


















« Prev Next »

A population is defined as a group of individuals of the same species living and interbreeding within a given area. Members of a population often rely on the same resources, are subject to similar environmental constraints, and depend on the availability of other members to persist over time. Scientists study a population by examining how individuals in that population interact with each other and how the population as a whole interacts with its environment. As a tool for objectively studying populations, population ecologists rely on a series of statistical measures, known as demographic parameters, to describe that population (Lebreton et al. 1992). The field of science interested in collecting and analyzing these numbers is termed population demographics, also known as demography.
Broadly defined, demography is the study of the characteristics of populations. It provides a mathematical description of how those characteristics change over time. Demographics can include any statistical factors that influence population growth or decline, but several parameters are particularly important: population size, density, age structure, fecundity (birth rates), mortality (death rates), and sex ratio (Dodge 2006). We introduce each of these in turn.
Population Size
The most fundamental demographic parameter is the number of individuals within a population (Lebreton et al. 1992). Population size is defined as the number of individuals present in a subjectively designated geographic range. Despite the simplicity in its concept, locating all individuals during a census (a full count of every individual) is nearly impossible, so ecologists usually estimate population size by counting individuals within a small sample area and extrapolating that sample to the larger population. Regardless of the challenges in measuring population size, it is an important characteristic of a population with significant implications for the dynamics of the population as a whole (Lebreton et al. 1992).
Populations display distinctive behaviors based on their size. Small populations face a greater risk of extinction (Caughley 1994). Individuals in these populations can have a hard time finding quality mates so, fewer individuals mate and those that do risk inbreeding (Hamilton 1967). Additionally, individuals in small population are more susceptible to random deaths. Events like fire, floods, and disease have a greater chance of killing all individuals in the population.
Large populations experience their own problems. As they approach the maximum sustainable population size, known as carrying capacity, large populations show characteristic behavior. Populations nearing their carrying capacity experience greater competition for resources, shifts in predator-prey relationships, and lowered fecundity. If the population grows too large, it may begin to exceed the carrying capacity of the environment and degrade available habitat (Figure 1).

Figure 1: Swarms of locusts exceed carrying capacity with huge population sizes.
These short-lived spikes in population size produce swarms capable of destroying farms as they move across the agricultural landscapes, eating everything in their path.
Photo courtesy of Compton Tucker/NASA GSFC.
Population Density
A more complete description of a population's size includes the population density — the size of a population in relation to the amount of space that it occupies. Density is usually expressed as the number of individuals per unit area or volume (Lebreton et al. 1992). For example: the number of crows per square kilometer or the number of plankton per liter (Andren 1992, Sterner 1986). Like all population properties, density is a dynamic characteristic that changes over time as individuals are added to or removed from the population. Closely related species of Gannet birds will maintain very different densities (Figure 2 ). Birth and immigration — the influx of new individuals from other areas — can increase a population's density, while death and emigration — the movement of individuals out of a population to other areas — can decrease its density (Lebreton et al. 1992).

Figure 2: Gannets can persist at very high densities.
They have developed exaggerated territorial behavior as an adaptation to sustain these densely packed colonies.
Photo courtesy of Follash via Wikimedia Commons.
Similar to population size, population density displays distinctive characteristics at both high and low values. Density-dependent factors, including competition, predation, migration and disease, intensify within populations as density increases. In contrast, density-independent factors, such as weather, fire regimes, and flooding, impact populations regardless of their specific densities (Lebreton et al. 1992).
Age Structure
Not all individuals contribute equally to a population. Occasionally, researchers find it useful to characterize the different contributions made by different individuals. First, individuals are sorted into age-specific categories called cohorts, such as "juveniles" or "subadults" (Dodge 2006). Researchers then create a profile of the size and age structures of the cohorts to determine the reproductive potential of that population, in order to estimate current and future growth. Usually, a rapidly expanding population will have larger reproductive cohorts, stable populations show a more even distribution of age classes, and rapidly declining populations have large older cohorts (Lebreton et al. 1992).
Age structure can be represented graphically with a population pyramid (Figure 3). Although a population's age structure is not always pyramidal in shape, most populations have younger cohorts that are larger than older cohorts. For example, Sherman and Morton's studies of the Tioga Pass Belding's ground squirrels revealed birth cohorts larger than 300 individuals and less than 10 individuals in cohorts over the age of six (Sherman & Morton 1984).
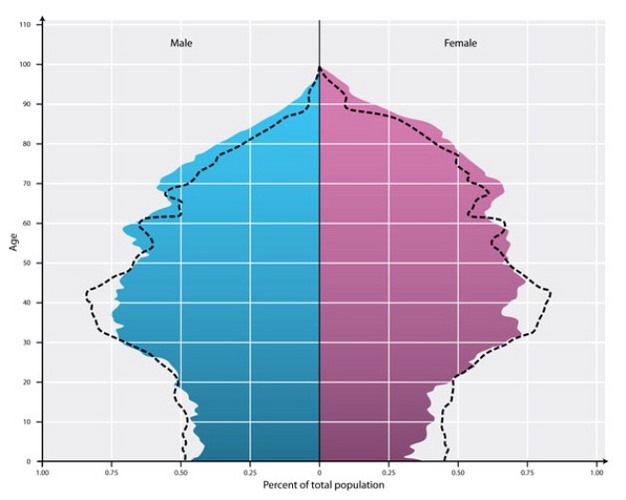
Figure 3: An age-structure pyramid from the Italian township of Matelica displays the dominant middle-age age class.
An age-structure pyramid from the Italian township of Matelica displays the dominant middle-age age class.
Photo courtesy of Qc72 via Wikimedia Commons.
Fecundity
As age structure suggests, some individuals within a population have a greater impact on population-level processes, such as growth. Fecundity describes the number of offspring an individual or a population is able to produce during a given period of time (Martin 1995) (Figure 4). In demographic studies, fecundity is calculated in age-specific birth rates, which may be expressed as the number of births per unit of time, the number of births per female per unit of time, or the number of births per 1,000 individuals per unit of time. Maximum (or physiological) fecundity is the theoretical maximum number of offspring produced in a population assuming no ecological constraints. However, since every ecosystem implements constraints on its populations, ecologists prefer to measure realized (or ecological) fecundity, which is the observed number of offspring produced in a population under actual environmental conditions.
Figure 4: A communal nest of skink eggs displays the fecundity of several individuals.
© 2012 Nature Education Photo courtesy of Kika Tarsi. All rights reserved. 
While maximum fecundity is a constant for populations, realized fecundity varies over time based on the size, density, and age structure of the population. External conditions, such as food and habitat availability, can also influence fecundity. Density-dependent regulation provides a negative feedback if the population grows too large, by reducing birth rates and halting population growth through a host of mechanisms (Lebreton et al. 1992). In white-footed mice, for example, populations regulate their reproductive rate via a stress hormone. As population densities increase, so do aggressive interactions between individuals (even when food and shelter are unlimited). High population densities lead to frequent aggressive encounters, triggering a stress syndrome in which hormonal changes delay sexual maturation, cause reproductive organs to shrink, and depress the immune system (Krohne 1984).
Mortality
Another individual trait that affects population growth is mortality. Mortality is the measure of individual deaths in a population and serves as the counterbalance to fecundity. Like fecundity, mortality is measured in rates, usually expressed as the number of individuals that die in a given period (deaths per unit time) or the proportion of the population that dies in a given period (percent deaths per unit time). A population theoretically has a minimum mortality — the theoretically minimum number of deaths in a population assuming no ecological constraints. The minimum mortality is always lower than the realized (or ecological) mortality, which is the observed number of losses in a population under actual environmental conditions. Much like realized fecundity, realized mortality varies over time based on dynamics internal and external to the population (Benrey & Denno 1997).
To visualize mortality and fecundity within a population, ecologists create life tables to display age-specific statistical summaries of a population's survival patterns. First developed by Roman actuaries, life tables were used to estimate how long individuals of a particular age category were expected to live in order to value life insurance products (Trenerry 1926). Raymond Pearl (1928) first introduced the life table to biology when he applied it to laboratory studies of the fruit fly, Drosophila. Life tables are particularly useful for species with discrete developmental stages and mortality rates that vary widely from one stage to the next (Figure 5).

Figure 5: The Green Stink Bug (Nezara viridula) assumes different body forms through metamorphosis between different discrete life stages.
Photo courtesy of Jovo26 via Wikimedia Commons
Sex Ratio
Sexually reproducing organisms must find mates in order to produce offspring. Without comparable numbers of males and females, mating opportunities may be limited and population growth stunted. Thus, ecologists measure the number of males and females within a population to construct a sex ratio, which can help researchers predict population growth or decline. Much like population size, sex ratio is a simple concept with major implications for population dynamics. For example, stable populations may maintain a 1:1 sex ratio and therefore keep their growth rate constant, whereas declining populations may develop a 3:1 sex ratio favoring females, resulting in an increased growth rate. In species where males contribute significantly to offspring rearing, populations may instead maintain a ratio skewed towards males (Hamilton 1967).
Interestingly, sex ratio is not always random but can be manipulated at birth by environmental or physiological mechanisms. All crocodiles and many reptiles utilize a strategy called environmental sex determination, wherein incubation temperature determines the sex of each individual (Delmas et al. 2008). For example, low temperatures will produce males and high temperatures will produce females. In times of limited resources or high population densities, females can manipulate the sex ratios of their clutch by spending more or less time incubating their eggs (Girondot et al. 2004).
Summary
A population is a group of individuals of the same species interacting within the same space. The health and behavior of a population is determined by the way those individuals interact with each other and with their surrounding environment. We can determine the status of a population by measuring and calculating elements common to all populations, such as size, density, fecundity, mortality, sex ratio, and age structure. These demographic statistics provide researchers with standardized metrics for comparing populations and assessing extinction risk. Just as a doctor can't diagnose an illness with only a blood pressure measurement, we can't manage populations based on demographic parameters alone. However, demographics provide a baseline for monitoring population health and offer insight into the mechanisms driving population declines.
Glossary
age-specific: The age of the individual is important for statistical purposes.
clutch size: The number of offspring one female produces in one reproductive cycle.
cohort: Group of all individuals sharing a statistical factor (such as age or developmental stage)
density-dependent factors: Depending on the local density of the population
density-independent factors: Not linked to the local density of the population
discrete developmental stages: Non-overlapping and structurally distinct growth stages. E.g. tadpoles are one discrete developmental stage and adult frogs are another.
ecosystem: A natural system including the interaction of all living and non-living elements.
extinction: No longer existing.
extrapolating: Estimating an unknown value by assuming that a known value can translate (without distortion) to the scale of the unknown value.
growth rate: The rate of change of population size over time.
inbreeding: Breeding of closely related individuals, often with negative genetic consequences.
incubated: Provided with a heat source during embryonic development.
life tables: Specific format of statistical summary of demographic parameters.
migration: Populations moving from one geographic location to another.
objectively: To study without bias and by measurable and repeatable metrics.
offspring: The individual produced from the reproduction of its parents.
parameter: A value in an equation that does not vary. These values can change between different equations of similar form.
predator-prey relationships: How populations of predators are interacting with populations of prey.
predation: The act of killing another living organism for food.
physiological: The parts and functions of living organisms.
reproductive organs: Specialized collection of cells used to exchange gametes between sexually reproducing organisms.
rates: A mathematical term for the number of things or events happening in a given amount of time.
rearing: To invest energy in the growth and development of offspring after they are born.
subjectively designated geographic range: A parcel of land, the size of which is chosen without using standardized criteria. Picked at the discretion of the researcher.
sustainable: System able to be maintained itself indefinitely without supplement.
sexual maturation: An individual reaching a stage of development where it is able to sexually reproduce.
stress hormone: Chemical compounds synthesized in the body to chemically communicate a stress reaction to various systems within that organism.
statistic: A number acting as a description for more numbers.
theoretical: A rational, rather than practical, description of natural phenomena. Often using mathematical, conceptual, or mechanical models to study the underlying functional framework of nature.
References and Recommended Reading
Andren, H. Corvid density and nest predation in relation to forest fragmentation: A landscape perspective. Ecology 73, 794-804 (1992).
Bull, J. Evolution of environmental sex determination from genotypic sex determination. Heredity 47, 173-184 (1981).
Caughley, G. Directions in conservation biology. Journal of Animal Ecology 63, 215-244 (1994).
Delmas, V., Pieau, C. & Girondot, M. A mechanistic model of temperature-dependent sex determination in a chelonian, the European pond turtle. Functional Ecology 22, 84-93 (2008).
Dodge, Y. The Oxford Dictionary of Statistical Terms. Oxford, UK: Oxford University Press, 2006.
Benrey, B & Denno, R. F. The slow-growth-high-mortality hypothesis: A test using the cabbage butterfly. Ecology 78, 987-999 (1997).
Girondot, M. et al. "Implications of temperature-dependent sex determination for population dynamics," Temperature-Dependent Sex Determination in Vertebrates, 148-155, eds. N. Valenzuela & V. Lance. Smithsonian Books, 2004.
Hamilton, W. D. Extraordinary sex ratios. Science 156, 477-488 (1967).
Harcombe, P. A. Tree life tables. BioScience 37, 557-568 (1987).
Hutchinson, G. E. Population studies: Animal ecology and demography. Bulletin of Mathematical Biology 53, 193-213 (1991).
Jiguet, F. et al. Bird population trends are linearly affected by climate change along species thermal ranges. Proceedings of the Royal Society B: Biological Sciences 277, 3601-3608 (2010).
Krohne, D. T., Dubbs, B. A. & Baccus, R. An analysis of dispersal in an unmanipulated population of Peromyscus leucopus. American Midland Naturalist 112, 146-156 (1984).
Lebreton, J-D. et al. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecological Monographs 62, 67-118 (1992).
Martin, T. E. Avian life history evolution in relation to nest site, nest predation, and food. Ecological Monographs 65, 101-127 (1995).
Pearl, R. The Rate of Living, Being an Account of Some Experimental Studies on the Biology of Life Duration. New York, NY: Alfred A. Knopf, 1928.
Stearns, S. C. Life history tactics: A review of the ideas. The Quarterly Review of Biology 51, 3-47 (1976).
Sterner, R. Herbivores' direct and indirect effects on algal populations. Science 231, 605-607 (1986).
Trenerry, C. F. The Origin and Early History of Insurance, Including the Contract of Bottomry. London, UK: P. S. King & Son, 1926.










