


















« Prev Next »
Scientists have devoted decades of effort to understanding how deoxyribonucleic acid (DNA) replicates itself. In simple terms, replication involves use of an existing strand of DNA as a template for the synthesis of a new, identical strand. American enzymologist and Nobel Prize winner Arthur Kornberg compared this process to a tape recording of instructions for performing a task: "[E]xact copies can be made from it, as from a tape recording, so that this information can be used again and elsewhere in time and space" (Kornberg, 1960).
In reality, the process of replication is far more complex than suggested by Kornberg's analogy. Researchers typically utilize simple bacterial cells in their experiments, but they still do not have all the answers, particularly when it comes to eukaryotic replication. Nonetheless, scientists are familiar with the basic steps in the replication process, and they continue to rely on this information as the basis for continued research and experimentation.
The Molecular Machinery of Bacterial DNA Replication
A typical bacterial cell has anywhere from about 1 million to 4 million base pairs of DNA, compared to the 3 billion base pairs in the genome of the common house mouse (Mus musculus). Still, even in bacteria, with their smaller genomes, DNA replication involves an incredibly sophisticated, highly coordinated series of molecular events. These events are divided into four major stages: initiation, unwinding, primer synthesis, and elongation.
Initiation and Unwinding
During initiation, so-called initiator proteins bind to the replication origin, a base-pair sequence of nucleotides known as oriC. This binding triggers events that unwind the DNA double helix into two single-stranded DNA molecules. Several groups of proteins are involved in this unwinding (Figure 1). For example, the DNA helicases are responsible for breaking the hydrogen bonds that join the complementary nucleotide bases to each other; these hydrogen bonds are an essential feature of James Watson and Francis Crick's three-dimensional DNA model. Because the newly unwound single strands have a tendency to rejoin, another group of proteins, the single-strand-binding proteins, keep the single strands stable until elongation begins. A third family of proteins, the topoisomerases, reduce some of the torsional strain caused by the unwinding of the double helix.
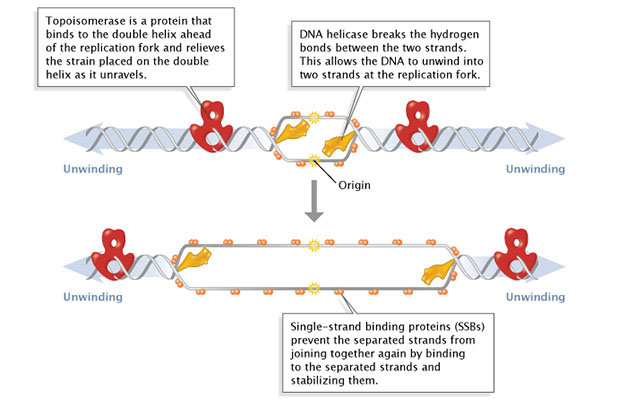
Figure 1: Facilitation of DNA unwinding.
During DNA replication, several proteins facilitate the unwinding of the DNA double helix into two single strands. Topoisomerases (red) reduce torsional strain caused by the unwinding of the DNA double helix; DNA helicase (yellow) breaks hydrogen bonds between complementary base-pairs; single-strand binding proteins (SSBs) stabilize the separated strands and prevent them from rejoining.
© 2014 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved. 
As previously mentioned, the location at which a DNA strand begins to unwind into two separate single strands is known as the origin of replication. As shown in Figure 1, when the double helix unwinds, replication proceeds along the two single strands at the same time but in opposite directions (i.e., left to right on one strand, and right to left on the other). This forms two replication forks that move along the DNA, replicating as they go.
Primer Synthesis
Primer synthesis marks the beginning of the actual synthesis of the new DNA molecule. Primers are short stretches of nucleotides (about 10 to 12 bases in length) synthesized by an RNA polymerase enzyme called primase. Primers are required because DNA polymerases, the enzymes responsible for the actual addition of nucleotides to the new DNA strand, can only add deoxyribonucleotides to the 3'-OH group of an existing chain and cannot begin synthesis de novo. Primase, on the other hand, can add ribonucleotides de novo. Later, after elongation is complete, the primer is removed and replaced with DNA nucleotides.
Elongation
Finally, elongation--the addition of nucleotides to the new DNA strand--begins after the primer has been added. Synthesis of the growing strand involves adding nucleotides, one by one, in the exact order specified by the original (template) strand. Recall that one of the key features of the Watson-Crick DNA model is that adenine is always paired with thymine and cytosine is always paired with guanine. So, for example, if the original strand reads A-G-C-T, the new strand will read T-C-G-A.
DNA is always synthesized in the 5'-to-3' direction, meaning that nucleotides are added only to the 3' end of the growing strand. As shown in Figure 2, the 5'-phosphate group of the new nucleotide binds to the 3'-OH group of the last nucleotide of the growing strand. Scientists have yet to identify a polymerase that can add bases to the 5' ends of DNA strands.
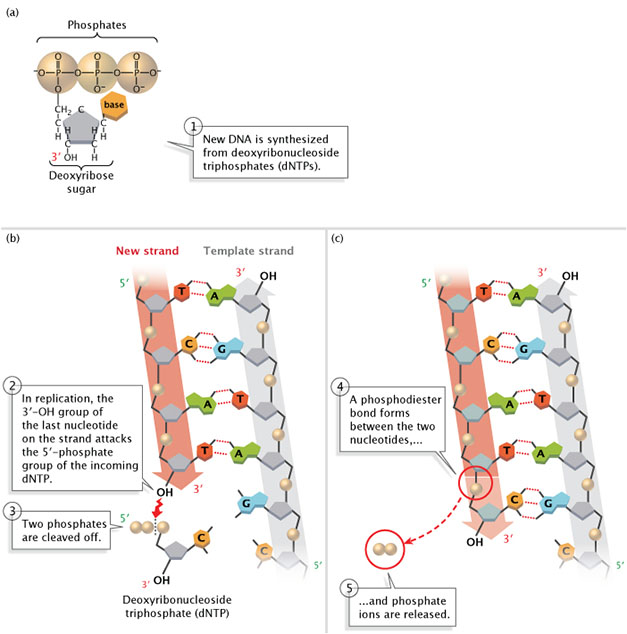
Figure 2: New DNA is synthesized from deoxyribonucleoside triphosphates (dNTPs).
(A) A deoxyribonucleoside triphosphate (dNTP). (B) During DNA replication, the 3'-OH group of the last nucleotide on the new strand attacks the 5'-phosphate group of the incoming dNTP. Two phosphates are cleaved off. (C) A phosphodiester bond forms between the two nucleotides, and phosphate ions are released.
© 2014 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved.
The Discovery of DNA Polymerase
While studying E. coli bacteria, enzymologist Arthur Kornberg discovered that DNA polymerases catalyze DNA synthesis. Kornberg's experiment involved mixing all of the basic "ingredients" necessary for E. coli DNA synthesis in a test tube, including nucleotides, E. coli extract, and ATP, and then purifying and testing the enzymes involved. Using this method, Kornberg not only discovered DNA polymerases, but he also performed some of the initial work demonstrating how enzymes add new nucleotides to growing DNA chains (Kornberg, 1959).
Scientists have since identified a total of five different DNA polymerases in E. coli, each with a specialized role. For example, DNA polymerase III does most of the elongation work, adding nucleotides one by one to the 3' end of the new and growing single strand. Other enzymes, including DNA polymerase I and RNase H, are responsible for removing the RNA primer after DNA polymerase III has begun its work, replacing it with DNA nucleotides (Ogawa & Okazaki, 1984). When these enzymes finish, they leave a nick between the section of DNA that was formerly the primer and the elongated section of DNA. Another enzyme called DNA ligase then acts to seal the bond between the two adjacent nucleotides.
DNA Polymerase Only Moves in One Direction
After a primer is synthesized on a strand of DNA and the DNA strands unwind, synthesis and elongation can proceed in only one direction. As previously mentioned, DNA polymerase can only add to the 3' end, so the 5' end of the primer remains unaltered. Consequently, synthesis proceeds immediately only along the so-called leading strand. This immediate replication is known as continuous replication. The other strand (in the 5' direction from the primer) is called the lagging strand, and replication along it is called discontinuous replication. The double helix has to unwind a bit before the synthesis of another primer can be initiated further up on the lagging strand. Synthesis can then occur from the 3' end of that new primer. Next, the double helix unwinds a bit more, and another spurt of replication proceeds. As a result, replication along the lagging strand can only proceed in short, discontinuous spurts (Figure 3).
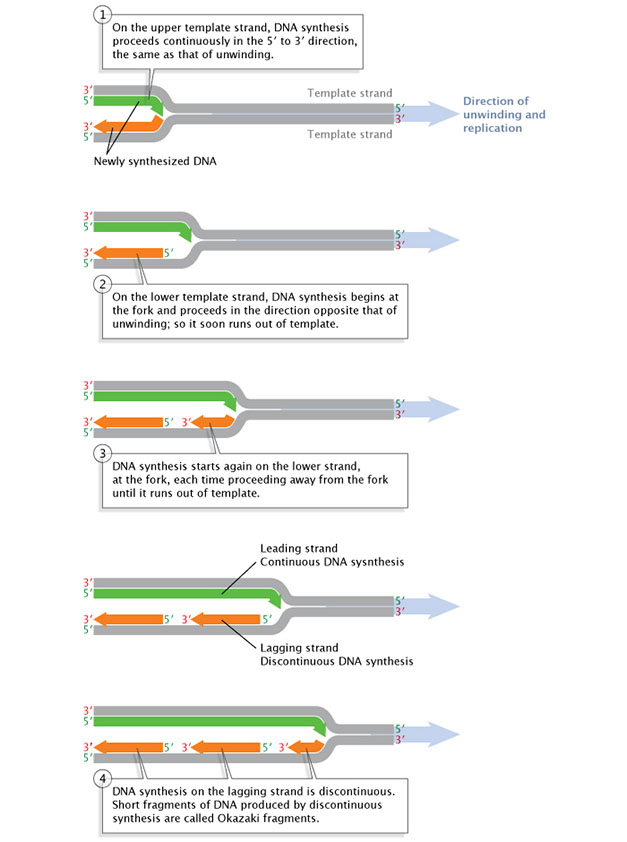
Figure 3: Replication of the leading DNA strand is continuous, while replication along the lagging strand is discontinuous.
After a short length of the DNA has been unwound, synthesis must proceed in the 5' to 3' direction; that is, in the direction opposite that of the unwinding.
© 2014 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved.
The fragments of newly synthesized DNA along the lagging strand are called Okazaki fragments, named in honor of their discoverer, Japanese molecular biologist Reiji Okazaki. Okazaki and his colleagues made their discovery by conducting what is known as a pulse-chase experiment, which involved exposing replicating DNA to a short "pulse" of isotope-labeled nucleotides and then varying the length of time that the cells would be exposed to nonlabeled nucleotides. This later period is called the "chase" (Okazaki et al., 1968). The labeled nucleotides were incorporated into growing DNA molecules only during the initial few seconds of the pulse; thereafter, only nonlabeled nucleotides were incorporated during the chase. The scientists then centrifuged the newly synthesized DNA and observed that the shorter chases resulted in most of the radioactivity appearing in "slow" DNA. The sedimentation rate was determined by size: smaller fragments precipitated more slowly than larger fragments because of their lighter weight. As the investigators increased the length of the chases, radioactivity in the "fast" DNA increased with little or no increase of radioactivity in the slow DNA. The researchers correctly interpreted these observations to mean that, with short chases, only very small fragments of DNA were being synthesized along the lagging strand. As the chases increased in length, giving DNA more time to replicate, the lagging strand fragments started integrating into longer, heavier, more rapidly sedimenting DNA strands. Today, scientists know that the Okazaki fragments of bacterial DNA are typically between 1,000 and 2,000 nucleotides long, whereas in eukaryotic cells, they are only about 100 to 200 nucleotides long.
The Challenges of Eukaryotic Replication
Bacterial and eukaryotic cells share many of the same basic features of replication; for instance, initiation requires a primer, elongation is always in the 5'-to-3' direction, and replication is always continuous along the leading strand and discontinuous along the lagging strand. But there are also important differences between bacterial and eukaryotic replication, some of which biologists are still actively researching in an effort to better understand the molecular details. One difference is that eukaryotic replication is characterized by many replication origins (often thousands), not just one, and the sequences of the replication origins vary widely among species. On the other hand, while the replication origins for bacteria, oriC, vary in length (from about 200 to 1,000 base pairs) and sequence, except among closely related organisms, all bacteria nonetheless have just a single replication origin (Mackiewicz et al., 2004).
Eukaryotic replication also utilizes a different set of DNA polymerase enzymes (e.g., DNA polymerase δ and DNA polymerase ε instead of DNA polymerase III). Scientists are still studying the roles of the 13 eukaryotic polymerases discovered to date. In addition, in eukaryotes, the DNA template is compacted by the way it winds around proteins called histones. This DNA-histone complex, called a nucleosome, poses a unique challenge both for the cell and for scientists investigating the molecular details of eukaryotic replication. What happens to nucleosomes during DNA replication? Scientists know from electron micrograph studies that nucleosome reassembly happens very quickly after replication (the reassembled nucleosomes are visible in the electron micrograph images), but they still do not know how this happens (Annunziato, 2005).
Also, whereas bacterial chromosomes are circular, eukaryotic chromosomes are linear. During circular DNA replication, the excised primer is readily replaced by nucleotides, leaving no gap in the newly synthesized DNA. In contrast, in linear DNA replication, there is always a small gap left at the very end of the chromosome because of the lack of a 3'-OH group for replacement nucleotides to bind. (As mentioned, DNA synthesis can proceed only in the 5'-to-3' direction.) If there were no way to fill this gap, the DNA molecule would get shorter and shorter with every generation. However, the ends of linear chromosomes—the telomeres—have several properties that prevent this.
DNA replication occurs during the S phase of cell division. In E. coli, this means that the entire genome is replicated in just 40 minutes, at a pace of approximately 1,000 nucleotides per second. In eukaryotes, the pace is much slower: about 40 nucleotides per second. The coordination of the protein complexes required for the steps of replication and the speed at which replication must occur in order for cells to divide are impressive, especially considering that enzymes are also proofreading, which leaves very few errors behind.
Summary
The study of DNA replication started almost as soon as the structure of DNA was elucidated, and it continues to this day. Currently, the stages of initiation, unwinding, primer synthesis, and elongation are understood in the most basic sense, but many questions remain unanswered, particularly when it comes to replication of the eukaryotic genome. Scientists have devoted decades to the study of replication, and researchers such as Kornberg and Okazaki have made a number of important breakthroughs. Nonetheless, much remains to be learned about replication, including how errors in this process contribute to human disease.
References and Recommended Reading
Annunziato, A. T. Split decision: What happens to nucleosomes during DNA replication? Journal of Biological Chemistry 280, 12065–12068 (2005)
Bessman, M. J., et al. Enzymatic synthesis of deoxyribonucleic acid. II. General properties of the reaction. Journal of Biological Chemistry 233, 171–177 (1958)
Kornberg, A. The biological synthesis of deoxyribonucleic acid. Nobel Lecture, December 11, 1959. (link to transcript)
———. Biological synthesis of deoxyribonucleic acid. Science 131, 1503–1508 (1960)
Lehman, I. R., et al. Enzymatic synthesis of deoxyribonucleic acid. I. Preparation of substrates and partial purification of an enzyme from Escherichia coli. Journal of Biological Chemistry 233, 163–170 (1958)
Losick, R., & Shapiro, L. DNA replication: Bringing the mountain to Mohammed. Science 282, 1430–1431 (1998)
Mackiewicz, P., et al. Where does bacterial replication start? Rules for predicting the oriC region. Nucleic Acids Research 32, 3781–3791 (2004)
Ogawa, T., & Okazaki, T. Function of RNase H in DNA replication revealed by RNase H defective mutants of Escherichia coli. Molecular and General Genetics 193, 231–237 (1984)
Okazaki, R., et al. Mechanism of DNA chain growth. I. Possible discontinuity and unusual secondary structure of newly synthesized chains. Proceedings of the National Academy of Sciences 59, 598–605 (1968)










