





















« Prev Next »

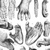
The Old World monkeys of Africa and Asia comprise approximately 132 species in the family Cercopithecidae. They exhibit striking diversity in habitat, distribution, diet, and social behavior. Living in deserts, rainforests, snowy mountains, and even cities, Old World monkeys include two especially species-rich genera and several exceptionally widespread species. They are medium to large monkeys (typically 4–20 kg), with the under 2 kg talapoin and over 30 kg male mandrill and snub-nosed monkey at each extreme. Old World monkeys differ from New World monkeys in having downward-pointing nostrils (Figure 1) and only two pre-molars, while the presence of tails in nearly all forms differentiates them from apes. There are two subfamilies in Cercopithecidae: Cercopithecinae includes monkeys with simple digestive systems and cheek pouches, and Colobinae includes monkeys with guts anatomically specialized for feeding on leaves.

Figure 1
Some Old World monkeys live in and on the outskirts of cities, such as this Japanese macaque living in a remnant forest overlooking downtown Kyoto.
© 2012 Nature Education All rights reserved. 
Colobinae
Recent classifications propose three African and seven Asian genera (Groves 2005). Arboreal colobus are found throughout African forests. In the west, the smallest and least-studied genus, Procolobus (olive colobus), has the most restricted range and is represented by a single species. Piliocolobus (red colobus) includes 16 allopatric species, widely distributed from Senegal to Zanzibar; many are endangered, and one possibly extinct (IUCN 2010). From Guinea to Ethiopia, the five mostly allopatric black-and-white Colobus species have the broadest distribution and occupy the widest variety of forest habitats.
Like their African cousins, most Asian colobines are primarily arboreal, though the forests they inhabit cover a wider range of latitudes and altitudes, from lowland tropical to montane. Grey langurs (Semnopithecus), the most terrestrial colobines, occur throughout western Asia from Sri Lanka to Nepal in habitat ranging from moist, scrub, and conifer forest to gardens and temples. Surilis (Presbytis) and lutongs (Trachypithecus) occupy dipterocarp forest and rubber plantations in Indonesia and Malaysia, with Trachypithecus species extending north to China and west to India. Six allopatric taxa in the Trachypithecus Francoisi group, many of which are endangered, live almost exclusively in limestone karst habitat, sleeping in caves and clambering on rocky cliffs.
All three douc species (Pygathrix) of Vietnam, Cambodia, and Laos are rare and endangered, the grey-shanked douc (P. cinerea) critically so (<700 wild individuals, IUCN 2010). Two other colobine genera, each containing one species, live on Indonesian islands, the endangered proboscis monkey (Nasalis larvatus) in peat swamps and mangrove forests of Borneo, and the critically endangered pig-tailed langur (Simias concolor) in the Mentawai island chain. The four snub-nosed monkey species (Rhinopithecus), also endangered, occupy the most extreme environments — the montane forests of China, Vietnam, and Myanmar, where a new species was recently discovered (Geissman et al. 2010). Yunnan snub-nosed monkeys (R. bieti) spend half the year in snow at elevations up to 4700 meters (Figure 2; Long et al. 1994).

Figure 2
Old World monkeys occupy a broad range of habitats, including — for these Yunnan snub-nosed monkeys (Rhinopithecus bieti) — temperate forests where it snows in winter.
© 2012 Nature Education Photo courtesy of Cyril Grueter. All rights reserved.
Diet
Colobines are leaf-eating specialists with high-cusped molars and multi-chambered stomachs housing mutualistic microbes that digest cellulose and detoxify plant defensive compounds. Leaves are a major component of many species' diets, up to 92% in a population of Ugandan red colobus (Piliocolobus tephrosceles; Snaith and Chapman 2008). The proportion of seeds and fruits in the diet can increase seasonally, however, reaching 87% seeds in the maroon leaf monkey (Presbytis rubicunda; Davies 1991). The winter diet of some mountain-dwelling snub-nosed monkeys is almost exclusively lichen (Kirkpatrick 2010). Colobines are discriminating leaf-eaters, often preferring certain species and younger leaves with fewer structural and chemical defenses. Many African colobines select leaves high in protein and/or low in fiber (Fashing 2010). A growing appreciation of variability and selectivity in colobine diets has led researchers to recognize that these monkeys compete for food more than had been previously thought (Snaith & Chapman 2007).
Ranging and activity
Home ranges of most colobines are small (<1 km2), reflecting the relative abundance of the leaves they eat. Even those with larger ranges typically travel <1 km/day. The enormous home ranges of Nepal gray langurs (Semnopithecus schistaceus) and snub-nosed monkeys (>10 km2) likely reflect their relatively unproductive montane habitat (Kirkpatrick 2010). Range sizes of many colobines vary seasonally, tracking the proportion of different dietary components.
West African olive and red colobus often associate with Diana monkeys (Cercopithecus diana), a cercopithecine. A likely benefit to both colobus and Diana monkeys is mutual predator detection: red colobus, traveling higher in the canopy, are better positioned to detect shared aerial predators, while Diana monkeys sound the alarm (recognized by colobus) for arboreal and terrestrial predators (Fashing 2010). One of the most dangerous species to both is a fellow primate: at some sites over 90% of chimpanzees' mammalian diet is red colobus, who may suffer major long-term decline as a result (Lwanga et al. 2011).
The more leaves consumed, the more time gut microbes need to process them. Colobine field researchers, therefore, excel at watching their stationary subjects digest. One of the largest colobines, the proboscis monkey, spends up to 75% of daylight hours resting (Figure 3; Matsuda et al. 2009).

Figure 3
Proboscis monkeys (Nasalis larvatus), like other colobines, spend a large part of their day resting.
© 2012 Nature Education Photo courtesy of Brian Cretin. All rights reserved.
Social life
Colobine groups are typically moderate-sized (10–20), containing multiple adult females and 1–2 adult males. Multi-male groups of some Asian colobines are best described as traveling assemblages of one-male units. Snub-nosed monkeys are a striking extreme — 20 one-male units can form bands of over 300 individuals (Kirkpatrick 2010). Other multi-male groups may be only temporary, occurring during male take-overs or when a son has matured. The African red colobus is an exception to this pattern, living permanently in larger multi-male groups of 25–50 (Fashing 2010).
Colobines exhibit various patterns of sex-biased dispersal (Fashing 2010, Kirkpatrick 2010). Male-biased dispersal characterizes Asian colobines and African black-and-white colobus, although females may also transfer groups; in some species, dispersing males join all-male groups. In contrast, females are generally the ones to disperse in African red and olive colobus. A notable exception is the endangered Zanzibar red colobus (Piliocolobus kirkii) living at high densities in human gardens, where philopatric females may obtain food more successfully when supported by female kin (Siex 2003).
The prevalence of one-male units in Asian colobines suggests strong male-male competition for resident status. Indeed, interactions between units can be aggressive, and species with the most frequently associating units are the most sexually dimorphic (Grueter and van Schaik 2009). The folivorous diet of colobines rarely leads to aggressive food competition between females, and between-group encounters involve mainly males, who appear to be defending mates. Unlike Asian colobine males, however, male guerezas (Colobus guereza; Figure 4) and ursine colobus (C. vellerosus) appear to defend food resources, perhaps as indirect mate defense. Indeed, female ursine colobus seem to evaluate males based on how well they fight (Teichroeb et al. 2009).

Figure 4
All colobines, here an East African guereza (Colobus guereza), include leaves as a major dietary component.
© 2012 Nature Education Photo courtesy of Yvonne de Jong. All rights reserved.
Infanticide, a dramatic expression of male-male competition, has been reported in several colobines but is apparently absent in some populations and species. Killing another male's offspring can be an adaptive reproductive strategy if the infanticidal male is not the father and if females become sexually receptive sooner than they would have otherwise and then mate with the infanticidal male. Strong support for this sexual-selection explanation comes from gray langurs and ursine colobus (C. vellerosus; Fashing 2010). In Thomas's langur (Presbytis thomasi), infanticide by extra-group males may induce females without infants to transfer with the infanticidal males to smaller groups (Wich and Sterck 2010).
Cercopithecinae
Though the numbers of genera are comparable, cercopithecines are generally more ecologically flexible, widespread, and well-studied than colobines. They include both arboreal and semi-terrestrial forms and inhabit the greatest variety of habitats of any major primate group. Of the two cercopithecine tribes, Papionini, which includes macaques (Macaca), baboons (Papio), geladas (Theropithecus gelada), mandrills and drills (Mandrillus), and mangabeys (Cercocebus, Lophocebus, Rungwecebus), is generally better-studied, especially those taxa that are common and relatively easy to observe, such as savannah baboons living semi-terrestrially in open habitat. In contrast, it can be a challenge to follow mangabeys and the many guenons (Cercopithecus, Allenopithecus, Miopithecus) of the Cercopithecini tribe through their dense forest habitats (Cords in press).
Forest-dwelling species typically have more restricted geographic distributions than those living in open environments, though there are obvious exceptions, such as blue and DeBrazza's monkeys (Cercopithecus mitis, Figure 5; C. neglectus), which are distributed widely in sub-Saharan forests, and geladas, which are limited to high, grassy plains in Ethiopia (Figure 6; Cords in press). The limited distributions of many forest-dwellers make them especially vulnerable to habitat destruction and hunting; two such species — the dryas monkey (C. dryas) of the central Congo basin and the recently discovered Tanzanian kipunji (Rungwecebus kipunji) — are critically endangered.

Figure 5: Strong social bonds among female kin are typical in cercopithecines.
Here, two blue monkey (Cercopithecus mitis) sisters engage in a grooming bout.
© 2012 Nature Education Photo courtesy of Marina Cords. All rights reserved.

Figure 6: Geladas (Theropithecus gelada) occupy high grasslands in Ethiopia, sleeping at night on cliffs.
Photograph shows a one-male unit, the basal unit of their nested society.
© 2012 Nature Education Photo courtesy of Peter Fashing. All rights reserved.
Macaques, the only Asian cercopithecines, are second only to Cercopithecus in number of species and to Homo in geographic range. Their African representatives, Barbary macaques (Macaca sylvanus), are the only non-human primates north of the Sahara, and an introduced European population lives in Gibraltar. Asian macaques span a large area from Japan to India, occupying habitats as varied as lowland rainforests, mountains, temples and cities. As with colobines, most cercopithecines are tropical, but several macaques occur at higher, sometimes snowy elevations, including Barbary, rhesus (M. mulatta), Japanese (M. fuscata), and Taiwanese macaques (M. cyclopsis). Macaque diversity is also evident in the contrast between the success of ecologically adaptable ‘weed' species, such as the widely distributed rhesus and long-tailed (M. fascicularis) macaques, and the critically endangered Celebes crested macaque (M. nigra) of Sulawesi (Figure 1) and Pagai Island macaque (M. pagensis), found in the same disappearing forests as the equally endangered pig-tailed langur.
Diet
Cercopithecine diets are typically more diverse than those of leaf-loving colobines (Cords in press). Baboons and macaques are famous for their dietary flexibility, and in general cercopithecines can be characterized as opportunistic yet selective. As temperate habitats are generally less species-rich than tropical ones, diets in northern latitudes and high elevations are correspondingly less diverse, including primarily leaves and seeds, than those of equatorial and lowland species. Tropical guenons and mangabeys eat the most fruit, whereas geladas eat grass almost exclusively. All cercopithecines have cheek pouches that serve to transport fruits, seeds, and sometimes leaves away from the watchful eyes of predators and competitors alike.
Ranging and activity
Cercopithecines generally travel farther than colobines, as their foods are more scattered. They also spend less time resting and more time socializing, with grooming a major form of social exchange. Many of the forest-dwelling species form mixed-species groups, which may function in predator defense. These associations are especially common among guenons and mangabeys, less so among sympatric macaques and colobines in the same forest.
Social life
Group size and composition are highly variable across and within cercopithecine species (Cords in press). Typical groups include 10–100 individuals, but temporary associations of geladas can surpass 1200 individuals, and mandrill ‘hordes,' which are persistent social units, average 620 members (Swedell 2010).
The groups of many papionins contain multiple males, but some, such as hamadryas baboons (Papio hamadryas), geladas, and, possibly, drills (Mandrillus leucophaeus) and guinea baboons (P. papio), have smaller one-male groups as basal social units. These units are nested into a hierarchical system of bands and troops. Less frequently, single- and multi-male mangabey groups merge into ‘supergroups.' Such sub-grouping may be a way to maintain flexibility in face of changing ecological conditions and to negotiate shared resources such as watering holes and sleeping cliffs. Mandrills show a different type of social flexibility — unique among primates, their massive hordes lack adult males outside of the breeding season (Abernethy et al. 2002).
In contrast to most papionins, almost all guenon groups include just one adult male for most of the year. During breeding seasons, however, resident males may be overwhelmed by an influx of mate-seeking bachelors. Non-resident cercopithecine males are generally solitary; only in geladas, Japanese macaques, and some guenons do there exist all-male groups similar to those of Asian colobines (Cords in press).
Almost all cercopithecine females remain in their natal groups for life; hamadryas baboons are the sole exception. Females form the stable social core, and kin-directed spatial proximity, grooming, and coalitionary support are common (Figure 5; Cords in press). Stable matriline-based hierarchies often develop as a result, but dominance style varies by species. ‘Despotic' species, such as rhesus and Japanese macaques, baboons, and vervets, exhibit strict and steep female dominance relations, with strong kin biases and little reconciliation. In contrast, three Sulawesi macaques (M. nigra, M. tonkeana, M. maurus) exemplify a more symmetrical ‘egalitarian' dominance style (Thierry 2010). Members of the Cercopithecini tribe, however, do not always conform to the traditional cercopithecine pattern: female patas monkeys, though philopatric, only occasionally form stable, matrilineally based hierarchies, and there is only limited evidence in blue monkeys that female rank correlates with any proxy of biological fitness (Foerster et al. 2011). Neither do female blue monkeys form frequent coalitions; they may instead simply minimize contests over food by avoiding each other while feeding.
Relations between male cercopithecines are typically antagonistic. Grooming is rare, though the males of some species form coalitions to improve competitive success. Rather than being kin-biased — males, being the dispersing sex, are not closely related to each other — the decision to participate in coalitions seems to depend instead on reciprocity or mutual advantage in conjunction with differential bargaining positions.
Females typically mate with more than one male, at times seeking extra-group partners, and may engage in sexual activity even when they cannot conceive. Such behavior may confuse paternity, thus forestalling infanticide, which does occur in many cercopithecines. Copulation calls, sexual swellings, and color changes signaling ovulation may incite mate guarding, male-male fighting, and/or sperm competition. Females of many species show preferences for individual males, and not necessarily the highest-ranking ones. In multi-male groups, the degree to which females can exercise mate choice depends on factors such as breeding synchrony, degree of sexual dimorphism, and habitat (i.e., the feasibility of hidden rendezvous). In one-male guenon groups, influxes of extra-group males make it particularly difficult for resident males to monopolize breeding.
Contributions to Primatology
Studies of Old World monkeys have contributed importantly to our understanding of primates in general. Comparisons among species and populations, with their diverse diets and social patterns, have helped especially in understanding the relations between feeding ecology and social evolution.
References and Recommended Reading
Abernethy, K. A., White, L.J.T. et al. Hordes of mandrills (Mandrillus sphinx): extreme group size and seasonal male presence. Journal of Zoology 258, 131-137 (2002).
Cords, M. The behavior, ecology and social evolution of Cercopithecine monkeys. In The Evolution of Primate Societies. ed. Mitani, J., Call, J. et al. eds. Chicago: University of Chicago Press, in press.
Davies, A. G. Seed-eating by red leaf monkeys (Presbytis rubicunda) in dipterocarp forest of northern Borneo. International Journal of Primatology 12, 119-144 (1991).
Fashing, P. J. African colobine monkeys: their behavior, ecology, and conservation. In Primates in Perspective, 2nd edition. ed. Campbell, C. J., Fuentes, A. et al. New York: Oxford University Press, 2010.
Foerster, S., Cords, M. et al. Social behavior, foraging strategies, and fecal glucocorticoids in female blue monkeys (Cercopithecus mitis): potential fitness benefits of high rank in a forest guenon. American Journal of Primatology 73, 870-882 (2011).
Geissmann, T., Lwin, G. et al. A new species of snub-nosed monkey, Genus Rhinopithecus Milne-Edwards, 1872 (Primates, Colobinae), from northern Kachin State, northeastern Myanmar. American Journal of Primatology 73, 96-107 (2011).
Groves C. Order Primates. In Mammal Species of the World. A Taxonomic and Geographic Reference, 3rd edition. ed. Wilson, D. E. & Reeder, D. M. Baltimore: Johns Hopkins University Press, 2005.
Grueter, C. C., van Schaik, C. P. Sexual size dimorphism in Asian colobines revisited. American Journal of Primatology 71: 609-616 (2009).
IUCN. 2010. IUCN Red List of Threatened Species. Version 2010.4. Downloaded on 13 May 2011.
Kirkpatrick, R.C. The Asian colobines: diversity among leaf-eating monkeys. In Primates in Perspective, 2nd edition. ed. Campbell, C. J., Fuentes, A. et al. New York: Oxford University Press, 2010.
Long, Y.C., Kirkpatrick, R.C. et al. Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (Rhinopithecus bieti). Primates 35, 241-250 (1994).
Lwanga, J., Struhsaker, T. et al. Primate population dynamics over 32.9 years at Ngogo, Kibale National Park, Uganda. American Journal of Primatology 73, 997-1011 (2011).
Matsuda, I., Tuuga, A. et al. The feeding ecology and activity budget of proboscis monkeys. American Journal of Primatology 71, 478-492 (2009).
Swedell, L. African papionins: diversity of social organization and ecological flexibility. In Primates in Perspective, 2nd edition. ed. Campbell, C. J., Fuentes, A. et al. New York: Oxford University Press, 2010.
Siex, K. S. Effects of population compression on the demography, ecology, and behavior of the Zanzibar red colobus monkey (Procolobus kirkii). Ph.D. dissertation. Duke University, 2004.
Snaith, T. V., Chapman, C. A. Primate group size and interpreting socioecological models: do folivores really play by different rules? Evolutionary Anthropology 16, 94-106 (2007).
Snaith, T. V., Chapman, C. A. Red colobus monkeys display alternative behavioral responses to the costs of scramble competition. Behavioral Ecology 19, 1289-1296 (2008).
Teichroeb, J. A., Wikberg, E.C. & Sicotte, P. Female dispersal patterns in six groups of ursine colobus (Colobus vellerosus): infanticide avoidance is important. Behaviour 146, 551-582 (2009).
Thierry, B. The macaques: A double-layered social organization. In Primates in Perspective, 2nd edition. ed. Campbell, C. J., Fuentes, A. et al. New York: Oxford University Press, 2010.
Wich, S. A., Sterck, E. H. M. Langurs: Ecology, sexual conflict and social dynamics. In Indonesian Primates. ed. Gursky-Doyen, S. & Supriatna, J. New York: Springer, 2010.










