly arranged, 6-kb, linear DNA molecule — was identified in the rodent malarial parasite P. yoelli (Vaidya & Arasu 1987, Vaidya et al. 1989). Analysis of its DNA and protein sequences showed that it had similarities to mitochondrion-encoded proteins. Two years later, Iain Wilson and his colleagues identified a similar 6-kb element in P. falciparum, and to everyone's surprise, they confirmed that the 6-kb element was the real mitochondrial DNA.
How did they prove that the 6-kb element was the mitochondrial genome? Wilson and his colleagues sequenced the element and detected typical mitochondrial genes in the genome, such as the cytochrome and cytochrome oxidase genes (Feagin et al. 1991). Later, they demonstrated that when purifying subcellular fractions enriched in mitochondria, only the linear genome co-purified with the mitochondria, not the circular DNA. This confirmed that the mitochondria and circular DNA were located in different cellular compartments (Wilson et al. 1992). The scientists could not have been more intrigued. Since the 6-kb element represented the mitochondrial genome, what was the nature of the circular extrachromosomal DNA element? The scientists began exploring alternative explanations for the origin and function of the circular genome.
An Organelle with a Green Past
Iain Wilson teamed with an expert mitochondriologist, Donald Williamson, and researchers Malcolm Gardner and Jean Feagin to study malarial extrachromosomal DNAs. In 1991, the group published a paper in Parasitology Today (now Trends in Parasitology) entitled "Have malaria parasites three genomes?" (Wilson et al. 1991). This title was intentionally provocative. At the time, biologists agreed - and this was the second dogma - that the only eukaryotic cells with three different genomes were the plants and algae, which had nuclear, mitochondrial and plastid genomes. In the Parasitology Today article, Wilson and his group analyzed the circular extrachromosomal DNA. They determined that the size of the molecule was 35 kb using restriction mapping and sequencing (Gardner et al. 1991a). The scientists learned that the circular element contained large inverted repeats with duplicated sets of ribosomal RNAs (rRNAs), which was characteristic of chloroplast genomes. Moreover, when they performed a phylogenetic analysis of the conserved regions of the small subunit rRNA, they noticed similarities between the extrachromosomal genome and several prokaryotic and chloroplast counterparts (Gardner et al. 1991b). Wilson and his colleagues' results were leading them to the conclusion that malaria contained a plastid. Where was this DNA molecule located?
Many scientists participated in the race to locate the 35-kb DNA molecule within the parasite's cell. Araxie Kilejian suggested the possibility that the 35-kb element resided in the "spherical bodies," but there was no experimental evidence to support her hypothesis (Kilejian 1991). It would take another six years before the 35-kb DNA was conclusively shown to associate with the "spherical bodies," following cross-field collaborations between plant scientists and parasitologists. How did these collaborations come about?
A Definition of a New Organelle
Since Geoffrey McFadden and Ross Waller were plant scientists, they did not typically read journals on parasitology. Quite serendipitously, they came across Wilson's paper in Parasitology Today. As plant scientists, they were well aware of the dogma that only algae and plants have three genomes. So, when they read the paper's title about malaria parasites having three genomes, this was equivalent to saying that malarial parasites were plants! McFadden and Waller were intrigued - they wanted to "see" the putative plastid first hand and understand the meaning of this amazing discovery, but where could they start? As their knowledge of parasites was minimal, they sought help from Alan Cowman, a malariologist, and Naomi Lang-Unnasch, who researched Toxoplasma. After several unsuccessful attempts to locate the 35-kb DNA within the Plasmodium parasites, they decided to try experimenting with Toxoplasma. They were aware of research by Lang-Unnasch, who had isolated a plastid-like ribosomal RNA gene from Toxoplasma parasites. Since ribosomal RNAs are very abundant transcripts, they thought this would be a sensitive probe for in situ hybridization (Egea & Lang 1995). They labeled the probe and used it to perform high-resolution, in situ hybridizations of ultrathin sections of Toxoplasma parasites, and then viewed the sections under an electron microscope. The electron micrographs showed that the probe labeled a single, small, oviform organelle located in the mid to anterior region of the cell, which was anterior to the nucleus and near the Golgi apparatus (Figure 4). In contrast, the mitochondria were not labeled. These experiments clearly showed that the 35-kb genome was located in the ovoid organelle (McFadden et al. 1996). Moreover, when observing the micrographs, McFadden and his group noticed that the organelle was surrounded by at least two membranes. Like plants, and green and red algae (more recently, four membranes have been identified in the Plasmodium apicoplast; Ralph et al. 2004) (Figure 5). As in most algae, they noticed that only one plastid and one mitochondrion was present per cell. 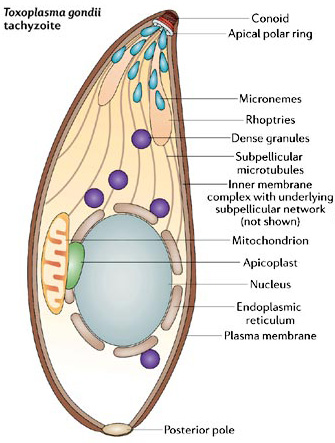
One year later, in 1997, the McFadden group's results were confirmed by Sabine Köhler and her colleagues (one of whom was Iain Wilson). The general conclusion was that the apicoplast was indeed a vestigial plastid — a finding that broke another dogma. In their article, Köhler and colleagues proposed naming the recently identified organelle the "apicoplast," which was a combination of the words "apicomplexan" and "plastid" (Köhler et al. 1997). Finally, the new organelle had a name. Since then, apicoplasts have been detected in all apicomplexan parasites studied, with the exception of Cryptosporidium (responsible for human cryptosporidiosis) (Figure 6).
The analysis of both the genomic sequence and the organization of the 35-kb apicoplast genome led to two main conclusions. The first was that the apicoplast genome encodes enough transfer RNAs (tRNAs) and ribosomal genes for a minimal, but sufficient, translation of the protein-encoding genes present in the element. The second conclusion was that the genomic organization showed a close relationship to algae (Wilson et al. 1996). In 2008, in an exciting twist in the story, scientists studied a minute marine alga that provided evidence that apicomplexan parasites evolved from photosynthetic algal ancestors (Keeling 2008).
Figure 6: The apicomplexan's plastid
© 2006 Nature Publishing Group Baum, J. et al. Regulation of apicomplexan actin-based motility. Nature Reviews Microbiology 4, 621–628 (2006). All rights reserved.
The Apicoplast as a Drug Target
© 2002 Nature Publishing Group Gardner, M. J. et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 419, 498–511 (2002). All rights reserved.
Two different lines of evidence showed that apicoplasts are essential in the apicomplexan parasites. First, chemicals affecting apicoplast metabolism resulted in parasite death. Second, parasites that are unable to replicate the apicoplast also die. Amazingly, in both cases, the parasites only die in the next generation. This means that the parasites can survive with no apicoplast (or with a chemically damaged apicoplast) while remaining in the infected host cell. However, the parasite is unable to establish a successful new infection. This phenomenon is called "delayed death". How is this possible? One hypothesis is that the apicoplast synthesizes a molecule that is needed for the infection process (Ralph et al. 2004).
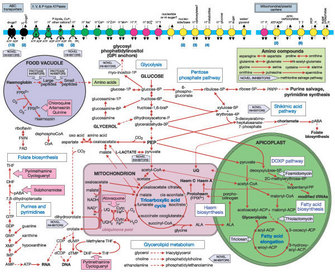
Since apicoplasts share evolutionary similarities with chloroplasts and prokaryotes (cyanobacteria), they stand out as an attractive target for known antibiotics and herbicides. Today, researchers have learned that apicoplasts are sensitive to several antibacterials and herbicides. Plant scientists are already exploring non-toxic herbicides that may act upon the apicoplast, and together with parasitologists, they are using bioinformatics and experimental approaches to identify key proteins and metabolic pathways in order to develop new drugs (Ralph et al. 2004) (Figure 7).
Summary
Apicoplasts, with their green past, present a unique difference between the host and the parasite, offering an excellent target for the development of new chemotherapeutic alternatives. Could apicoplasts be the long searched for "Achilles' heel" to prevent the spread of malaria? Only time will tell. What is certain is that it will no longer be uncommon to see plant scientists reading journals on parasitology, or parasitologists making their way through plant journals.
References and Recommended Reading
Egea, N. & Lang-Unnasch, N. Phylogeny of the large extrachromosomal DNA of organisms in the phylum Apicomplexa. Journal of Eukaryotic Microbiology 42, 679–684 (1995). doi: 10.1111/j.1550-7408.1995.tb01615.x
Feagin, J. E., Gardner, M. J., et al. The putative mitochondrial genome of Plasmodium falciparum. Journal of Eukaryotic Microbiology38, 243–245 (1991). doi: 10.1111/j.1550-7408.1991.tb04436.x
Gardner, M. J., Feagin, J. E., et al. Organization and expression of small subunit ribosomal RNA genes encoded by a 35–kilobase circular DNA in Plasmodium falciparum. Molecular and Biochemical Parasitology 48, 77–88 (1991b). doi:10.1016/0166-6851(91)90166–4
Gardner, M. J., Williamson, D. H., et al. A circular DNA in malaria parasites encodes an RNA polymerase like that of prokaryotes and chloroplasts. Molecular and Biochemical Parasitology 44, 115–23 (1991a). doi:10.1016/0166-6851(91)90227–W
Keeling, P. J. Bridge over troublesome plastids. Nature 451, 896–897 (2008). doi:10.1038/451896a
Kilejan, A. Circular mitochondrial DNA from the avian malarial parasite Plasmodium lophurae. Biochimica et Biophysica Acta 390, 276–284 (1975). doi:10.1016/0005-2787(75)90348–2
Kilejian, A. Spherical bodies. Parasitology Today 7, 309; author reply 309 (1991).
Köhler, S., Delwiche, C. F., et al. A plastid of probable green algal origin in apicomplexan parasites. Science 275, 1485–1488 (1997). doi: 10.1126/science.275.5305.1485
McFadden, G. I., Reith, M. E., et al. Plastid in human parasites. Nature 381, 482 (1996). doi:10.1038/381482a0
Ralph, S. A., van Dooren, G. G., et al. Tropical infectious diseases: metabolic maps and functions of the Plasmodium falciparum apicoplast. Nature Reviews Microbiology 2, 203–16 (2004). doi:10.1038/nrmicro843
Scholtyseck, E. & Piekarski, G. Electron microscopic studies on merozoites of Eimeria (Eimeria perforans and E. stidae) and Toxoplasma gondii. On the systematic position of T. gondii. Zeitschrift für Parasitenkunde 26, 91–115 (1965).
Vaidya, A. B. & Arasu, P. Tandemly arranged gene clusters of malaria parasites that are highly conserved and transcribed. Molecular and Biochemical Parasitology 22, 249–257 (1987). doi:10.1016/0166-6851(87)90056–9
Vaidya, A. B., Akella, R., et al. Sequences similar to genes for two mitochondrial proteins and portions of ribosomal RNA in tandemly arrayed 6-kilobase-pair DNA of a malarial parasite. Molecular and Biochemical Parasitology 35, 97–107 (1989). doi:10.1016/0166-6851(89)90112–6
Wilson, R. J. M., Fry, M., et al. Subcellular fractionation of the two organelle DNAs of malaria parasites. Current Genetics 21, 405–408 (1992). doi: 10.1007/BF00351702
Wilson, R. J. M., Gardner, M. J., et al. Have malaria parasites three genomes? Parasitology Today 7, 134–136 (1991). doi:10.1016/0169–4758(91)90276–T
Wilson, R. J. M., Denny, P. W., et al. Complete gene map of the plastid-like DNA of the malaria parasite Plasmodium falciparum. Journal of Molecular Biology 261, 155–172 (1996). doi:10.1006/jmbi.1996.0449










