



















« Prev Next »
Historical Perspective
Throughout the history of ecology, scientists have observed variability across time and space in the abiotic and biotic components of ecosystems. But early ecologists did not have the technology or concepts to explicitly deal with spatial heterogeneity, so there was a tendency to develop explanations by grouping organisms into uniform and recognizable units. For example, scientists were struck by the relatively consistent associations of plant species and grouped vegetation into community types (Mueller-Dombois & Ellenberg 1974). Compared to vegetation, where observed change was rather slow, observations of fluctuating populations ranging from bacteria and protozoans in the laboratory to snowshoe hares (Lepus americanus) in the boreal forest, led scientists to mathematical theories that explicitly focused on temporal dynamics (Kingsland 1995). But the resulting models treated the environment as spatially homogeneous. Such views of nature and the theory about dynamics led to “equilibrium” concepts (May 1973) that dominated ecological thinking from the 1920s through the 1980s.
During the 1980s, advances in the accessibility of computing, remotely sensed satellite and aerial imagery, development of geographic information systems (GIS, ARC/INFO was first released in 1982), and spatial statistical methods (Fortin & Dale 2005), enabled ecologists to observe and analyze spatial heterogeneity ranging from local habitats to entire continents. The technology enhanced a paradigm shift occurring in ecology and the emergence of landscape ecology as a sub-discipline within ecology (Wu & Loucks 1995). Landscape ecology specifically recognizes that disturbance, whether anthropogenic or caused by natural processes, creates spatial heterogeneity that is the normal condition of ecosystems. In landscape ecology particularly, a “non-equilibrium” view emerged, that links disturbance in time and space to system structure and function in feedback loops that influence the ecology and evolutionary trajectories in the ecosystems. The International Association of Landscape Ecology was formed in 1982. In 1986, Forman and Godron published their seminal text on landscape ecology. This work was important, not only because it outlined principles, but also because it brought together the North American scientific interest — typically focused on heterogeneity in ecosystems — with more anthropocentric scientific traditions of geography, landscape architecture, and planning, rooted in the long history of landscape alteration in Europe.
During the 1980s, advances in the accessibility of computing, remotely sensed satellite and aerial imagery, development of geographic information systems (GIS, ARC/INFO was first released in 1982), and spatial statistical methods (Fortin & Dale 2005), enabled ecologists to observe and analyze spatial heterogeneity ranging from local habitats to entire continents. The technology enhanced a paradigm shift occurring in ecology and the emergence of landscape ecology as a sub-discipline within ecology (Wu & Loucks 1995). Landscape ecology specifically recognizes that disturbance, whether anthropogenic or caused by natural processes, creates spatial heterogeneity that is the normal condition of ecosystems. In landscape ecology particularly, a “non-equilibrium” view emerged, that links disturbance in time and space to system structure and function in feedback loops that influence the ecology and evolutionary trajectories in the ecosystems. The International Association of Landscape Ecology was formed in 1982. In 1986, Forman and Godron published their seminal text on landscape ecology. This work was important, not only because it outlined principles, but also because it brought together the North American scientific interest — typically focused on heterogeneity in ecosystems — with more anthropocentric scientific traditions of geography, landscape architecture, and planning, rooted in the long history of landscape alteration in Europe.
Terminology and Concepts
Imaging and mapping technology naturally promoted a patch-corridor-matrix approach to landscape ecology. Examining the map of an area in North Dakota (Figure 1) helps to define important vocabulary and illustrates some typical questions studied by landscape ecologists. A patch is an area of habitat differing from its surroundings, often the smallest ecologically distinct landscape feature in a landscape mapping and classification system. In Figure 1, wetlands and perennial grasslands would likely be patches of focal interest for the study of ecological processes. The matrix is the majority of the surrounding landscape (i.e., not the patches); in this case the matrix primarily consists of fields of agricultural crops. The large proportion of the landscape classified as matrix may have profound influences on the ecological processes in the landscape; for example, consider the flow of pesticides from the farmed matrix to the wetland patches in Figure 1. Finally, corridors are narrow patches that may act as links or barriers in a landscape. Beyond the image of narrow patches, corridors are functionally important landscape structures influencing dispersal of plants and animals in the landscape (Haddad et al. 2003).
Composition is the relative proportion of habitat types in the landscape, regardless of spatial distribution; for example, in Figure 1 perennial grasslands comprise 17%, wetlands about 18%, and croplands about 56% of the landscape (Phillips et al. 2002). Configuration refers to almost limitless aspects of landscape heterogeneity, especially the physical and spatial distribution of landscape elements. Configuration metrics that apply across an entire landscape would include characteristics such as the dendritic pattern of streams in a watershed, or the diversity of habitat types. Ecologists have been particularly interested in how the spatial distribution of elements affects ecological processes. For example, “is the percentage of wetlands occupied by tiger salamanders (Ambystoma tigrinum) influenced by whether wetlands are distributed randomly or clumped to some degree?” (Figure 1). Another sense of configuration focuses on the characteristics of patches (e.g., the size, shape, or perimeter to area ratio [P/A] of individual patches). This latter emphasis is often prompted by interest in a particular organism that uses a patch type as habitat (e.g., grassland birds nesting in the landscape) (Figure 1). P/A is a measure of the amount of edge (or ecotone) between a focal patch type and the surrounding matrix. Ecotones represent transitional zones that often have important influences on ecological processes. For instance in Figure 1, landscape ecologists have asked, “do predators search for nests in the same patterns along edges compared to interiors of grassland patches?” (Phillips et al. 2002).
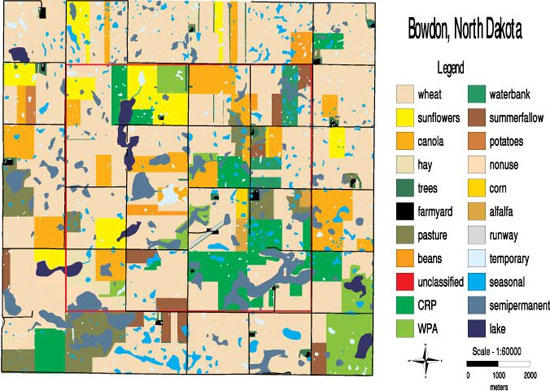
Figure 1: A map of classified habitats in the landscape near Bowdon, North Dakota, USA
Patches of interest include the wetlands (blue) and perennial grasslands (green) surrounded by a matrix of agricultural crops (primarily tan, yellow and orange). Note the gravel roads (black) spaced one mile apart. The red box encloses 16 mi2.
© 2010 Nature Education Courtesy of William Clark & Michael Phillips. All rights reserved. 
There are many other terms (Forman & Godron 1986) and metrics (McGarigal & Marks 1995) that are used to describe landscapes, but two others are particularly important to understand. In an ecological sense scale is the resolution at which an organism perceives its environment (e.g., consider a mouse population versus a moose population). Scale is important to plants too; “what is the detectable limit over which seeds disperse from a plant?” Often scale is also used in a technological sense referring to the smallest area that can be resolved into a single type on the landscape. Spatial scale of remotely sensed vegetation may be as small as 0.5 m, although for large areas it is more typically 30 m, or greater (see here). Extent is the extended range of study, or the area included within the landscape boundary, such as a national park or state.
Themes of Study and Application
The development and dynamics of spatial heterogeneity in landscapes is a central theme of ecological studies, especially the effects of conversion of natural ecosystems into human dominated systems such as agricultural or urban land use. As natural habitat is altered in a landscape (e.g., forest in Figure 2) both the composition (forest area) and the configuration (spatial pattern of patches) change. This conversion is called fragmentation (Figure 3). Evidence is mounting that change in composition has a dominant effect on composition of the biota, whereas variation in configuration has a lesser effect, except at very low proportion of patch composition in the landscape (Fahrig 1997). Such ideas have practical consequences for the conservation of biodiversity. Rhetorically, “will protection of a Single Large patch of habitat or Several Small patches (the SLOSS tradeoff; Simberloff & Abele 1976) have equivalent effects on species persistence or biodiversity?"

Figure 2: Clearcut logging in forests of the Pacific Northwest, USA
Notice that some cut patches are relatively recent, with bare ground exposed, whereas vegetation has begun to reestablish itself in others.
© 2010 Nature Education Courtesy of Marli Bryant Miller. All rights reserved.
Another theme relating configuration and conservation is the potential value of corridors linking habitats. Across large parts of the globe, biodiversity is affected by the interactions between climate changes and landscape linkages, and land use, which may block (barriers) or enhance (corridors) species persistence (Opdam & Wascher 2004). Although ecologists have debated the scientific evidence concerning the effect of corridors (Beier & Noss 1998), like SLOSS, the idea of landscape linkages has intuitive appeal for conservation. For example, conservationists interested in large predators have used knowledge of their movements, coupled with habitat data in GIS analyses, to identify focal areas for conservation corridors (Figure 4). Conversely, when the motivating question is related to the spread of an invasive insect like emerald ash borer (Agrilus planipennis), or an emerging infectious disease like avian influenza, then corridors and patches that act as stepping stones have negative rather than positive value (With 2002). Recent combinations of population genetics and landscape ecology have strengthened the ability to understand how species respond to landscape change, because they have the potential to reveal patterns of dispersal linkages and barriers across extensive regions (Manel et al. 2003). Landscape genetics provides a conceptual framework and tools to link the ecological short-term and local spatial scale with the long-term and large spatial scale helpful in understanding adaptation.
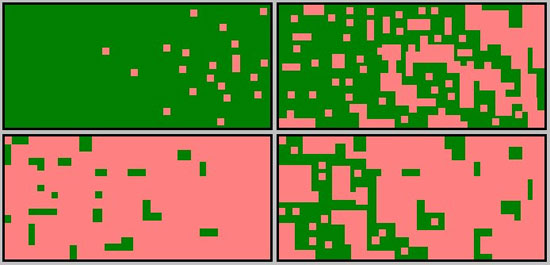
Figure 3: Process of landscape fragmentation
Fragmentation can be summarized in several different phases. Clockwise, from the upper left panel: (a) perforation (initial small openings), (b) dissection (larger intrusions of change, often along physical features), (c) dissipation (spread and coalescing of alteration), and eventually, (d) shrinkage (reduction of patch size), and attrition (loss of patches).
© 2010 Nature Education Courtesy of Michigan Forests Forever Teachers Guide. All rights reserved.
Although there has been much research emphasis on how organisms respond to landscape heterogeneity, it is important to recognize that organisms comprise the heterogeneity, and often cause it themselves. For example, herbivores as different as bison (Bison bison) (Knapp et al. 1999) and prairie dogs (Cynomys spp.) (Johnston 1995) influence large scale distribution of vegetation types and microhabitats in North American prairies. In natural systems, organisms may have the same ecosystem engineering effect as the pervasive influence of humans, although the effect of human influence has been the focus of far less attention in the development of landscape theory.
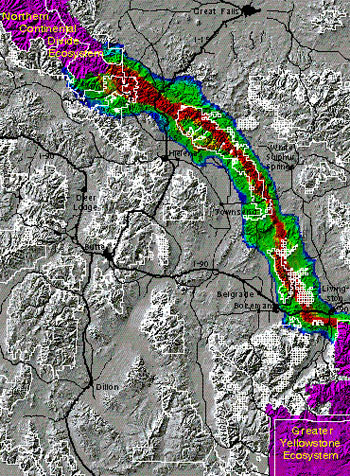
Figure 4: A corridor linking grizzly bear habitat in Montana, USA
This corridor was delineated based on grizzly bear habitat preferences and landscape structure. For conservation planning purposes it highlights the importance of protected national parks and forests.
© 2010 Nature Education Courtesy of Richard Walker & Lance Craighead. All rights reserved.
Ecosystem and landscape scientists share a common interest in how spatial heterogeneity affects abiotic and biotic processes in ecosystems (Turner 2005). Whether the compartments are abstract units of an ecosystem model or patches on a landscape, scientists are developing a unified framework to understand the dynamic flows of materials such as water, nutrients, and chemicals, both in time and space (Costanza et al. 2002). The ideas can be applied to suggesting strategies for managing soil fertility and erosion, nutrient cycling, and pollution. Solving such problems requires the consideration of continuous environmental gradients, requiring the integration of approaches used in geomorphology, surface analysis, and spatial statistics. This interest has led to the emergence of a dynamic mosaic paradigm (Cushman et al. 2010), which complements the patch-corridor-matrix paradigm. Landscape concepts regarding loss and fragmentation of vegetation cover around the world have become fundamental to understanding the carbon cycle, and predicting the consequences of global climate change (Houghton 1995).

Figure 5: Schwerin, Germany
The interdependence of natural systems with human socio-economic systems in the landscape is illustrated.
© 2010 Nature Education All rights reserved.
More than 75% of Earth’s ice-free land shows evidence of alteration as a result of human residence and land use (Ellis & Ramankutty 2008). Furthermore, the legacy of anthropogenic land use and natural events, like fire, is clearly important, and has measurable effects on ecosystem function, even hundreds of years after landscape alteration (Turner 2005). Even in the simple landscape shown in Figure 1 you can see the rectangular fields and spacing of the 1 mi2 sections that are a legacy of the Public Land Survey. This anthropogenic scaling is fundamental to most ecological processes in this landscape. Natural processes that affect the stability and resilience of ecosystems apply equally well in wild lands, and the vast majority of the biosphere where humans live (Figure 5). The challenge to landscape ecologists is to meld the ecology-centered spatial view with the society-centered holistic view to the benefit of understanding how spatial-temporal heterogeneity affects the resilience of the ecosystems of the earth on which all organisms depend (Wu 2006).
References and Recommended Reading
Beier, P. & Noss, R. F. Do habitat corridors provide connectivity? Conservation Biology 12, 1241–1252 (1998).
Costanza, R., Voinov, A. et al. Integrated ecological economic modeling of the Patuxent River watershed, Maryland. Ecological Monographs 72, 203–232 (2002).
Cushman, S. A., Evans, J. S. et al. Landscape ecology: past, present, and future. In Spatial Complexity, Informatics, and Wildlife Conservation, eds. S. A. Cushman & F. Huettmann(New York: Springer, 2010): 65–72.
Ellis, E. C. & Ramankutty, N. Putting people in the map: anthropogenic biomes of the world. Frontiers in Ecology and the Environment 6, 439–447 (2008).
Fahrig, L. Relative effects of habitat loss and fragmentation on population extinction. Journal of Wildlife Management 61, 603–610 (1997).
Forman, R. T. T. & Godron, M. Landscape Ecology. New York, NY: John Wiley & Sons, 1986.
Fortin, M. J. & Dale, M. R. T. Spatial Analysis: A Guide for Ecologists. New York, NY: Cambridge University Press, 2005.
Haddad, N. M., Bowne, D. R. et al. Corridor use by diverse taxa. Ecology 84, 609–615 (2003).
Houghton, R. A. Land-use change and the carbon cycle. Global Change Biology 1, 275–287 (1995).
Johnston, C. A. Effects of animals on landscape pattern. in Mosaic Landscapes and Ecological Processes, eds. L. Hansson et al. (London: Chapman and Hall, 1995): 57–80.
Kingsland, S. E. Modeling Nature. Chicago, IL: University of Chicago Press, 1995.
Knapp, A. K. et al. The keystone role of bison in North American tallgrass prairie. Bioscience 49, 39–50 (1999).
Manel, S. et al. Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology and Evolution 18, 189–197 (2003).
May, R. M. Stability and Complexity in Model Ecosystems. Princeton, NJ: Princeton University Press, 1973.
McGarigal, K. & Marks, B. J. FRAGSTATS, spatial pattern analysis program for quantifying landscape structure. In USDA Forest Service General Technical Report PNW-GTR-351 (1995).
Mueller-Dombois, D. & Ellenberg, H. Aims and Methods of Vegetation Ecology. New York, NY: John Wiley & Sons, 1974.
Opdam, P. & Wascher, D. Climate change meets habitat fragmentation: linking landscape and biogeographical scale levels in research and conservation. Biological Conservation 117, 285–297 (2004).
Phillips, M. L. et al. Analysis of predator movement in prairie landscapes with contrasting grassland composition. Journal of Mammalogy 85, 187–195 (2004).
Simberloff, D. S. & Abele, L. G. Island biogeography theory and conservation practice. Science 191, 285–286 (1976).
Turner, M. G. Landscape ecology: what is the state of the science? Annual Review of Ecology, Evolution and Systematics 36, 319–344 (2005).
With, K. A. The landscape ecology of invasive spread. Conservation Biology 16, 1192–1203 (2002).
Wu, J. Landscape ecology, cross-disciplinarity, and sustainable science. Landscape Ecology 21, 1–4 (2006).
Wu, J. & Loucks, O. L. From balance of nature to hierarchical patch dynamics: a paradigm shift in ecology. Quarterly Review of Biology 70, 439–466 (1995).










