


















« Prev Next »

Top-down forces (i.e., predation) have long been recognized as important community structuring mechanisms. Predators in marine, freshwater, and terrestrial environments may affect prey populations and communities by consuming lower trophic levels (lethal or consumptive effect) or by altering prey traits including behavior, morphology, or habitat use (nonlethal or nonconsumptive effect). Lethal and nonlethal interactions between predators and prey alter patterns of energy flow, community diversity and composition, and the importance of competitive interactions.
Despite the obvious importance of predators in communities, it is widely accepted that the effects of predators are minimized when environmental conditions interfere with their foraging activities. A primary mechanism is the restriction of foraging time available to mobile predators due to the risk of injury accompanied by reduced locomotory performance or efficiency in adverse conditions. Consequently, models of community regulation note the importance of predation on community functioning in benign environments, an effect which declines in stressful conditions. For example, hydrodynamic stress associated with waves reduces the effectiveness of predators, as these conditions limit predator mobility and foraging activity, whereas predation becomes an important community-structuring agent on leeward shores protected from wave action.
Environmental conditions may not be stressful in the sense of presenting physical conditions that are life-threatening, but may nevertheless negatively impact predator foraging if they limit the ability of predators to find and/or consume prey. For example, green crab predation was significantly reduced in areas of the Damariscotta River (a tidal estuary in southern Maine) with high flow velocities, even though green crabs were more abundant in high flow sites. Similarly, fish, insects, and copepods experienced diminished foraging success in more rapid flows that were not otherwise harmful. Thus environmental conditions can impact consumers by decreasing their ability to detect or handle their prey, not by physically preventing them from inhabiting a given location, as is the case on wave-swept shores.
Behavioral research suggests that environmental conditions such as hydrodynamics (i.e., flow rates) may have particularly large impacts in communities where predators hunt prey using chemical cues. Hydrodynamic forces such as flow velocity and turbulence can have significant effects on the structure of waterborne chemical odor plumes via mixing and dilution. These processes strongly affect how organisms perceive chemical signals from potential predators and prey. For example, blue crabs (Callinectes sapidus) often locate prey by following chemical odor plumes, but their chemoreceptive abilities and foraging success decline as flow velocities and turbulence increase. These observations, combined with field studies examining green crab predation rates in varying flows in the Damariscotta River, initially led ecologists to the implicit assumption that predator foraging success would decline as environmental conditions (e.g., swift and/or turbulent flow) limit predators’ ability to locate and consume prey.
However, recent studies have shown that a given environmental factor, such as flow, may also impact prey behavior. For example, behavioral studies with hard clams (e.g., Mercenaria mercenaria), indicate they detect chemical cues emanating from predators and injured conspecifics, and respond by reducing their feeding (pumping) time. Clam predators, including blue crabs, locate clams by detecting chemical odors produced by clams as they pump water. By reducing their pumping time, clams minimize the amount of chemicals they release into the environment, and consequently become harder for predators to detect, which increases their survival rate. Like crabs, clams are less responsive to predators in faster flows, and can become more vulnerable to these predators when turbulence increases. Another wrinkle in hard clam-predator interactions is that knobbed whelks (Busycon carica) are actually more efficient predators in faster, more turbulent flows, and predation of clams by whelks increases with increased turbulence. Thus, flow affects predators, prey, and the outcomes of these interactions, but the effects are specific to the species involved.
Numerous studies have shown that predators can affect prey populations and community composition via both lethal and nonlethal mechanisms. However, predicting and modeling the magnitude and prevalence of lethal v. nonlethal effects, and partitioning out the relative contribution of each to the overall degree of top-down control in communities, remains an important goal of current ecological studies. The goal of my research is to examine how environmental conditions, like hydrodynamic flow, affect top-down control, and the relative magnitude of lethal v. nonlethal effects in natural communities, by affecting the sensory capabilities of predators and prey.
In simplest terms, whether a predator successfully attacks and consumes a prey organism, or whether the prey is able to avoid being eaten, often depends upon advantages in perceptive ability. That is, the organism (predator or prey) better able to sense the other is more likely to survive in a given predator-prey interaction. Since nonlethal predator effects occur when prey detect and respond to predators, conditions that inhibit prey sensory abilities should minimize nonlethal predator effects. These types of conditions may simultaneously increase lethal effects if prey are less able to detect predator threats and are more vulnerable to consumers. Conversely, conditions that are favorable for prey sensory capabilities should increase the occurrences of predator detection and nonlethal predator effects, but decrease the lethal effects of predators.
Using hard clams (Mercenaria mercenaria) and blue crabs (Callinectes sapidus) as a model system, I have been investigating the effects of flow on the outcomes of clam-crab interactions. These organisms detect one another using chemical cues, and the sensory abilities of both are reduced in faster, more turbulent flows. In a recently published field study, I and two colleagues found that crab predation on clams was highest at intermediate flow velocities and turbulence levels (Figure 1). In slower, less turbulent flows, clams are better able to detect and avoid crab predators, but clam ability to do so declines when flow velocity and turbulence increase. Although, crabs are also less able to detect clams in more turbulent flows, they are less affected by these conditions than their clam prey, and consequently are more effective predators. However, when flow velocity and turbulence increase to a point where crabs are no longer able to detect clams, predation again declines. In this system, nonlethal predator effects predominate in slow flows, lethal predator effects predominate in intermediate flows, and both diminish in very fast flows when conditions impair the sensory abilities of both organisms.
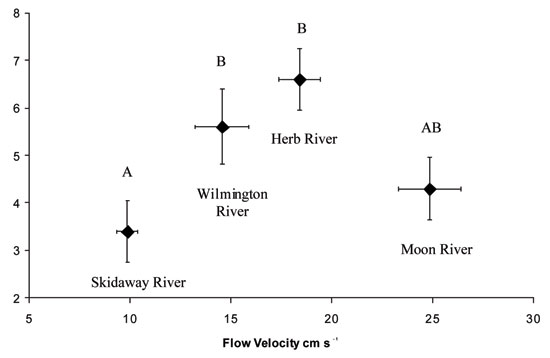
Figure 1: Effects of flow on hard clam predation by blue crabs
Mean number of clams consumed is plotted against mean flow velocity, error bars are +/- SE. Letters denote significant differences in mean predation levels. Letters AB indicate that the intermediate value is not statistically different than A or B. Predation is highest at intermediate flow velocities. (This figure was previously published by Smee et al. 2010 in Ecology.)
More recently I have been examining how flow rates affect predatory interactions in rocky intertidal systems. In New England, the green crab (Carcinus maenas) consumes the dog whelk (Nucella lapillus), and dog whelks in turn consume barnacles (Semibalanus balanoides). Several authors have noted that green crabs cause a trophic cascade which increases barnacle abundance by consuming dog whelks, and by causing dog whelks to seek refuge and not consume barnacles. Thus, both lethal and nonlethal predator effects are active in this system. Using laboratory behavioral assays, members of my lab have investigated how flow velocity and turbulence influence green crab ability to forage on dog whelks and dog whelk responses to green crabs. Our results differ somewhat from clam-blue crab interactions in that slower flows seem to benefit green crabs and intermediate flows are best for dog whelks, although ongoing field studies will be needed to validate these observations. Despite differences, results from hard clam-blue crab and dog whelk-green crab interactions suggest that environmental conditions like hydrodynamic flow can have large effects on the outcomes of predatory interactions, as well as the occurrence of lethal and nonlethal predator effects. Future studies will continue to investigate how environmental conditions influence lethal and nonlethal predator effects by influencing the sensory abilities of predators and prey.
References and Recommended Reading
Beyer, H., Merrill, E. et al. Willow on Yellowstone's northern range: evidence for a trophic cascade? Ecological Applications 17, 1563-1571 (2007).
Carpenter, S. R., Kitchell, J. F. et al. Cascading trophic interactions and lake productivity. Bioscience 35, 634-639 (1985).
Estes, J., Tinker, M. et al. Killer whale predation on sea otters linking oceanic and nearshore ecosystems. Science 282, 473 (1998).
Estes, J. A. & Palmisano, J. F. Sea otters: Their role in structuring nearshore communities. Science 185, 1058-1060 (1974).
Grabowski, J. H. Habitat complexity disrupts predator-prey interactions but not the trophic cascade on oyster reefs. Ecology 85, 995-1004 (2004).
Gruner, D. S. Attentation of top-down and bottom-up forces in a complex terrestrial community. Ecology 85, 3010-3022 (2004).
Jackson, J., Kirby, M. et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 293, 629 (2001).
Kats, L. B. & Dill, L. M. The scent of death: Chemosensory assessment of predation risk by prey animals. Ecoscience 5, 361-394 (1998).
Morrison, L. W., & Porter, S. D. Testing for population-level impacts of introduced Pseudacteon tricuspis flies, phorid parasitoids of Solenopsis invicta fire ants. Biological Control 33, 9-19 (2005).
Myers, R., Baum, J. et al. Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science 315, 1846 (2007).
Paine, R. T. Food web complexity and species diversity. American Naturalist 100, 65-75 (1966).
Preisser, E. L., Bolnick, D. I. et al. Scared to death? The effects of intimidation and consumption in predator-prey interactions Ecology 86, 501-509 (2005).
Preisser, E. L., Bolnick, D. I. et al. Resource dynamics influence the strength of non-consumptive predator effects on prey. Ecology Letters 12, 315-323 (2009).
Regan, H. M., Lupia, R. et al. The Currency and Tempo of Extinction. American Naturalist 157, 1-10 (2001)
Schmitz, O., Beckerman, A. et al. Behaviorally mediated trophic cascades: effects of predation risk on food web interactions. Ecology 78, 1388-1399 (1997).
Schmitz, O. J. Direct and indirect effects of predation and predation risk in old-field interaction webs. American Naturalist 151, 327-342 (1998).
Schmitz, O. J., Grabowski, J. H. et al. From individuals to ecosystem function: Toward an integration of evolutionary and ecosystem ecology. Ecology 89, 2436-2445 (2008).
Sih, A., Crowley, P. et al. Predation, competition, and prey communities: a review of field experiments. Annual Review of Ecology and Systematics 16, 269-311 (1985).
Stachowicz, J., Bruno, J. et al. Understanding the effects of marine biodiversity on communities and ecosystems. Annual Review of Ecology, Evolution and Systematics 38, 739-766 (2007).
Trussell, G. C., Ewanchuk, P. J. et al. Field evidence of trait-mediated indirect interactions in a rocky intertidal food web. Ecology Letters 5, 241-245 (2002).
Trussell, G. C., Ewanchuk, P. J. et al. Trait-mediated effects in rocky intertidal food chains: predator risk cues alter prey feeding rates. Ecology 84, 629-640 (2003).
Werner, E. E. & Peacor, S. D. A review of trait-mediated indirect interactions in ecological communities. Ecology 84, 1083-1100 (2003).










