


















« Prev Next »

In natural and agricultural ecosystems, omnivorous insects are omnipresent. Whether they are 'true omnivores' that require both plant and prey resources to complete their life-cycle, 'herbivores' that occasionally or incidentally consume prey, or 'predators' that drink plant-provided nectar, insect omnivores are a key part of any ecosystem. More than 40 families of insects from over 12 orders are described as omnivorous (Coll & Guershon 2002). Many insects are life-history omnivores, which means that they act as herbivores during one stage of their life, then as predators during another. For example, most parasitoid wasps feed upon their host (another arthropod) during larval development, and then nectar-feed as adults. The more omnipresent form of omnivory in insects, which will be the focus of this article, is when insects consume plant and prey resources during the same developmental stage. These insects consume prey to varying extents, and utilize plant resources in diverse ways, some directly feeding on plant tissues (e.g. true bugs) and others just on nectar (e.g. wasps and ants). Despite their prevalence and ecological importance, ecologists have a hard time incorporating insect omnivores into trophic webs or other ecological models (Polis & Strong 1995). Consuming both plant and prey resources, omnivores function both as 'herbivores' and as 'predators', so they can, respectively, decrease or increase the strength of trophic cascades (Figure 1). The dynamic trophic role that omnivorous insects occupy raises important questions: what environmental factors cause an omnivore to decide to consume plant versus prey resources? How does this trophic role affect the community of organisms in which the omnivore lives? How can humans best utilize omnivorous insects in the biological control of pests?
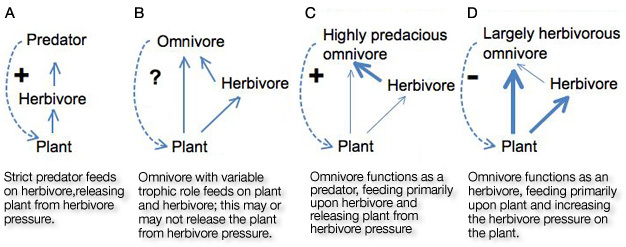
Figure 1
Trophic relationship between plant, herbivore and predator, with and without a plant-feeding, omnivorous predator. Solid lines depict energy transfer, and dotted lines depict ecological relationships (the effect of one organism on the success of another). Figure 1A summarizes a conventional trophic cascade whereby herbivores are killed by predators, resulting in increased plant abundance because of a release from herbivory. In Figure 1B, increasing the abundance of omnivorous predators does not necessarily lead to an increase in plant abundance, since the omnivorous predators plant-feed in addition to consuming herbivores. In Figure 1C, a highly predacious omnivore functions ecologically as a predator, increasing plant fitness by controlling its herbivore (a trophic cascade). In Figure 1D a largely herbivorous omnivore functions as an herbivore, further limiting plant fitness (no trophic cascade).
© 2011 Nature Education All rights reserved. 
Ecology and Evolution of Insect Omnivory
Omnivory as an adaptation by herbivores to obtain sufficient nutrients.
Plant tissue tends to be very protein-poor, which places selective pressure on herbivorous insects to obtain sufficient protein. Insects have managed to cope with this challenge in various ways. Many herbivorous insects, for example, house endosymbiotic microbes in their gut that convert carbohydrates into protein-rich nutrients (Douglas 1998). Many phloem-feeding bugs (e.g., aphids), on the other hand, filter out protein as they consume massive amounts of protein-poor plant sap; the carbon-rich, sugary honeydew that these insects produce is the protein-filtered sap. Herbivorous bugs commonly consume protein-rich pollen grains and seeds from their host plants to complement the protein-poor plant tissues they feed on most of the time. Pollen and seed consumption are considered evolutionary precursors to prey-hunting, being both nutritionally similar and requiring similar physiological adaptations to consume and digest them (Eubanks et al. 2003).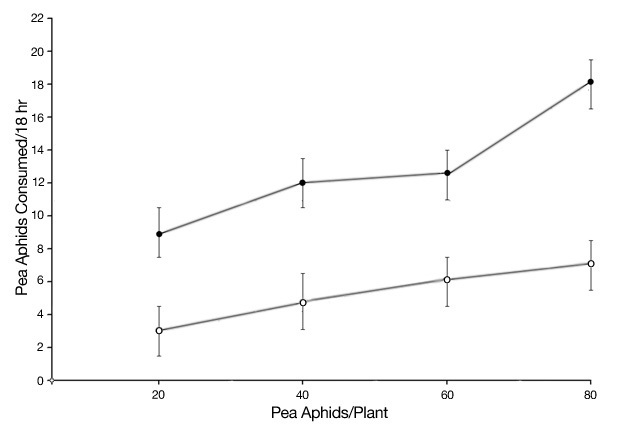
An alternative to these adaptations, utilized by omnivorous insects, is to supplement plant-derived food resources with protein-rich prey. However, in order to obtain prey items an insect must be able to locate, capture, consume, and digest the prey. This often requires adaptations such as acute vision, special mouthpart morphology, venom, and proteinases for digestion. Since capturing a prey item is generally more difficult than feeding from a suitable host plant, prey-hunting by omnivores is more common when protein access from their host plant is insufficient. As such, host plant quality determines the extent to which omnivores hunt prey, which defines their trophic role as herbivore or predator. An experiment conducted by Eubanks and Denno (2000) confirms this prediction. The big-eyed bug Geocoris punctipes is a generalist omnivore that feeds on a range of herbivores, including aphids. Eubanks and Denno manipulated the availability of protein-rich pea pods to Geocoris, and then measured the amount of aphids consumed. As predicted, bugs that consumed the protein-rich pea pods hunted for less prey compared with bugs without access to the pea pods (Figure 2).
Figure 2
Access to high-nutrient plant resources (pea pods) causes omnivorous big-eyed bugs to hunt for less prey (pea aphids). Closed circles represent prey hunting by big-eyed bugs deprived of nutrient-rich pea pods; open circles represent prey-hunting by big-eyed bugs with access to nutrient-rich pea pods. From Eubanks & Denno (2000).
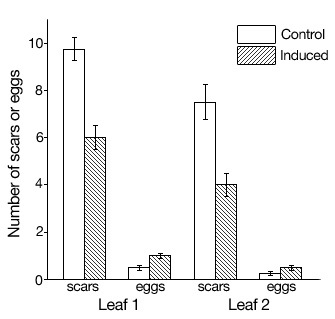
Eubanks and Denno demonstrated that host plant quality determines the trophic role taken by omnivorous insects. From the perspective of the plant, the degree to which a resident omnivore feeds on the plant itself versus the herbivores that consume the plant has important fitness consequences. Agrawal et al. (1999) demonstrated that plants can manipulate the degree to which their resident omnivores consume prey by inducing defensive responses that lower the quality of the plant resource available to the omnivores. To demonstrate this, Agrawal et al. experimentally inflicted herbivore damage on young cotton plants. After removing the herbivores, damaged plants were allowed to grow for a period of time before herbivores and omnivores were placed on them. Cotton responds to herbivore damage by inducing chemical changes that reduces its palatability (e.g., Karban 1997). They then measured the degree to which omnivores fed on plant versus prey resources on induced and un-induced plants. They found that omnivores on induced cotton plants not only fed less on the cotton, but also consumed more of its herbivores (Figure 3). This study, and prior work on the topic have been carried out in agricultural systems, in which the interacting species often lack a long evolutionary history. We currently have a very limited understanding as to what extent plants manipulate the foraging behavior of their resident omnivores in nature, where greater specialization is likely. The exception to this rule is ant-plant mutualisms, which have been widely studied and yielded fascinating insights.
Figure 3
Induced plant defense lowers the quality of plant resources available to omnivorous thrips. This reduces the amount of plant-feeding by the thrips (measured as plant scars), and increases the amount of herbivore eggs they consume. From Agrawal et al. (1999).
Omnivory as part of a mutualistic relationship between plant and predator.
Ant-plant mutualisms demonstrate the importance of omnivory in nature. Plants from a wide range of taxa produce nectar outside of their flowers (extrafloral nectar) to attract ants that consume potentially damaging herbivores. The conventional interpretation is that the ants defend their nectar resource from potential damage, including damage by the plant's herbivores. Ness et al. (2009) demonstrated that the protein content of the extrafloral nectar offered to ants determines the degree to which they hunt for herbivores. Ness et al. manipulated the protein content of extrafloral nectar, then measured how intensely the ants hunted herbivorous caterpillars. Indeed, ants fed protein-poor nectar hunted more intensely and killed more herbivores compared with ants that were supplemented with protein sources. Ness et al. conclude that protein-poor nectar provides fuel for the ants, while simultaneously placing them at a protein deficit. The result was hyped-up ants that are extremely hungry for protein-rich prey.
Importance of Insect Omnivores in Agriculture
2 Why omnivores are effective at suppressing pest populations.
Omnivorous insects are widely used in biological control programs. One of the key challenges in controlling pests with natural enemies is how to attract the natural enemies of the pests before the pests cause significant damage. Omnivores are able to persist in the absence of prey by utilizing plant resources, which allows these insects to maintain high populations before pests arrive. Similarly, once pest populations have been reduced to low densities, the omnivores can persist by plant-feeding (Lalonde et al. 1999). Of course, this can have negative effects on the yield of the plant on which the omnivore is feeding; effective use of omnivores in biological control requires an analysis of both the benefits of the omnivore via pest control, and the costs associated with plant feeding. In many cases, omnivorous insects have relatively tight associations with their host plants compared with insects that are exclusively predators. Omnivorous dicyphine plant bugs, for instance, have adapted to live on hairy plants that many other insects are unable to utilize (Wheeler 2001). These omnivores are important for controlling pests of solanaceous greenhouse crops, such as tomatoes, which are also hairy and difficult for most insects to function on. Other insect predators have failed as a natural pesticide because they cannot forage efficiently on the hairy plant surface.
Use of floral nectar to attract 'predators' and parasitoids.
As mentioned above, many insects that are classically considered predators also consume plant-provided nectar. For example, nectar fuels the foraging efforts of predatory and parasitic wasps, both of which are important agents of biological pest control. Many farmers plant rows of flowering plants among their crops to attract these beneficial insects (Landis et al. 2000). Nectar not only attracts the natural enemies of many agricultural pests to the farm landscape, but also increases their foraging efficiency and longevity once they have arrived.
Conclusion
Omnivory is widespread among insects, both in natural and in agricultural systems. The degree to which these omnivores feed upon plant versus prey resources determines their functional role as functional herbivores or predators. Which role they play can drive ecological interactions in the community. Plants may be able to mediate this trophic role by manipulating the amount of protein available to omnivores. Understanding the interactions between plants and omnivores has provided great insight into how some mutualisms evolve in nature. Because of the importance of insect omnivores as biological control agents, understanding insect omnivory is integral to developing efficient pest control programs.
Glossary
Trophic webs: The feeding relationships between organisms in a population.
Ecological models: Mathematical representations of ecological processes. Ecologists use ecological models to understand complex processes. Model validity refers to how accurately the ecological model estimates real processes.
Trophic cascades: When predators in a trophic web suppress the abundance of their prey, the result is a smaller prey population. As a result, the prey of the prey (plants if the prey is an herbivore, or other animals if it is a predator) is released from predation pressure and increases in abundance.
Biological control of insect pests: In agriculture, pests can be controlled by the use of chemical or biological tools. In biological control, natural biological processes are utilized to control pests; predatory insects and insect diseases are two commonly used tools of biological control.
Endosymbiotic microbes: An endosymbiont is an organism that lives inside the body of another organism. Many insects have microbes that live in their guts and aid in food digestion while additionally providing their hosts with valuable by-products, such as protein.
Nutrients: Chemicals that organisms need to grow and survive. Ecologists and entomologists usually use the word nutrient to mean protein, or other nitrogen-rich compounds that are usually hard for insects to come by.
Phloem: In vascular plants the phloem is the living tissue that transports photosynthates (sugary compounds) throughout the plant. Phloem tends to be high in sugar and low in protein.
Proteinases: A compound needed for the digestion of protein compounds. Proteinases are proteases that begin the hydrolytic breakdown of proteins, usually by splitting them into polypeptide chains.
Inducible plant responses: Plants respond to external and internal stimuli by changing their structural and chemical composition. For example, plants usually respond to damage from herbivores by increasing the production of chemical defenses and toxins in order to prevent subsequent damage by the herbivore.
Ant-plant mutualisms: Mutually beneficial interactions between ants and plants. There is a wide range of ant-plant mutualisms over a spectrum of specialization, from loose, temporary associations to life-long obligate relationships.
Extrafloral nectar: Sugary compounds produced by plants outside of the inflorescence. These sugar gifts function to attract beneficial insects such as ants and wasps to the plant, usually to increase the predation rate of herbivores also found on the plant. Omnivores, which feed on plant and prey, are able to utilize the nectar as fuel to hunt for prey.
Natural enemies: The natural predators of other organisms (typically predators of insect pests on farms). As such, attracting natural enemies to the farm can be beneficial if it increases predation upon the pests.
Dicyphini: A tribe of plant bugs (family: Miridae) in the subfamily Bryocorinae. Many dicyphine plant bugs are highly predatory omnivores, and tend to be specialized to hairy, or hairy-sticky plants. These insects are important agents of biological control on many greenhouse crops.
Solanaceae: A family of flowering plants used in agriculture and medicine, including tomato, pepper, potato, eggplant, and many more.
References and Recommended Reading
Agrawal, A. A., Kobayashi, C., & Thaler, J. S. Influence of prey availability and induced host plant resistance on omnivory by western flower thrips. Ecology 80, 518-523 (1999).
Coll, M., & Guershon, M. Omnivory in terrestrial arthropods: mixing plant and prey diets. Annual Review of Entomology 47, 267-297 (2002).
Douglas, A. E. Nutritional interactions in insect-microbial symbioses: aphids and their symbiotic bacteria Buchnera. Annual Review of Entomology 43, 17-37 (1998).
Eubanks, M. D., & Denno, R. F. Host plants mediate omnivore-herbivore interactions and influence prey suppression. Ecology 81, 936-947 (2000).
Eubanks, M. D., Styrsky, J. D., & Denno, R. F. The evolution of omnivory in heteropteran insects. Ecology 84, 2549-2556 (2003).
Karban, R., & Baldwin, I. T. Induced Responses to Herbivory. Chicago, IL: University of Chicago Press, 1997.
Lalonde, R. G., McGregor, R. R., & Gillespie, D. R. Plant-feeding by arthropod predators contributes to the stability of predator-prey population dynamics. Oikos 87, 603-608 (1999).
Landis, D. A., Wratten, S. D., & Gurr, G. M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Review of Entomology 45, 175-201 (2000).
Ness, J. H., Morris, W. F. & Bronstein, J. L. For ant-protected plants, the best defense is a hungry offense. Ecology 90, 2823-2831 (2009).
Polis, G. A. & Strong, D. R. Food web complexity and community dynamics. The American Naturalist 147, 813-846 (1995).
Wheeler, A. G. Biology of the Plant Bugs. Ithaca, NY: Cornell University Press, 2001.










