


















« Prev Next »

Humans depend to a large degree on goods provided by natural and managed ecosystems. These goods and other benefits provided by ecosystems to mankind are collectively referred to as ecosystem services. Anthropogenic activities impact the diversity of organisms found in ecosystems aboveground and belowground, and thus influence the provision of ecosystem services. Consequently, there has been increasing scientific interest in the link between biodiversity and the provision of ecosystem services but the research has focused mainly on aboveground systems. Here we give a brief introduction to the importance of ecosystem services provided by soils to the well being of humans, and then show how soil biota contribute to the provision of ecosystem services. We will then explore the relationship between soil biodiversity and ecosystem services, and discuss why biodiversity theoretically might influence the rate and stability of ecosystem service provision.
Ecosystem Services
The term ‘ecosystem services' was defined in the Millennium Ecosystem Assessment (MEA 2005) as ‘the benefits people obtain from ecosystems', both natural and managed. These services may be categorized as provisional, regulative, cultural or supporting services, also referred to as supporting processes (Table 1). The first three categories have a direct impact on human well-being, whereas the latter has an indirect impact by supporting provisional, regulative, and cultural services. For simplicity we will refer to all categories as services throughout this article. However, all these services, whether direct or indirect, are essential for human life and the well-being of humans (Costanza et al. 1997, Daily et al. 1997, Wall 2004, MEA 2003, 2005).
|
Service type |
Examples of goods or services provided |
| Provisional | Fiber; Food; Freshwater; Fuel wood and other essential resources |
| Regulative | Climate regulation; Disease control and suppression of pathogens; Water purification and regulation |
|
Supporting (processes) |
Nutrient cycling; Primary production; Soil formation |
| Cultural | Aesthetics; Cultural heritage and sense of place; Educational; Recreational; Spiritual and religious |
| Table 1: Classification of ecosystem services. Based on Millennium Ecosystem Assessment (MEA, 2003). | |
Quantifying the value of ecosystem services is of great importance for developing arguments and programs for protecting them. The value of some ecosystem services, such as the provision of food, fuel and fiber, can be quantified, but estimates often reflect single services rather than incorporating all services provided by an ecosystem (but see Nelson et al. 2009, and Barbier 2011). Moreover, for other services, such as mitigation of drought and floods, climate regulation, soil erosion prevention and water filtration, and services provided in the form of recreational, aesthetic or cultural values, it is very difficult to assign a price tag particularly because values vary across national and local boundaries. Nonetheless, preliminary estimates of the value of ecosystem services provided by Earth's biodiversity are on the order of trillions of US dollars annually (Daily et al. 1997, Costanza et al. 1997). This value is likely grossly underestimated, but points toward the great impact such services have on our lives and how a continued provision of these services is necessary to maintain the human population.
Currently there is a rapid increase in human population size. Both because of this growth and changing patterns of consumption, humans are altering ecosystems on a global scale. In addition to the development of land for roads and buildings, there is an increasing demand for food associated with the increasing human population, which has had a big influence on the landscape. For instance, cropland and pastures now occupy approximately 40% of the Earth's land surface area (Foley et al. 2005). Such anthropogenic impacts have contributed to an unprecedented increase in the rate of species extinction globally, which is currently estimated to be 100 to 1000 times higher than average background rates (May 2010). How will losses of species affect the provision of ecosystem services?
This pattern of anthropogenic impacts on natural as well as managed ecosystems, and the large role living organisms play in ecosystem processes such as nutrient cycling, has resulted in increased interest in the connection between biodiversity and ecosystem services. The observed diversity of biota within a community is often referred to as an ecosystem's ‘biodiversity' or biological diversity. Although the term biodiversity may refer to anything from the number and relative abundance of genes within a community to ecosystem types within a landscape (as defined by the 1992 Convention on Biological Diversity (http://www.cbd.int/cop10/), we will use the term as a reference to the number of species within a community.
Over the last few decades, numerous studies and experiments have investigated whether more species in a community would help maintain the provision of ecosystem services, or specifically if there is a positive relationship between biodiversity and ecosystem functioning (functioning or process rate is often considered to be equivalent to the term ‘services' but the actual relationship is still being discussed and quantified). Most studies have focused on aboveground terrestrial and aquatic ecosystems and have shown, in general, a consensus for the relationship between biodiversity and ecosystem function, stability and resource use efficiency. Although it is still being resolved, evidence shows that increased species richness on average leads to greater functioning: for example, productivity in plant communities increases nutrient retention in ecosystems, and provides greater stability in terrestrial and aquatic ecosystems (Tilman 2000, but see also Balvanera et al. 2006, Cardinale et al. 2006, Ives & Carpenter 2007).
Despite the bias towards studies in aboveground and aquatic ecosystems there is, as we show in the following sections, evidence for a similar positive relationship between soil biodiversity and ecosystem services. First, however, we introduce the soil biota and show how this biota contributes to the provision of ecosystem services, and then we provide an overview of evidence for a relationship between soil biodiversity and ecosystem services. Finally we discuss some key mechanisms that may theoretically contribute to the observed biodiversity-ecosystem relationships in soils as well as elsewhere. However, some soil services, such as carbon sequestration, are optimal in systems with inherently low species richness. Carbon is best sequestered in ecosystems where organic matter is allowed to accumulate due to impaired decomposition (i.e., where decomposition rates are lower than net primary production, such as in peatlands where water logging and low pH impair decomposition and species richness). Hence, whether the relationship between biodiversity and ecosystem services in any ecosystem is negative, neutral or positive will depend upon the service in question (i.e., what service are we measuring from a system?), whereas the relationship between biodiversity and process rates (henceforth 'function') are more likely to be positive. For simplicity we interpret greater ecosystem functioning as evidence of a positive response to increases in biodiversity.
Soil Biota and their Contribution to Human Well-Being
Soil is a biologically, physically, and chemically diverse entity that forms the basic substrate of terrestrial ecosystems, supports many human activities and provides a multitude of highly valuable ecosystem services (Dominati et al. 2010). Soil was defined by Bardgett (2005) as the ‘interface between the atmosphere and lithosphere, the outermost shell of the Earth', and is formed over time through the interactions between weathering of the parent material, local climate and topography, and the organisms that live therein. All these organisms are collectively referred to as the soil biota, and they form an integral part of soils. Soil biota range from single celled microorganisms to nematodes that live in the water film on the surface of soil aggregates, and the microarthropods that live in air-filled soil pores (Figure 1), to larger soil animals, such as earthworms and other ecosystem engineers that can manipulate the soil structure and create their own habitat. This biota plays a large role in the regulation of many of the processes occurring in soils and the services that depend on them (Table 2). Current estimates of the contribution of soil biota to ecosystem services provided by soils globally range from 1.5 to 13 trillion US Dollars annually (Van der Putten et al. 2004, and European Soil Bureau Network 2005).

Figure 1: Picture showing microarthropods extracted from a single soil core measuring 3.5 cm diameter and 5 cm depth collected in a birch woodland, Scotland.
Abundances often amount to several 100’s of thousands individuals per m2.
© 2012 Nature Education Courtesy of U.N. Nielsen. All rights reserved. 
| Service type | Goods or services provided |
| Provisional | Provision of clean water through infiltration; Production of food, fodder, fuel, fibre and raw materials; Physical support |
| Regulative | Mitigation of floods and droughts through water retention; Regulation of biogeochemical cycling, global climate patterns, animal and plant populations, and potential pests and pathogens; Erosion control; Bioremediation; Translocation of nutrients, particles, gasses (incl. trace gasses) |
| Supporting (processes) | Nutrient cycling incl. decomposition, retention and delivery of nutrients; Soil biological activity; Soil formation; Water cycling |
| Cultural | Recreation; Spirituality; Aesthetics; Sense of place |
| Table 2: Ecosystem services provided by soil and its biota. Based on Daily et al., 1997; Wall, 2004; Palm et al., 2007; and Dominati et al., 2010. | |
In particular, soils and its biota are essential for agricultural production and thus provision of a high proportion of our food. For example, at the turn of the millennium approximately 94% and 99% of our intake of protein and calories, respectively, originated from cultivated systems (FAOSTAT 2003). Soils are also important for the provision of many other important ecosystem services including water retention and infiltration rates, which help replenish ground water and provide clean drinking water while mitigating the effect of floods and droughts. At the same time, increased infiltration/retention rates decrease surface water runoff, which help minimize the loss of nutrients and soil erosion that are vital for sustainable agricultural production. Greater infiltration and retention rates also help prevent siltation and nutrient enrichment of rivers and lakes that may impair the functioning of these ecosystems. Moreover, soil biota are essential for biogeochemical cycling which supports plant production and also affect global climate regulation (primarily by influencing global carbon dynamics). Soils and the biomass therein function as a carbon storage system with about 2,500 petagrams (1 Pg = 1018 g) of carbon stored in soils globally (Woodward et al. 2009).
Brussaard et al. (1997) highlighted four main services (or functions) of soil biota: decomposition of organic matter, nutrient cycling, bioturbation, and suppression of soil borne diseases and pests. However, not all soil species contribute to each of these functions and thus soil biota are often categorized into functional groups (a functional group being defined as a set of species that perform the same function, see Coleman et al. 2004, or Bardgett 2005 for details). Some key functional groups are listed in Barrios (2007) and include microsymbionts, decomposers, elemental transformers, soil ecosystem engineers, soil-borne pests and pathogens, and microregulators. Plant roots also have a large impact on soil function, both physically and biologically, but as this article focuses on soil biota, plant roots will only be considered when there is a direct link between the biota and the plant root. Each of the functional groups mentioned above plays a particular role in the overall function and provisioning of ecosystem services of soils. For instance, microsymbionts, such as mycorrhizal-forming fungi and nitrogen-fixing bacteria, help plants acquire nutrients which can increase plant productivity; decomposers help release nutrients from organic material, which promotes nutrient cycling; soil ecosystem engineers may alter the physical soil structure (e.g., by burrowing and promoting the formation of micro- and macro-aggregates) which may increase soil stability, water holding capacity, water infiltration rates and aeration, whilst also limiting soil erosion. On the whole, a healthy soil with high functioning is dependent on the presence of all these functional groups, and a loss of species may impair functioning and provision of ecosystem services.
Evidence for a Biodiversity-Ecosystem Services Relationship in Soils
The previous paragraph highlights the importance of soil biota for soil function and ecosystem services. One of the key questions for maintaining continued provision of ecosystem services provided by soils and their biota is whether functioning depends on the number of species present (i.e., biodiversity), on key species, species traits (i.e., functional group, life-cycle and history, stress tolerance, etc.) or on the composition of the communities (Figure 2).
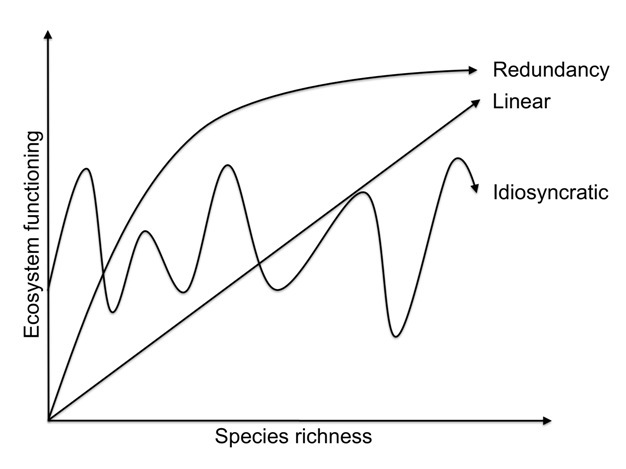
Figure 2: Graphic representation of three potential types of positive relationships between species richness and ecosystem functioning.
Note that such relationships can also be negative and that the relationships types are not mutually exclusive. The linear relationship would occur if the addition of any new species enhances functioning, whereas the redundancy relationship occurs if multiple species have the same influence on functioning. In the latter case, adding a new species only have a positive influence on functioning if it possess a trait not already found in the community, and the chance of this being the case decrease progressively as species richness increase. An idiosyncratic relationship indicates a system where species differ in their ability to enhance functioning, or where biotic interactions enhance (e.g. facilitation), or inhibit (e.g. competition), functioning. In this case the inclusion of single species have disproportionally large negative or positive impacts on functioning and the overall community composition is therefore more important for functioning than species richness per se.
© 2012 Nature Education Modified from Nielsen et al. (2011). All rights reserved.
One aspect of soils is its often-mentioned high biodiversity. Due to this high richness of species it has repeatedly been hypothesized that most species of soil biota are redundant (i.e., that the loss of a few species generally will not affect the rate of an ecosystem function or the services provided). There is some evidence in support of this hypothesis. A recent review of experiments exploring the relationship between carbon cycling and soil biodiversity concluded that although species richness, on average, led to greater functioning (measured as greater biomass, decomposition rates, and/or respiration), especially in very species poor communities, the relationship was predominantly idiosyncratic (or non-linear) (Nielsen et al. 2011). This review indicates that the presence of certain (key) species in soil communities has a disproportionately large effect on function compared to the number of species (see Nielsen et al. 2011). This pattern is well known in both natural and manipulated soil systems, and there are numerous examples of key species in soils. For example, the earthworm, Lumbricus terrestris, along with other species of earthworms, facilitates decomposition by fragmenting litter, thus increasing the surface area and preconditioning the litter for decomposition by other invertebrates and the microbial community. These earthworms redistribute the fragmented litter throughout the soil horizons, and by burrowing they enhance the soil structure (e.g., increase aeration, water infiltration). Other soil species have similarly large effects on soil function, such as certain species of potworms and termites, which have a similar function to L. terrestris, particularly in peatlands and savannas, respectively. Another example is the influence that rhizobia have on primary productivity within ecosystems. Rhizobia (a group of bacteria) are micro-symbionts of plants. Plant roots ‘infected' with the bacteria develop specialized nodules in which the bacteria live. Here they obtain sugars from the plants, which they use as an energy source to fix (transform) nitrogen from the air. Part of this nitrogen in turn becomes available for the plant, giving the plant a competitive edge and increasing the capacity for greater biomass production, particularly in nitrogen limited systems.
There is ample evidence that soil communities are adapted to specific environmental conditions and resource types. Hence, any changes in environmental conditions may have a negative effect on species richness and thus influence the performance of these communities, and severely impair functioning. The latter is most likely to occur in situations where the communities are species poor, such as managed systems or in extreme environments, which will limit the community's ability to adapt to new conditions. For example, some naturally occurring soils are very species poor due to external constraints (often due to unfavourable environmental conditions). The soils in the McMurdo Dry Valleys, Antarctica, a polar desert with an average annual temperate of -20°C, support only a few species of nematodes, tardigrades, rotifers, and microarthropods, and a limited microbial community (Figure 3). Here ecosystem functioning often relies on only one or a few species, and it has been hypothesized that the loss of species therefore are more likely to have negative impacts on functioning (Nielsen et al. 2011). Some evidence supports this notion. Barrett et al. (2008) showed how a climate induced 65% decrease in the abundance of the dominant nematode Scottnema lindsayae alone was estimated to lead to a 32% decrease in carbon turnover within Taylor Valley. Conversely, in this system even if the environmental conditions improve (i.e., temperature and water availability increase), soil functioning is unlikely to increase rapidly due to a limited regional species pool and a potential lack of species able to take advantage of increased resource input (i.e., primary productivity) and quality.

Figure 3: Wright Valley in the McMurdo Dry Valleys, Antarctica (77°31’S, 161°34’E).
The polar deserts of the Dry Valleys represent some of the harshest living conditions on Earth. The soils are generally very dry and highly saline, and inhabited by only a few species of higher soil fauna: microarthropods, nematodes, rotifers and tardigrades.
© 2012 Nature Education Courtesy of U.N. Nielsen. All rights reserved.
Despite the evidence for a large contribution of certain key species to ecosystem functioning there is also evidence that refutes the redundancy hypothesis and demonstrates that overall species richness has a positive influence on ecosystem functioning. Brussaard et al. (2007) concluded from a review of experiments and studies that there is great certainty in the ability of soil biodiversity to facilitate ecosystem stability towards stress and disturbance; that microbial diversity provides protection against soil borne disease; and that mycorrhizal diversity (a micro-symbiont) positively influences nutrient use efficiency, and in some cases water use efficiency. Moreover, some studies have shown that high species richness within functional groups may promote higher process rates and maintain higher species richness over time (Wohl et al. 2004), while the loss of a few rare microbes have been shown to impact plant-herbivore relationships aboveground (Hol et al. 2010). This suggests that the loss of species that may appear redundant or to have little impact on overall ecosystem functioning can have unexpected consequences. Hence, even though the biodiversity-ecosystem functioning relationship has been better explored in aboveground terrestrial and aquatic ecosystems (see, for example, Hooper et al. 2005, Cardinale et al. 2006) than in soil systems there is growing evidence for a similar relationship belowground.
We emphasize however, that the realized effect of increasing biodiversity on ecosystem function and services varies considerably between ecological studies both aboveground and belowground, and will be context dependent, influenced by environmental conditions and dependent on the spatial and temporal scale of investigation (e.g., Tylianakis et al. 2008, Hiddink et al. 2009, Bardgett & Wardle 2010, Nielsen et al. 2011). Moreover, many studies have been criticized for not being applicable to natural systems (see, for example, Raffaelli 2004, Naeem 2008). In particular, it has been highlighted that the majority of diversity-functioning studies, whether these were manipulating aboveground or belowground communities, use randomly assembled communities when species extinctions in natural systems are not random. This is critical as such differences could influence the outcome of said studies. For example, Bracken et al. (2008) showed how realistic increases in biodiversity of marine tide pools (based on naturally occurring communities) had a strong positive influence on nitrogen use efficiency, whereas random increases in species richness had no measurable effect on nitrogen uptake. A similar pattern may occur in soils. Moreover, evidence indicates species with particular traits are often lost more readily than others, which suggests that certain functions, in soils and elsewhere, may be particularly sensitive to species extinctions (Bardgett & Wardle 2010). Consequently, there is a potential for the loss of these functions due to even minor losses of species. It is critical that such patterns be considered in context for future studies investigating the biodiversity-ecosystem service relationship.
Theoretical Links Between Biodiversity and Ecosystem Functioning
So how can soil biodiversity promote ecosystem functioning and stability and help maintain the provision of ecosystem services? There are several hypotheses (not mutually exclusive) that may contribute to the answer. Although these hypotheses or mechanisms have mainly been coined through work performed in aboveground and aquatic systems, the evidence outlined in the previous section suggests that such mechanisms are likely to have a similar impact belowground. These mechanisms as noted from aboveground plant experiments primarily may be roughly grouped as those related to ‘sampling effects' (i.e., where a positive response to increased richness occurs due to increased probability of including highly influential species), and those related to ‘interaction effects' (i.e., where greater functioning or provision of services arise through interactions between species).
Sampling effects
Greater functioning in soils with species rich communities may prevail simply because such communities are more likely to contain species that have a disproportionally large influence on process rates (i.e., key species), or due to the presence of more specialized species that can utilize particular resources or niches, than species poor communities. The inclusion of more specialized species would allow greater resource use efficiency without any direct interactions between species. Moreover, diverse communities are more likely to include species that can cope with externally inflicted stresses or disturbances than species poor communities (sensu ‘the insurance hypothesis', see Tilman & Downing 1994, Yachi & Loreau 1999) and the former communities are therefore likely to show greater stability during such events (see Figure 4, and Ives & Carpenter 2007). The phrase ‘Portfolio-effect' was coined by Tilman et al. (1998) and compares communities with the stock market: having more stocks that respond differently to external stimuli (i.e., they are not correlated) will provide a buffering effect to market fluctuations and limit the loss of capital to stochastic events (Figure 4a). The same is true for communities, where a greater species pool will limit the risk of all species being unable to adapt to external stimuli, and therefore species rich communities will provide more stable functioning and biomass over time. Due to the usually high species richness observed in soils this is likely to be of particular importance in extreme soil environments such as the polar deserts where there are less species. Here ecosystem functioning will rely on fewer species, and each species is therefore likely to have a greater influence on overall functioning. Consequently, a loss of species may have a greater impact on functioning in species poor communities than in species rich communities. However, even though species rich communities generally are more stable with respect to biomass than species poor communities, the densities of individual species within these communities will be highly variable over time (i.e., stability depends on the temporal and spatial scale of investigation).

Figure 4: Diagram showing how the ‘Portfolio-effect’ (A) and community evenness (B) may lead to a positive diversity-ecosystem functioning relationship.
The ‘Portfolio-effect’ compares communities with a stock portfolio (Tilman et al., 1998). Consider two different portfolios: one with 2 stocks (left hand side) and one with 4 stocks (right hand side) each with a total value of $100 (top panel). A stochastic event causes a 50% reduction in the value of stock A (bottom panel). In the portfolio with only 2 stocks this leads to an overall loss of $25, whereas only $12.5 are lost in the portfolio with 4 stocks. Similarly, a decrease in the abundance of one species during disturbance events will have a larger impact in species poor communities than species rich communities, and functioning in diverse communities are therefore likely to show a lower response. Moreover, the relative abundance of species within communities may also influence how ecosystem functioning responds to disturbances. Consider two communities with four species each, but one is dominated by one species (low evenness; left hand side) while all four species are evenly represented in the other (high evenness; right hand side). A disturbance event leads to a 50% decrease in the abundance of species 1 in both communities. In the community dominated by one species this leads to a 30% reduction in overall abundance but only a 12.5% reduction in the community with high evenness. This is likely to result in a greater reduction in functioning in the community with high dominance than in the community with high evenness. Thus communities with many species and high evenness are likely to show smaller responses to external stimuli than communities with few species and high dominance by single species.
© 2012 Nature Education All rights reserved.
There is also increasing evidence for the same effect of community evenness (Figure 4b). Evenness refers to the relative abundance of species within a community, and communities that are made up of species with similar abundances are said to show high evenness. In contrast, communities dominated by one or a few species show low evenness. Hence, in ecosystems where communities show high natural evenness, stress or disturbances are unlikely to impair all species partaking in the provision of a service, making these communities more resistant/resilient towards disturbances. Communities with low evenness may on the other hand lose the dominant species, which is likely to play a significant role in local ecosystem processes, due to stress or a disturbance with subsequent impacts on ecosystem functioning.
Interaction effects
In contrast to the mechanisms listed in the above section, positive biodiversity-services relationships may also result through increased interactions between species (e.g., competition, mutualism, complementarity, facilitation, etc.) in more species rich communities. Greater competition between species could increase resource use efficiency by forcing species to become more specialized in their resource use, and positive interactions such as mutualism and facilitation can lead to greater productivity of single species. As noted earlier, this has been studied primarily in aquatic and aboveground systems, but similar interactions are likely to occur in soils. For instance, as outlined earlier microbial symbionts of plant roots can increase nutrient acquisition by the plants, while the microbes in turn receive sugars produced during plant photosynthesis (an example of a mutualistic relationship between two organisms), and the fragmentation and preconditioning of organic material by invertebrates can facilitate microbial decomposition.
Increased species richness can also alter interaction strengths between species and through this influence the stability of the communities. One proposed mechanism that may underlie the diversity-stability relationship is the ‘Weak-interaction effect' (McCann 2000, Figure 5). In communities with a high proportion of weak interactions (i.e., predators are not strongly dependent on one type of prey), it is more unlikely to observe run-away consumption of certain prey items, which would alter community dynamics and lead to unstable conditions. Overall, interactions will on average be weaker in a highly diverse community than in a species poor community, and thus diverse communities are likely to be more stable. Although our knowledge of interactions within the soil food web is limited, there is evidence for a high proportion of weak interactions in soils (de Ruiter et al. 1995, 1998), which would contribute to the stability of these food webs. If the biodiversity of such food webs were to be impaired, stronger predator-prey interactions are likely to become more dominant, resulting in more un-stable soil food webs.
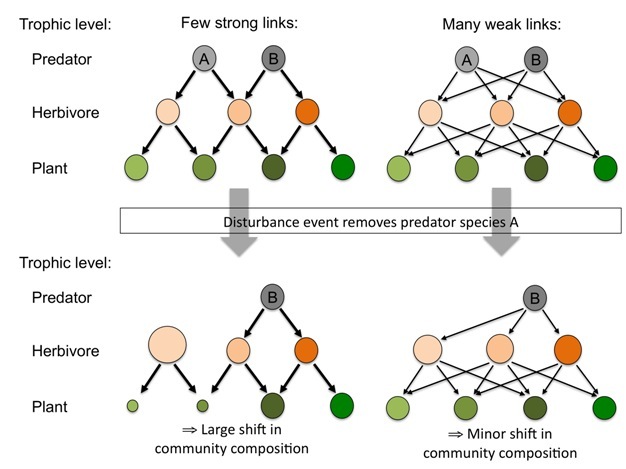
Figure 5: Diagram showing how the dominance of weak interactions within simplified food webs may lead to more stable communities.
The top pane shows the interactions and their strength between predator and herbivores, and between herbivores and plants before (top) and after (bottom) a disturbance event. Arrows signify interactions, and larger arrows indicate stronger interactions, and the size of the circles indicates the abundance of each organism. One food web (left hand side) is dominated by a few strong interactions (or links) whereas the other food web (right hand side) is dominated by more but weaker interactions. A stochastic disturbance event then removes predator A from each food web. In the food web dominated by strong interactions this leads to the release of predation of one of the herbivores, which subsequently become more abundant and this causes a decrease in the two species of plants that this species feed on. The removal of one predator from this food web therefore causes a large shift in community composition of herbivores with cascading effects on plant species abundances. In contrast, the removal of predator A from the food web dominated by weak interactions has little impact on the relative abundance of herbivores (at most a slight increase for all species), and there are no significant cascading effects. In natural communities weak interactions are more likely to dominate as the number of species increase, and because of this species rich communities may show less response to disturbance events than species poor communities.
© 2012 Nature Education All rights reserved.
All of the mechanisms mentioned in this and the previous section may influence biodiversity-ecosystem services relationships but they are not mutually exclusive, nor is this an exhaustive list of potential mechanisms underlying the biodiversity-ecosystem service relationship. Therefore any combination of these (and other) factors may act in concert to produce the observed biodiversity-ecosystem services relationships making it difficult to interpret which underlying mechanism(s) is responsible. However as we have shown, there is much theoretical, as well as empirical, support of the proposed positive relationship between soil biodiversity and ecosystem functioning.
Summary
Soil biodiversity is the basis for many ecosystem services that people depend upon such as food and fiber. Whether soil species loss will affect the provision of ecosystem services has been of increasing interest and research. Currently, there is ample evidence for an on average positive effect of soil biodiversity on ecosystem functioning and through this, ecosystem services, but the relationship is often idiosyncratic and context dependent. Hence, it appears that the soil biodiversity-ecosystem services relationship is more dependent on the inclusion of certain key species and/or species with particular traits than species richness per se (Brussaard et al. 2007). However, some evidence suggests that species richness is important in certain soils. For example, positive biodiversity-ecosystem function relationships are more pronounced in species poor ecosystems than species rich ecosystems (Nielsen et al. 2011), and such ecosystems may therefore be more sensitive to species loss. Moreover, there is some evidence that functionally redundant species can be important for functioning (Wohl et al. 2004), and we have yet to determine if the continued provision of multiple ecosystem services (i.e., multifunctionality) simultaneously is more responsive to biodiversity changes as has been observed in grasslands (Hector & Bagchi 2007). In contrast, the stability (i.e., resistance and resilience) of ecosystem services to disturbance and stress generally appear to be more strongly related to species richness (Brussaard et al. 2007), a pattern that has also been observed in many other habitats (Ives & Carpenter 2007). Hence, even though the number of species present in soils may not always appear to have a strong direct influence on ecosystem services we conclude that soil biodiversity is important for the provision of ecosystem services. Thus, conserving soil biodiversity is of high value to ensure the continued provision of ecosystem services.
Glossary
biogeochemical cycling: The cycling of nutrients by biota and chemical weathering through organic and mineral pools including living organisms (plants and animals), soils, oceans, atmosphere etc.
bioturbation: The physical rearrangement of soil particles by fauna and plant roots. Some organisms such as the earthworms have a high impact on the soil structure by burrowing and can create or alter the living space for other organisms.
complementarity: Building on the idea of niche differentiation or that species differ in their resource requirements, complementarity suggests that a more diverse community can use resources more efficiently and through this enhance productivity.
decomposition: The breakdown of complex organic compounds to more simple compounds that can be readily utilized by plants and other organisms. Decomposition is essential for nutrient cycling.
elemental transformers: Microorganisms that can obtain energy through the transformation of elements including the bacteria that take part in nitrification and denitrification.
facilitation: A mechanism through which one organism promotes or enhances the growth or survival rate of another organism by altering the physical and/or the biotic environment.
global carbon dynamics: The transfer of carbon between global pools water, soil, atmosphere, vegetation.
litter: Organic material derived from vegetation including leaves, twigs, roots etc.
microregulators: Organisms, such as grazers, parasites and predators, responsible for regulating the population size or biomass of other organisms.
microsymbionts: Mainly referring to microorganisms that form associations with plants. Many microsymbionts such as nitrogen fixing bacteria and arbuscular mycorrhizal fungi help the plant acquire nutrients in exchange for plant organic carbon generally provided in the form of sugars.
mutualism: An interaction between two organisms in which both partners benefit.
primary production: Organic material produced by plants and other photosynthesizing organisms.
resource use efficiency: The efficiency with which an organism can take up and utilize a resource.
soil horizons: Soils can be divided into a series of different horizons. Overall, we differentiate between the organic and the mineral horizons. The organic horizon is generally comprised of a litter (or L) layer, a layer of fragmented (F) litter a humus (H) layer. The L layer is dominated by fresh litter, whereas the F layer is dominated by fragmented and partly decomposed litter and the H layer by humified (or greatly decomposed) litter and the latter often mixed with mineral soil from the lower soil layers. The litter layer is the zone of greatest biotic activity and has the highest abundances of soil fauna. The mineral horizons are all less influenced by organic matter and the influence tend to decrease with increasing depth.
stochastic: Random or non-deterministic event.
References and Recommended Reading
Balvanera, P. et al. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters 9, 1146-1156 (2006).
Barbier, E. B. Scarcity and Frontiers: How Economies Have Developed Through Natural Resource Exploitation. Cambridge, UK: Cambridge University Press, 2011.
Bardgett, R. D. The Biology of Soils: A Community and Ecosystem Approach. Oxford, UK: Oxford University Press, 2005.
Bardgett, R. D. & Wardle, D. A. Aboveground-Belowground Linkages: Biotic Interactions, Ecosystem Processes, and Global Change. Oxford Series in Ecology and Evolution. Oxford, UK: Oxford University Press, 2010.
Barrett, J. E. et al. Decline in a dominant invertebrate species contributes to altered carbon cycling in a low-diversity ecosystem. Global Change Biology 14, 1734-1744 (2008).
Barrios, E. Soil biota, ecosystem services and land productivity. Ecological Economics 64, 269-285 (2007).
Bracken, M. E. S. et al. Functional consequences of realistic biodiversity changes in a marine ecosystem. Proceedings of the National Academy of Sciences of the United States of America 105, 924-928 (2008).
Brussaard, L. et al. Soil biodiversity and ecosystem functioning in soil. Ambio 26, 563-570 (1997).
Brussaard, L., de Ruiter P. C., & Brown, G. G. Soil biodiversity for agricultural sustainability. Agriculture, Ecosystems and Environment 121, 233-244 (2007).
Cardinale, B. J. et al. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 443, 989-992 (2006).
Coleman, D. C., Crossley, Jr., D. A., & Hendrix, P. F. Fundamentals of Soil Ecology 2nd ed. Burlington, MA: Elsevier Academic Press, 2004.
Costanza, R. et al. The value of the world's ecosystem services and natural capital. Nature 387, 253-260 (1997).
Daily, G. C. et al. Ecosystem services: Benefits supplied to human societies by natural ecosystems. Issues in Ecology 2, 1-18 (1997).
De Ruiter, P. C., Neutel, A-M., & Moore, J. C. Biodiversity in soil ecosystems: The role of energy flow and community stability. Applied Soil Ecology 10, 217-228 (1998).
De Ruiter, P. C., Neutel, A-M., & Moore, J. C. Energetics, patterns of interaction strengths, and stability in real ecosystems. Science 269, 1257-1260 (1995).
Dominati, E., Patterson, M., & Mackay, A. A framework for classifying and quantifying the natural capital and ecosystem services of soils. Ecological Economics 6, 1858-1868 (2010).
European Soil Bureau Network. Soil Atlas of Europe. Ispra, Italy: European Commission, 2005.
FAOSTAT. Food and Agriculture Organization of the United Nations, Statistical Databases, Online at http://faostat.fao.org (2003).
Foley, J. A. et al. Global consequences of land use. Science 309, 570-574 (2005).
Hector, A. & Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 448, 188-191 (2007).
Hiddink, J. G. et al. Context dependency of relationships between biodiversity and ecosystem functioning is different for multiple ecosystem functions. Oikos 118, 1892-1900 (2009).
Hol, W. H. G. et al. Reduction of rare soil microbes modifies plant-herbivore interactions. Ecology Letters 13, 292-301 (2010).
Hooper, D. U. et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecological Monographs 75, 3-35 (2005).
Ives, A. R. & Carpenter, S. R. Stability and diversity of ecosystems. Science 317, 58-62 (2007).
May, R. M. Ecological science and tomorrow's world. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 41-47 (2010).
McCann, K. S. The diversity-stability debate. Nature 405, 228-233 (2000).
MEA. Millennium Ecosystem Assessment: A Framework for Assessment. Washington, DC: Island Press, 2003.
MEA. Millennium Ecosystem Assessment: Ecosystems and Human Well-being: Synthesis. Washington, DC: Island Press, 2005.
Naeem, S. Advancing realism in biodiversity research. Trends in Ecology and Evolution 23, 414-416 (2008).
Naeem, S. Species redundancy and ecosystem reliability. Conservation Biology 12, 39-45 (1998).
Nelson, E. et al. Modeling multiple ecosystem services, biodiversity conservation, commodity production, and tradeoffs at landscape scales. Frontiers in Ecology and the Environment 7, 4-11 (2009).
Nielsen, U. N. et al. Soil biodiversity and carbon cycling: A review and synthesis of studies examining diversity-function relationships. European Journal of Soil Science 62, 105-116 (2011).
Palm, C. et al. Soils: A contemporary perspective. Annual Review of Environment and Resources 32, 99-129 (2007).
Ptacnik, R. et al. Diversity predicts stability and resource efficiency in natural phytoplankton communities. Proceedings of the National Academy of Sciences of the United States of America 105, 5134-4138 (2008).
Raffaelli, D. How extinction patterns affect ecosystems. Science 306, 1141-1142 (2004).
Tilman, D. Causes, consequences and ethics of biodiversity. Nature 405, 208-211 (2000).
Tilman, D. & Downing, J. A. Biodiversity and stability in grasslands. Nature 365, 363-365 (1994).
Tilman, D., Lehman, C. L., & Bristow, C. E. Diversity-stability relationships: Statistical inevitability or ecological consequence? The American Naturalist 151, 277-282 (1998).
Tylianakis, J. M. et al. Resource heterogeneity moderates the biodiversity-function relationship in real world ecosystems. PLoS Biology 6, e122 (2008).
Van der Putten, W. H. et al. "The sustainable delivery of goods and services provided by soil biota," in Sustaining Biodiversity and Ecosystem Services in Soils and Sediments, ed. D. H. Wall, pages 15-43 (San Francisco, CA: Island Press, 2004).
Wall, D. H. ed. Sustaining Biodiversity and Ecosystem Services in Soils and Sediments. SCOPE 64 Washington, DC: Island Press, 2004.
Woodward, F. I. et al. Biological approaches to global environment change mitigation and remediation. Current Biology 19, R615-R623 (2009).
Wohl, D. L., Arora, S., & Gladstone, J. R. Functional redundancy supports biodiversity and ecosystem function in a closed and constant environment. Ecology 85, 1534-1540 (2004).
Yachi, S. & Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America 96, 1463-1468 (1999).










