



















« Prev Next »
What is Migration?
Most people think of migration as the seasonal movement of a flock of birds between their breeding and non-breeding sites. In fact, bird migration is probably the biological phenomenon that has attracted the most interest among non-scientists, and has one of the longest traditions of scientific investigation in biology (Berthold 2001). However, there are many other forms of animal migration, including journeys between east and west, complex round-trips involving land and ocean, altitudinal journeys up and down mountains, and vertical movements through the water column of oceans and lakes (Hoare 2009). What sets migration apart from other forms of movements is that migration typically involves travelling from one type of habitat to another (Aidley 1981).
Who Migrates?
Migrating animals are found in all major branches of the animal kingdom. They include taxa as diverse as fish, crustaceans, amphibians, reptiles, insects, mammals and slime moulds (Bowlin et al. 2010). The distance covered during some of the journeys, and the altitude at which they can take place, are simply astonishing (Table 1). Let’s take a closer look at the migrations undertaken by a bird, an insect and a mammal.
| Table 1. Guinness records held by animal migrants | ||
|
Smallest migrant |
1–2 mm long |
Zooplankton (crab larvae, copepods, etc.) |
|
Largest migrant |
24–27 m long |
Blue whale (Balaenoptera musculus) |
|
Longest mammal migration |
Up to 8,500 km each way |
Humpback whale (Megaptera novaeangliae) |
|
Longest insect migration |
Up to 4,750 km in the autumn |
Monarch butterfly (Danaus plexippus) |
| Longest recorded round-trip |
80,000 km |
Artic tern (Sterna paradisaea) |
| Highest migration altitude |
9,000 asl |
Bar-headed goose (Anser indicus) |
| Source: Hoare 2009, Egevang et al. 2010 | ||

Figure 1: Bar-tailed Godwit feeding on a bivalve on northward migration at Yalu Jiang River, Yellow Sea, China
© 2010 Nature Education Courtesy of Phil Battley. All rights reserved. 
Perhaps the best-known insect migrant is the monarch butterfly (Danaus plexippus) (Figure 3a). At the end of every summer, more than 100 million individuals sweep across North America to overwinter up to 4,750 km further south in California and Mexico. The complete migration requires an intergenerational relay. No single individual makes the entire round trip, as the duration of these journeys exceeds their lifespan. Instead, the females lay eggs during the migration, from which the next generation of migrants emerges. Monarchs specialize on milkweeds (Asclepiadaceae), which contain substances that are poisonous to vertebrates and thus protect the monarch from many potential enemies (Dingle 1996).
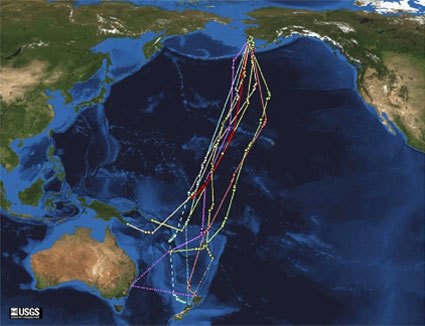
Figure 2: Flight tracks of nine bar-tailed godwits on southward migration equipped with satellite transmitters
One female flew directly from Alaska to the non-breeding grounds in New Zealand on a 11,680 km non-stop journey across the Pacific Ocean, which lasted over 8 days (Gill et al. 2009).
© 2010 Nature Education Courtesy of US Geological Survey. All rights reserved.
The common name of the southern right whale (Eubalaena australis) (Figure 3b) refers to the fact that they were considered the ‘right’ whale to hunt. They are slow-moving and have large amounts of blubber, which meant that they floated after being killed. Many tens of thousands were slaughtered until the international whaling ban in 1935. Southern right whales migrate from their Antarctic feeding areas to temperate breeding areas along the costs of Chile and Argentina, southern Africa, and Australia and New Zealand, covering 2,500 km each way. Their migration is fuelled entirely by fat accumulated during their four-month stay in the icy Southern Ocean around Antarctica, where they skim the surface waters for zooplankton. Amazingly, they will not feed until their return a year later (Hoare 2009).

Figure 3: (A) Overwintering monarch butterflies gather at such high densities that the branches of their trees bend with their weight. (B) When the Southern right whales arrive on their feeding grounds around Antarctica in December they are often near starvation. But they are able to double their body weight after a short stay in the food-rich polar seas.
© 2010 Nature Education 3A courtesy of Raina Kumra; 3B courtesy of Courtesy of Michael Catanzariti. All rights reserved.
Why Migrate?
Migration is an adaptive response to the seasonal or geographic variation of resources (Gauthreaux 1982). The annual cycle of the seasons produces large differences in the duration and intensity of solar energy received in each hemisphere at any given time. Many migrants take advantage of favorable food and weather conditions offered in certain areas during very limited time periods. For instance, migratory birds like the bar-tailed godwit that breed at high latitudes (e.g., in the arctic tundra) exploit the extraordinary abundance of food during a few weeks in early summer and profit from long days, which allow them to extend foraging time. By leaving these areas after breeding, they avoid the northern winters with short days, low temperatures, and low food availability (Pulido 2007). In other cases, the resources needed in different life stages may be found in different locations. For example, young salmon migrate to sea to take advantage of the great abundance of food and high growth potential available there, but must someday return upstream to the small rocky tributaries required for spawning, a trip sometimes involving thousands of kilometres (Figure 4; Dingle 1996).

Figure 5: Migrating kokanee salmon Oncorhynchus nerka in Meadow Creek, British Columbia, Canada
© 2010 Nature Education Courtesy of Margaret A. Warren. All rights reserved.
Preparing for Migration
Some migratory animals forage along their migratory routes, while others, such as the bar-tailed godwit crossing the Pacific Ocean, do not. Most migrants therefore feed intensively prior to departing for their journey to boost their fuel stores. This behaviour is known as hyperphagia, which is triggered by an internal circannual rhythm, and is seen in species as varied as monarch butterflies, caribou, and baleen whales. The main source of energy used for migration is fat. Fat stores about 8–10 times more energy than the equivalent mass of either carbohydrate or protein, making fat the fuel of choice (Jenni & Jenni-Eiermann 1998). Migrants are capable of storing and mobilizing large amounts of fat: birds that fly long distances can double their body weight prior to departure, and a monarch butterfly can store up to 125% of its lean dry weight as fat (Dingle 1996; Brower et al. 2006).
Accumulating energetic reserves is not the only physiological change taking place. As migration is energetically expensive, some migrants minimize excess weight and maximize flight efficiency by adjusting the size of their internal organs. In some avian migrants, organs related to flight, such as heart and flight muscles, increase in size in preparation for migration (Piersma 1998; Ramenofsky & Wingfield 2007). By contrast, organs related to feeding (e.g., stomach, gut, liver, and kidneys) become smaller before departure and return to normal size upon arrival, when feeding and digestion resume (Piersma & Gill 1998). Similar changes are observed in some insects: for example, monarch butterflies migrating south through North America in the autumn have no sexual organs — these only develop the following spring in the surviving individuals (Brower et al. 2006).
Accumulating energetic reserves is not the only physiological change taking place. As migration is energetically expensive, some migrants minimize excess weight and maximize flight efficiency by adjusting the size of their internal organs. In some avian migrants, organs related to flight, such as heart and flight muscles, increase in size in preparation for migration (Piersma 1998; Ramenofsky & Wingfield 2007). By contrast, organs related to feeding (e.g., stomach, gut, liver, and kidneys) become smaller before departure and return to normal size upon arrival, when feeding and digestion resume (Piersma & Gill 1998). Similar changes are observed in some insects: for example, monarch butterflies migrating south through North America in the autumn have no sexual organs — these only develop the following spring in the surviving individuals (Brower et al. 2006).
Navigation During Migration
Visible Cues
The most obvious cues used by migrants to find their way are visible cues such as local topography. Some species, including birds, insects and crabs, are able to make use of polarized light patterns, which are formed when light is scattered by airborne particles. As the sun’s position shifts throughout the day, the pattern of polarized light in the sky changes, allowing these species to find their way on cloudy days when the sun is not visible. When travelling at night, some migrants use the position of the stars based on the rotation of the night sky about a fixed point. In the northern hemisphere, this point is the Pole Star.
Invisible Cue
Other cues used for orientation are invisible, such as olfactory and magnetic cues. Salmon migrating in the open ocean can find the mouth of their natal river by constantly sampling the seawater for changing concentrations of mineral salts. Magnetic cues are used by a broad range of species, including birds, butterflies, salamanders, lobsters, bats, whales, turtles, and sharks. The Earth, which behaves like a giant magnet, has a doughnut-shaped magnetic field, made up of elliptical force lines running between the magnetic north and south poles. Depending on the species, the magnetic compasses used by animals detect the changing angle of the magnetic force lines, the north-south polarity, or even the strength of the Earth’s magnetic field.
What Triggers Migration?
In birds, there is a clear association between day length and the onset of migration. Dark-eyed juncos (Junco hyemalis) and American crows (Corvus brachyrhynchos) kept in artificially lengthened light periods move north when released, while controls, kept under natural light conditions, move south (Dingle 1996). Hormones are also known to play a role in controlling migration. Bird migration is thought to depend on gonadal condition in spring, but not in fall. In many insects, changes in the activity of juvenile hormone are responsible for transitions between migratory and reproductive life stages, and the gregarious swarms of locusts are under the influence of the hormone serotonin. The connections between day length, annual reproductive cycle and migration are now well established in birds, fishes, and other migrants.
Taking Advantage of Outside Help with Migration
Many ocean currents follow definite and reliable routes, and marine migrants often taken advantage of them. In summer, leatherback turtles (Dermochelys coriacea) pick up the warm Gulf Stream to glide across the North Atlantic to the coasts of northwest Europe. Likewise, throughout the tropics, the movements of manta rays (Manta birostris) and whale sharks (Rhincodon typus) show a clear correlation with warm currents. The larvae of countless pelagic fish, molluscs, and crustaceans rely on currents to drift away from their spawning areas.
For aerial migrants, ideal migration conditions are a sustained tailwind, a cloudless sky and low temperatures. Cool air prevents hard-working pectoral muscles from overheating, which is one of the reasons why some birds migrate at night, particularly when crossing deserts, where the extreme heat at midday could be fatal.
For aerial migrants, ideal migration conditions are a sustained tailwind, a cloudless sky and low temperatures. Cool air prevents hard-working pectoral muscles from overheating, which is one of the reasons why some birds migrate at night, particularly when crossing deserts, where the extreme heat at midday could be fatal.
Human History of Animal Migration
The earliest evidence of humans being aware of animal migration dates back to the Stone Age. Some rock art images, portraying animals moving across the African savannah, are as old as 20,000 years. They were produced by nomadic hunter-gatherers, and could have served as a field guide to potential food, or as a visual record of good hunting areas.
The philosophers of Ancient Greece were the first to develop a theory of migratory behaviour in animals, even though their conclusions nowadays seem rather fanciful. Aristotle (384–322 BCE) realized that a number of birds were migratory, and his creative explanation for the sudden disappearance of summer migrants, such as warblers and swallows, was that they had morphed into different species present in winter. His idea of transmutation persisted well into the Middle Ages in Europe. According to another theory, which was still widely believed in the 1800s, migratory birds disappeared into the mud at the bottom of ponds and lakes where they would spend the winter. Since then, our understanding of migration has indeed improved.
How Migration Is Studied
Bird Banding
Instrumental in studying migration were banding (also called ringing) studies. Banding dates back to 1899, when Danish teacher Hans Christian Mortensen visited European starling (Sturnus vulgaris) nests and gave each nestling an aluminum leg band engraved with a return address and a unique serial number. If anyone encountered one of his banded birds, they could send back information on the time and place where it was found. Since 1899, more than 200 million birds are estimated to have been banded worldwide, of which only a fraction have been recovered. However, even a recovery rate as low as 1 in 300 — the average for small birds — still provides valuable insights into the routes that migrants take. Alternatives to banding include labeling with dye and attaching plastic tags to the neck or back, which is also used for mammals (Hoare 2009).
Radar
The rapid development of radar during the Second World War enabled actual migratory journeys to be plotted for the first time. Modern radar is powerful enough to pinpoint the height, speed, and wing beat rate of individual birds and bats. Its aquatic equivalent — sonar — can detect shoals of fish moving underwater.
Tracking Devices
It is possible to study animal movements without directly observing an individual. Fitting animals with radio transmitters and using hand-held or stationary antennas allows following their whereabouts within a range of several kilometers. Alternatively, satellite transmitters beam signals to orbiting satellites, which then relay the data back to computers on the ground. GPS tags use the satellites of the Global Positioning System to record data such as location and time. They can be attached to mammals with a neck collar or to medium-sized birds with a backpack. Even smaller are geolocators, which are miniature light level loggers that can record sunset and sunrise, from which the location of the individual can be reconstructed. Some weigh less than 1 g and can last for many years. However, they need to be retrieved to access the data they store.
Stable Isotopes
It is now possible to measure the amount of stable isotopes such as deuterium (a form of hydrogen), oxygen, carbon, nitrogen, and sulfur in the tissue of migrants. Isotope levels in the plumage of a migratory bird match that of the vegetation of its breeding ground and can therefore serve as an indication of its place of birth (Hobson 2005). The same technique has been used to establish the hatching place of individual monarch butterflies wintering in Mexico.
Summary
Animal migration involves travelling from one type of habitat to another, which is often linked to the cycle of the seasons. Preparing for the migratory journey, which poses extreme energetic demands on the animal, usually involves a number of physiological changes, such as storing large amounts of fat and reducing the size of the organs that are not needed during migration. Migrants use a variety of cues to find their way, such as the magnetic field of the earth, changing concentration of minerals in the ocean water or polarized light. The breath-taking nature and raw beauty of animal migration has inspired humans for the past 20,000 years and continues to do so to this day.
Acknowledgments
Thanks to Alex Gerson, Chris Guglielmo, Alex Macmillan and Eddy Price for valuable comments.
References and Recommended Reading
Aidley, D. J. Animal Migration. Cambridge, UK: Cambridge University Press, 1981.
Berthold, P. Bird Migration: A General Survey. Oxford, UK: Oxford University Press, 2001.
Bowlin, M. S., Bisson, I. A. et al. Grand challenges in migration biology. Integrative & Comparative Biology (2010).
Brower, L. P., Fink, L. S. et al. Fueling the fall migration of the monarch butterfly. Integrative & Comparative Biology 46, 1123–1142 (2006).
Dingle, H. Migration: The Biology of Life on the Move. New York, NY: Oxford University Press, 1996.
Egevang, C. et al. Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proceedings of the National Academy of Sciences 107, 2078–2081 (2010).
Gauthreaux, Jr., S. A. The ecology and evolution of avian migration systems. Avian Biology 6, 93–68 (1982).
Gill, R. E. J., Tibbitts, L. T. et al. Extreme endurance flights by landbirds crossing the Pacific Ocean: ecological corridor rather than barrier? Proceedings of the Royal Society B, Biological Sciences 276, 447–457 (2009).
Hoare, B. Animal Migration. Remarkable Journeys by Air, Land and Sea. London, UK: Natural History Museum, 2009.
Hobson, K. A. Flying fingerprints: Making connections with stable isotopes and trace elements. In Birds of Two Worlds: the Ecology and Evolution of Migratory Birds. eds. Greenberg, R. & Marra, P. P. (Baltimore, MD: Johns Hopkins University Press, 2005): 235–246.
Jenni, L. & Jenni-Eiermann, S. Fuel supply and metabolic constraints in migrating birds. Journal of Avian Biology 29, 521–528 (1998).
Piersma, T. Phenotypic flexibility during migration: optimization of organ size contingent on the risks and rewards of fueling and flight? Journal of Avian Biology 29, 511–520 (1998).
Piersma, T. & Gill, R. E. J. Guts don't fly: small digestive organs in obese Bar-tailed Godwits. Auk 115, 196–203 (1998).
Pulido, F. The genetics and evolution of avian migration. BioScience 57, 165–174 (2007).
Ramenofsky, M. & Wingfield, J. C. Regulation of migration. BioScience 57, 135–143 (2007).










