



 Figure 1
Figure 1















« Prev Next »
Genetics owes a great debt to good drawing. Handmade graphic depictions of mitotic chromosomes by Walther Flemming (Figure 1) and meiotic chromosomes by Walter Sutton provided an early record of the physical path of chromosomes during cell division. The physical movement of chromosomes could then be correlated with cells' patterns of genetic inheritance. (The idea that genes were carried on cytological structures is now known as the chromosome theory.) Using such methods, researchers determined that although mitosis and meiosis are both forms of cell division, the results of these processes are actually quite different.
Gene Transmission in Mitosis
Mitosis occurs in somatic cells; this means that it takes place in all types of cells that are not involved in the production of gametes. Prior to each mitotic division, a copy of every chromosome is created; thus, following division, a complete set of chromosomes is found in the nucleus of each new cell. Indeed, apart from random mutations, each successive duplicate cell will have the same genetic composition as its parent, due to the inheritance of the same chromosome set and similar biological environment. This works well for replacing damaged tissue or for growth and expansion from an embryonic state.
Because the genes contained in the duplicate chromosomes are transferred to each successive cellular generation, all mitotic progeny are genetically similar. However, there are exceptions. For example, there are genetic variations that arise in clonal species, such as bacteria, due to spontaneous mutations during mitotic division. Furthermore, chromosomes are sometimes replicated multiple times without any accompanying cell division. This occurs in the cells of Drosophila larvae salivary glands, for example, where there is a high metabolic demand. The chromosomes there are called polytene chromosomes, and they are extremely large compared to chromosomes in other Drosophila cells. These chromosomes replicate by undergoing the initial phases of mitosis without any cytokinesis (Figure 2). Therefore, the same cell contains thick arrangements of duplicate chromosomes side by side, which look like strands of very thick rope. Scientists believe that these chromosomes are hyper-replicated to allow for the rapid and copious production of certain proteins that help larval growth and metamorphosis (Gilbert, 2008).
Gene Transmission in Meiosis

Figure 2: Examples of polytene chromosomes
Pairing of homologous chromatids results in hundreds to thousands of individual chromatid copies aligned tightly in parallel to produce giant, "polytene" chromosomes.
© 2007 Nature Publishing Group Novikov, D. et al. High-pressure treatment of polytene chromosomes improves structural resolution. Nature Methods 4, 483 (2007). All rights reserved. 
The greatest impact of Sutton's work has far more to do with providing evidence for Mendel's principle of independent assortment than anything else. Specifically, Sutton saw that the position of each chromosome at the midline during metaphase was random, and that there was never a consistent maternal or paternal side of the cell division. Therefore, each chromosome was independent of the other. Thus, when the parent cell separated into gametes, the set of chromosomes in each daughter cell could contain a mixture of the parental traits, but not necessarily the same mixture as in other daughter cells.
To illustrate this concept, consider the variety derived from just three hypothetical chromosome pairs, as shown in the following example (Hirsch, 1963). Each pair consists of two homologues: one maternal and one paternal. Here, capital letters represent the maternal chromosome, and lowercase letters represent the paternal chromosome:
- Pair 1: A and a
- Pair 2: B and b
- Pair 3: C and c
When these chromosome pairs are reshuffled through independent assortment, they can produce eight possible combinations in the resulting gametes:
- A B C
- A B c
- A b c
- A b C
- a B C
- a B c
- a b C
- a b c
A mathematical calculation based on the number of chromosomes in an organism will also provide the number of possible combinations of chromosomes for each gamete. In particular, Sutton pointed out that the independence of each chromosome during meiosis means that there are 2n possible combinations of chromosomes in gametes, with "n" being the number of chromosomes per gamete. Thus, in the previous example of three chromosome pairs, the calculation is 23, which equals 8. Furthermore, when you consider all the possible pairings of male and female gametes, the variation in zygotes is (2n)2, which results in some fairly large numbers.
But what about chromosome reassortment in humans? Humans have 23 pairs of chromosomes. That means that one person could produce 223 different gametes. In addition, when you calculate the possible combinations that emerge from the pairing of an egg and a sperm, the result is (223)2 possible combinations. However, some of these combinations produce the same genotype (for example, several gametes can produce a heterozygous individual). As a result, the chances that two siblings will have the same combination of chromosomes (assuming no recombination) is about (3/8)23, or one in 6.27 billion. Of course, there are more than 23 segregating units (Hirsch, 2004).
While calculations of the random assortment of chromosomes and the mixture of different gametes are impressive, random assortment is not the only source of variation that comes from meiosis. In fact, these calculations are ideal numbers based on chromosomes that actually stay intact throughout the meiotic process. In reality, crossing-over between chromatids during prophase I of meiosis mixes up pieces of chromosomes between homologue pairs, a phenomenon called recombination. Because recombination occurs every time gametes are formed, we can expect that it will always add to the possible genotypes predicted from the 2n calculation. In addition, the variety of gametes becomes even more unpredictable and complex when we consider the contribution of gene linkage. Some genes will always cosegregate into gametes if they are tightly linked, and they will therefore show a very low recombination rate. While linkage is a force that tends to reduce independent assortment of certain traits, recombination increases this assortment. In fact, recombination leads to an overall increase in the number of units that assort independently, and this increases variation.
While in mitosis, genes are generally transferred faithfully from one cellular generation to the next; in meiosis and subsequent sexual reproduction, genes get mixed up. Sexual reproduction actually expands the variety created by meiosis, because it combines the different varieties of parental genotypes. Thus, because of independent assortment, recombination, and sexual reproduction, there are trillions of possible genotypes in the human species.
Aberrations That Alter Chromosome Number
During cell division, chromosomes sometimes disappear. This occurs when there is some aberration in the centromere, and spindle fibers cannot attach to the chromosome to segregate it to distal poles of the cell. Consequently, the lost chromosome never properly groups with others into a new nuclear envelope, and it is left in the cytoplasm, where it will not be transcribed. Also, chromosomes don't always separate equally into daughter cells. This sometimes happens in mitosis, when sister chromatids fail to separate during anaphase. One daughter cell thus ends up with more chromosomes in its nucleus than the other. Likewise, abnormal separation can occur in meiosis when homologous pairs fail to separate during anaphase I. This also results in daughter cells with different numbers of chromosomes. The phenomenon of unequal separation in meiosis is called nondisjunction. If nondisjunction causes a missing chromosome in a haploid gamete, the diploid zygote it forms with another gamete will contain only one copy of that chromosome from the other parent, a condition known as monosomy. Conversely, if nondisjunction causes a homologous pair to travel together into the same gamete, the resulting zygote will have three copies, a condition known as trisomy (Figure 3). The term "aneuploidy" applies to any of these conditions that cause an unexpected chromosome number in a daughter cell.
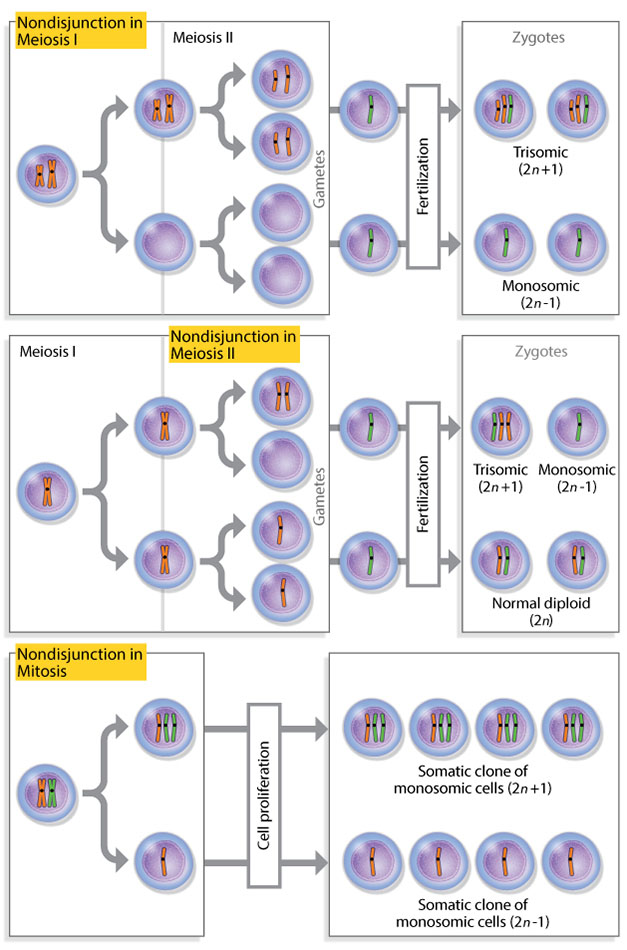
Figure 3: Nondisjunction results in daughter cells with unusual chromosome numbers.
Nondisjunction, in which chromosomes fail to separate equally, can occur in meiosis I (first row), meiosis II (second row), and mitosis (third row). These unequal separations can produce daughter cells with unexpected chromosome numbers, called aneuploids. When a haploid gamete does not receive a chromosome during meiosis as a result of nondisjunction, it combines with another gamete to form a monosomic zygote. When a gamete receives a complete homologous chromosome pair as a result of nondisjunction, it combines with another gamete to form a trisomic zygote.
© 2014 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved.
© 2014 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved.
Aneuploidy can also occur in humans. For instance, the underlying causes of Klinefelter's syndrome and Turner's syndrome are errors in sex chromosome number, and Down syndrome is caused by trisomy of chromosome 21. However, the severity of phenotypic abnormalities can vary among different types of aneuploidy. In addition, aneuploidy is rarely transferred to subsequent generations, because this condition impairs the production of gametes.
Overall, the inheritance of odd chromosome number arises from errors in segregation during chromosome replication. Often, it is these very exceptions or modifications of expected patterns in mitosis and meiosis that enrich our understanding of how the transfer of chromosomes is regulated from one generation to the next.
References and Recommended Reading
Belling, J., & Blakeslee, A. F. On the attachment of non-homologous chromosomes at the reduction division in certain 25-chromosome daturas. Proceedings of the National Academy of Sciences 12, 7–11 (1926)
Farmer, J. B., & Moore, J. E. S. On the maiotic phase (reduction divisions) in animals and plants. Quarterly Journal of Microscopical Science 48, 489–557 (1905)
Gilbert, S. F. Developmental Biology Companion Website (Sunderland, MA, Sinauer, 2010)
Hirsch, J. Behavior genetics and individuality understood. Science 42, 1436–1442 (1963) doi:10.1126/science.142.3598.1436
---. Uniqueness, diversity, similarity, repeatability, and heritability. International Journal of Comparative Psychology 17, 304–314 (2004)
Paweletz, N. Walther Flemming: Pioneer of mitosis research. Nature Reviews Molecular Cell Biology 2, 72–75 (2001) doi:10.1038/35048077 (link to article)
Pierce, B. Genetics: A Conceptual Approach (W.H. Freeman, New York, 2005)










