


















« Prev Next »

Mutualistic interactions, or mutualisms, are ubiquitous in nature. This type of species interaction involves the exchange of goods or services between two species, called mutualist partners. By definition, each species involved in a mutualism must receive a benefit from the interaction, and that benefit usually comes at a cost (Bronstein 1994). However, mutualist partners do not necessarily receive equal benefits or incur equal costs. While the activities of each partner benefits the other species in some way, neither species behaves altruistically. Instead, each species pursues its own selfish interest, and any benefit incurred by the mutualist partner is an unintended consequence of the interaction. Darwin acknowledged that mutualistic interactions are not altruistic when he wrote in On the Origin of Species (1859), " . . . I do not believe that any animal in the world performs an action for the exclusive good of another of a distinct species, yet each species tries to take advantage of the instincts of others . . . "
Many different kinds of organisms are involved in mutualisms, so the types of goods and services that are exchanged are highly variable, as are the mechanisms by which the exchanges are made. For example, the mutualistic interactions between flowering plants and their animal pollinators (Figure 1) are very different from interactions between acacia trees and the ants that inhabit and protect them (Janzen 1966), or interactions between plant and fungal species that form mycorrhizae (van der Heijden and Horton 2009). The variable nature of mutualistic interactions has made it difficult to develop quantitative mathematical models that generalize all mutualisms. However, quantitative models have been developed for particular types of mutualisms (e.g., plant-pollinator interactions) (Mitchell et al. 2009).

Figure 1: Foraging Bombus bee
This Bombus bee is foraging on the flowers of Dicentra cucullaria (L.) Bernh.; pollen is visible on the underside of the bee. (Courtesy of Julie Wittmer)
Pollination Mutualisms
Plant-pollinator interactions are good examples of mutualistic interactions because nearly three-quarters of all extant flowering plants (angiosperms) receive pollination services from animals (National Research Council 2007). Animal pollination is considered the ancestral form of pollination in angiosperms (Hu et al. 2008), and the rapid diversification of flowering plants that occurred 90–130 million years ago has been attributed, in part, to pollination mutualisms (Crepet 2008). The majority of animal pollinators are insects; bees, butterflies, moths, flies, wasps, and beetles are the most common insect pollinators (National Research Council 2007). Many birds and bats are also important pollinator species. Plants always receive pollination services through the interaction, but the goods or services received by the animal pollinators vary. Most pollinators receive food in the form of pollen or nectar, but some bees also use waxes and resins from flowers to build their hives (Michener 2007), and male euglossine bees use volatile compounds from orchid flowers as "perfume" to attract mates (Zimmermann et al. 2006). Other insect species, such as yucca moths, lay their eggs within the yucca flowers they pollinate, and some, but not all, of the seeds produced are consumed by the developing moth larvae (Pellmyr 2003).
Pollination mutualisms have been of interest to biologists for over a century. Darwin addressed their potential importance in the process of natural selection when he wrote, " . . . I can understand how a flower and a bee might slowly become, either simultaneously or one after the other, modified and adapted in the most perfect manner to each other, by the continued preservation of individuals presenting mutual and slightly favourable deviations of structure" (Darwin 1859).
Classical pollination studies were primarily focused on the reproductive cycles of the plant species and, other than their importance as pollen vectors, pollinators were largely ignored. These studies usually took one of two approaches — they were either focused on the evolutionary consequences of pollination dynamics or on the ecological processes that resulted in pollination. Current research in pollination biology attempts to join ecological and evolutionary approaches as a way to understand evolutionary processes in the ecological contexts in which they occur; for example, how pollinator foraging behaviors affect plant fitness by determining plant mating systems (Mitchell et al. 2009).
While mutualisms are often characterized by the kinds of goods or services exchanged, there are several other ways to characterize these interactions: by the degree of dependency, whether the interaction is obligate or facultative; by the degree of specificity, whether the interaction occurs between species pairs or between groups of species; and by the degree of physical association, how physically close the mutualist partners are to one another. These three methods of characterization include intermediates. Regardless of which method is used, it is important to consider the perspective of each mutualist partner when characterizing the interaction.
Pollination mutualisms have been of interest to biologists for over a century. Darwin addressed their potential importance in the process of natural selection when he wrote, " . . . I can understand how a flower and a bee might slowly become, either simultaneously or one after the other, modified and adapted in the most perfect manner to each other, by the continued preservation of individuals presenting mutual and slightly favourable deviations of structure" (Darwin 1859).
Classical pollination studies were primarily focused on the reproductive cycles of the plant species and, other than their importance as pollen vectors, pollinators were largely ignored. These studies usually took one of two approaches — they were either focused on the evolutionary consequences of pollination dynamics or on the ecological processes that resulted in pollination. Current research in pollination biology attempts to join ecological and evolutionary approaches as a way to understand evolutionary processes in the ecological contexts in which they occur; for example, how pollinator foraging behaviors affect plant fitness by determining plant mating systems (Mitchell et al. 2009).
While mutualisms are often characterized by the kinds of goods or services exchanged, there are several other ways to characterize these interactions: by the degree of dependency, whether the interaction is obligate or facultative; by the degree of specificity, whether the interaction occurs between species pairs or between groups of species; and by the degree of physical association, how physically close the mutualist partners are to one another. These three methods of characterization include intermediates. Regardless of which method is used, it is important to consider the perspective of each mutualist partner when characterizing the interaction.
Obligate versus Facultative Mutualisms
A mutualism is obligate when one species relies completely on another species for goods or services. Yucca moths and yucca plants have a reciprocal obligate relationship- the plants cannot make seeds without the yucca moth, and the moth larvae only reach maturity if they eat developing yucca seeds (Pellmyr 2003). However, mutualist partners do not necessarily rely on each other equally. For example, many dioecious plant species rely completely on animal pollinators for reproductive success, so from the plant perspective, the interaction is obligate. However, the pollinator species that visit the flowers may or may not require pollen or nectar from that plant species in order to survive and reproduce, so from the pollinator perspective, the interaction could range from obligate to facultative. Similarly, most bees rely on floral resources for all their needs (nectar, pollen, waxes, etc.), so from the bee perspective, the interaction is obligate. However, most bee species can utilize resources from a number of flowering plants, so they do not necessarily have obligate relationships with individual plant species. In contrast, some hermaphroditic plant species are self-compatible and also capable of self-pollination, so they can make seeds whether or not their flowers are visited by pollinators. The mutualism is facultative from the perspective of these plants, although the quality of seeds produced through self-pollination is often reduced relative to the quality of seeds produced through outcross pollination (Barrett 2003), so these plants still benefit from mutualistic interactions with pollinators.
Specific versus Diffuse Associations
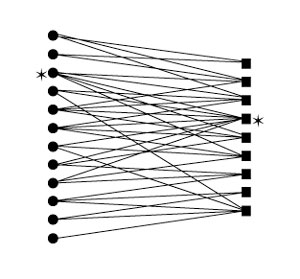
Figure 2: Pollination network
In this diagram of a pollination network, each circle represents an animal species and each square represents a plant species. The lines connecting circles with squares indicate pollination interactions between those species. Notice that some species have more mutualist partners than others (*).
Exhabitational versus Inhabitational Associations
Mutualisms can also be characterized based on the degree of physical association that occurs between the species. Most plant-pollinator interactions are exhabitational because pollinators live separately from the plants they interact with. However, some pollinators live inside the plants they pollinate for a portion of their lives, which is an inhabitational relationship. Fig wasps and fig plants (Cook & Rasplus 2003), and yucca moths and yucca plants (Pellmyr 2003), are good examples of this extremely rare relationship. Characterizations of the most exclusive mutualistic interactions are correlated, in that inhabitational relationships also tend to be highly specific and obligate. However, exhabitational relationships are not necessarily facultative or diffuse.
Mutualisms versus Other Species Interactions
To determine whether or not an interaction is a mutualism, it is necessary to determine the benefit received by each mutualist partner. The details of the interaction are important because one partner could cheat the other, and if so, the interaction is not a mutualism. One common example of “cheating” occurs when an insect removes nectar from flowers without pollinating those flowers (Darwin 1859, Michener 2007). The insect consumes nectar, so this interaction is called nectivory, a specialized form of herbivory, and the insect is often referred to as a "nectar thief." Less frequently, animal pollinators are deceived by plants into providing pollination services. For example, some orchid species deceive their male wasp pollinators by producing flowers that look and smell like female wasps. The male wasps provide pollination services when they mistakenly try to mate with the wasp-like flowers (Gaskett et al. 2008). This interaction is a form of parasitism because the wasps expend energy (and sometimes sperm) while providing pollination services, but do not receive any benefits in return.
Pollination Networks
Pollination mutualisms can affect the outcomes of other kinds of species interactions. Consider a simple example, with two animal-pollinated plant species and one animal pollinator species (Figure 3). If the two plant species flower simultaneously, then they will compete with one another for pollination services. If the pollinator prefers one plant species over the other, then the preferred plant species has a competitive advantage over the second plant species. Both of the plant species have a mutualistic relationship with the pollinator, but the relative fidelity of each mutualism affects the outcome of the competitive interaction between the two plant species. Most communities include many more flowering plants and animal pollinators than are represented in this example, resulting in a network of mutualistic interactions between plants and pollinators. Current research has demonstrated the usefulness of pollination networks as a framework for asking questions about the ecological and evolutionary processes that lead to observed patterns of biodiversity (Bascompte & Jordano 2007, Vasquez et al. 2009).
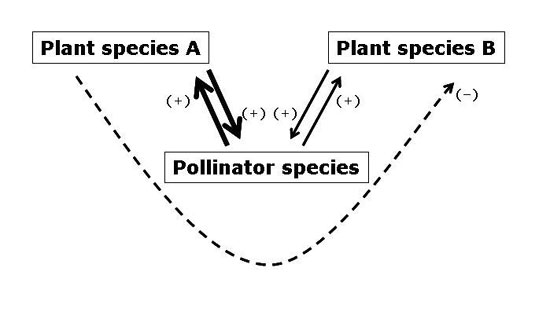
Figure 3: Plant species competing for pollination services
Visual representation of two plants species (Plant species A and B) competing for pollination services from a pollinator species. Each plant has a mutualistic relationship with the pollinator (beneficial interactions are identified with (+) signs), but the pollinator's preference for Plant species A (demonstrated by thicker arrows) causes a negative indirect effect (-) on Plant species B due to competition for pollination services.
Economic Implications
Humans rely on insect pollinators for approximately one third of the food we eat, and a single species provides most of the necessary pollination services — the European honey bee (Apis mellifera) (Klein et al. 2007). These insects are often considered to be an invasive species by ecologists because they have been broadly introduced by humans, and they are able to collect and utilize enormous quantities of floral resources (Buchmann 1996). Honey bees have suffered from a number of maladies over the last few decades, including mite infestations, viral infections, reduced immune function, and recently, colony collapse disorder (Ratnieks & Carreck 2010). While biologists continue to search for solutions to the problems plaguing European honey bee colonies, pollination ecologists are also investigating the potential of wild bees to pollinate agricultural crops (Richards 1996).
References and Recommended Reading
Barrett, S. C. H. Mating strategies in flowering plants: the outcrossing-selfing paradigm and beyond. Philosophical Transactions of the Royal Society of London Series B Biological Sciences 358, 991-1004 (2003).
Bascompte, J. & Jordano, P. Plant-animal mutualistic networks: the architecture of biodiversity. Annual Review of Ecology, Evolution, and Systematics 38, 567-593 (2007).
Bronstein, J. L. Our current understanding of mutualism. The Quarterly Review of Biology 69, 31-51 (1994).
Buchmann, S. L. Competition between honey bees and native bees in the Sonoran Desert and global bee conservation issues. In The Conservation of Bees. eds. Matheson, A., Buchmann, C. et al. (San Diego: Academic Press, 1996). 125-142.
Cook, J. M. & Rasplus, J. Y. Mutualists with attitude: coevolving fig wasps and figs. TRENDS in Ecology and Evolution 18, 241-248 (2003).
Crepet, W. L. The fossil record of angiosperms: requiem or renaissance? Annals of the Missouri Botanical Garden 95, 3-33 (2008).
Darwin, C. On the Origin of species, (1st ed., 1859). Complete and Unabridged Reproduction. New York, NY: CRW Publishing Limited, 2004.
Gaskett, A. C., Winnick, G. C. et al. Orchid sexual deceit provokes ejaculation. The American Naturalist 171, E206-E212 (2008).
Hu, S., Dilcher, D. L. et al. Early steps of angiosperm-pollinator coevolution. The Proceedings of the National Academy of Sciences of the United States of America 105, 240-245 (2008).
Janzen, D. H. Coevolution of mutualism between ants and acacias in Central America. Evolution 20, 249-275 (1966).
Klein, A. M. & Vaissière, B. E. Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society B. Biological Sciences 274, 303-313 (2007).
Landry, C. L., Rathcke, B. J. et al. Flower visitors to white mangrove: a comparison between three Bahamian islands and Florida. In The Proceedings of the 10th Symposium on the Natural History of the Bahamas. eds. Buckner, S. & McGrath, T. (San Salvador Island: Gerace Research Center, 2005) 84-94.
Michener, C. D. The Bees of the World, 2nd ed. Baltimore, MD: The John Hopkins University Press, 2007.
Mitchell, R. J., Irwin, R. E. et al. Ecology and evolution of plant-pollinator interactions. Annals of Botany 103, 1355-1363, 2009.
National Research Council. Status of Pollinators in North America. Washington, DC: National Academies Press, 2007.
Pellmyr, O. Yuccas, yucca moths, and coevolution: a review. Annals of the Missouri Botanical Garden 90, 35-55 (2003).
Ratnieks, F. L. W. & Carreck, N. L. Clarity on Honey Bee Collapse? Science 327, 152-153 (2010).
Richards, K. W. Comparative efficacy of bee species for pollination of legume seed crops. In The Conservation of Bees. eds. Matheson, A, Buchmann, S.L. et al. (San Diego: Academic Press, 1996) 81-103.
Van der Heijden, M. G. A. & Horton, T. R. Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. Journal of Ecology 97, 1139-1150 (2009)
Vasquez, D. P. & Bluthgen, N. Uniting pattern and process in plant-animal mutualistic networks: a review. Annals of Botany 103, 1445-1457 (2009).
Zimmermann, Y., Roubik, D. W. et al. Species-specific attraction to pheromonal analogues in orchid bees. Behavioral Ecology and Sociobiology 60, 833-843 (2006).










