









« Prev Next »

Cell Adhesion and Cell Communication
All cells rely on cell signaling to detect and respond to cues in their environment. This process not only promotes the proper functioning of individual cells, but it also allows communication and coordination among groups of cells — including the cells that make up organized communities called tissues. Because of cell signaling, tissues have the ability to carry out tasks no single cell could accomplish on its own.
Different types of tissues, such as bone, brain, and the lining of the gut, have characteristic features related to the number and types of cells they contain. Cell spacing is also critical to tissue function, so this geometry is precisely regulated. To preserve proper tissue architecture, adhesive molecules help maintain contact between nearby cells and structures, and tiny tunnel-like junctions allow the passage of ions and small molecules between adjacent cells. Meanwhile, signaling molecules relay positional information among the cells in a tissue, as well as between these cells and the extracellular matrix. These signaling pathways are critical to maintaining the state of equilibrium known as homeostasis within a tissue. For example, the processes involved in wound healing depend on positional information in order for normal tissue architecture to be restored. Such positional signals are also crucial for the development of adult structures in multicellular organisms. As tissues develop, clumps of unorganized cells grow and sort themselves according to signals they send and receive.
How Do Integrins Promote Tissue Structure and Function?
Within tissues, adhesive molecules allow cells to maintain contact with one another and with structures in the extracellular matrix. One especially important class of adhesive molecules is the integrins. Integrins are more than just mechanical links, however: They also relay signals both to and from cells. In this way, integrins play an important role in sensing the environment and controlling cell shape and motility.
Integrins are a diverse family of transmembrane proteins found in all animal cells. Even simple animals like sponges have these proteins. Each individual integrin consists of two main parts: an alpha subunit and a beta subunit. Variation in the alpha and beta subunits accounts for the wide variety of integrins observed throughout the animal kingdom. For example, humans alone have over 20 different kinds of integrins.
Integrins link the actin cytoskeleton of a cell to various external structures. The cytoplasmic portion of each integrin molecule binds to adaptor proteins that connect to the actin filaments inside the cell. The extracellular portion of the integrin then binds to molecules in the extracellular matrix or on the surface of other cells. Integrin attachments to neighboring cells can break and reform as a cell moves (Figure 1).
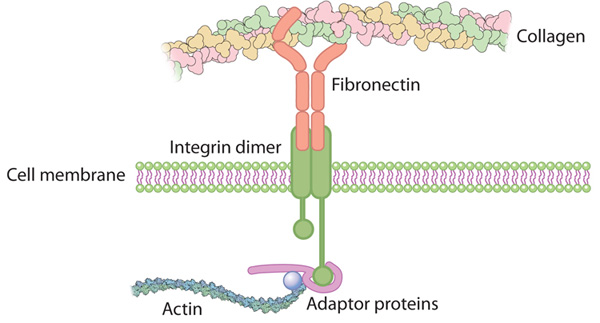
Figure 1: Integrin connects the extracellular matrix with the actin cytoskeleton inside the cell.
© 2010 Nature Education All rights reserved. 
How Else Do the Cells within a Tissue Stay in Contact?
Beyond integrins, cells rely on several other adhesive proteins to maintain physical contact. As an example, consider the epithelial cells that line the inner and outer surfaces of the human body — including the skin, intestines, airway, and reproductive tract. These cells provide a dramatic example of the different kinds of cell-to-cell junctions, but the same junctions also exist in a wide range of other tissues.
The side surfaces of epithelial cells are tightly linked to those of neighboring cells, forming a sheet that acts as a barrier. Within this sheet, each individual cell has a set orientation. Through integrins, the basal end of each cell connects to a specialized layer of extracellular matrix called the basal lamina. In contrast, the apical end of each cell faces out into the environment — such as the inner cavity or lumen of the gut.
The side-to-side junctions that link epithelial cells are diverse in their protein makeup and function. The adhesive transmembrane proteins anchoring these junctions have extracellular portions that interact with similar proteins on adjacent cells. Protein complexes within each cell further connect the transmembrane adhesive proteins to the cytoskeleton. In particular, adaptor complexes bind adherens junctions to cytoskeletal actin, and other adaptor complexes bind desmosomes to intermediate filaments. Both of these types of junctional complexes provide cells and tissues with mechanical support, and they additionally recruit intracellular signaling molecules to relay positional information to the nucleus.
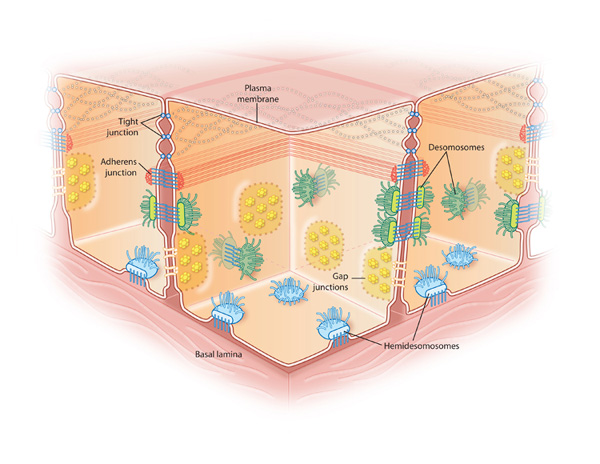
Figure 2: The different types of cell junctions
Tight junctions (blue dots) between cells are connected areas of the plasma membrane that stitch cells together. Adherens junctions (red dots) join the actin filaments of neighboring cells together. Desmosomes are even stronger connections that join the intermediate filaments of neighboring cells. Hemidesmosomes (light blue) connect intermediate filaments of a cell to the basal lamina, a combination of extracellular molecules on other cell surfaces. Gap junctions (yellow) are clusters of channels that form tunnels of aqueous connectivity between cells.
© 2010 Nature Education All rights reserved.
The lateral surfaces of epithelial cells also contain several other types of specialized junctions. Tight junctions form a seal between cells that is so strong that not even ions can pass across it. Gap junctions are involved in cellular communication — not just in epithelial tissue, but in other tissue types as well. Gap junctions are specialized connections that form a narrow pore between adjacent cells. These pores permit small molecules and ions to move from one cell to another. In this way, gap junctions provide metabolic and electrical coupling between cells. For example, cardiac tissue has extensive gap junctions, and the rapid movement of ions through these junctions helps the tissue beat in rhythm. Gap junctions may also open and close in response to metabolic signals (Figure 2, Figure 3).

Figure 3: A gap junction
In a gap junction, the lipid bilayer of adjacent cells is pierced through by proteins called connexons. These proteins group together and effectively form a group of communication tunnels between adjacent cells.
© 2010 Nature Education All rights reserved.
Cell Death Can Be Prompted by a Signal
Cell signaling isn't just central to tissue architecture and function: It also plays an important role in the balance between cell growth and death. Although it sounds like a bad thing, apoptosis — or the process of programmed cell death — is an essential aspect of development. Without it, repair and replenishment processes would overrun tissues with new cells. The orderly demise of a certain proportion of cells is therefore necessary for normal tissue turnover and maintenance of homeostasis. Apoptosis is distinct from necrosis, a messier form of cell death that causes cells to literally swell and burst. Necrotic cell death is not programmed; rather, it occurs in response to trauma or injury.
A range of extracellular and intracellular signals can trigger either cell growth or apoptosis. When cells receive these signals from their neighbors or from other aspects of the external environment, they carefully weigh them against each other before choosing a course of action. For instance, signals that indicate a lack of nutrients or the presence of toxins would likely stall cell growth and promote apoptosis. Within the cell, damage to the DNA or loss of mitochondrial integrity might also result in programmed cell death.
Cells self-destruct cleanly and quickly during apoptosis, thanks to the activation of a variety of enzymes — proteases and nucleases — that break down proteins and nucleic acids, respectively. In fact, scientists look for a characteristic pattern of fragmentation and nuclear condensation within tissues as evidence that apoptosis has occurred.
Conclusion
Some cell signaling occurs on a local level, such as when cells interact with the surrounding extracellular matrix or with their immediate neighbors. This type of signaling is especially important to the structure and function of tissues. Various signaling molecules allow the cells within a tissue to share information about internal and external conditions. This information helps the cells arrange themselves, coordinate their functions, and even know when to grow and when to die. Some of these signaling molecules also function in an adhesive capacity — not just relaying messages between the cells in a tissue, but physically joining these cells to one another.
eBooks
This page appears in the following eBook











