




















« Prev Next »

Deep inside a hive, a honey bee (Apis mellifera) dances wildly. Other bees cluster around, touching her body with their forelegs and antennae as she dances. Then one by one, the observers leave the dance, head to the hive entrance, and take flight in the same direction. The dancing bee has just communicated the direction and distance to an abundant food source (Figure 1). Within the hour, the foragers are returning, ready to relay the whereabouts of the resource to other foragers through dances of their own.

Figure 1: Waggle dance.
Honey bee (Apis mellifera) foragers communicate the location of rich food sources to other members of the hive through dances. When the food source is located more than 50 m from the hive, they perform the waggle dance (von Frisch 1974). The forager waggles its abdomen from side to side as it walks in a straight line on the comb. The angle at which the bee moves relative to vertical relates information about the direction of the food source relative to the sun. The bee then circles around before repeating the waggle run, dancing in a figure-eight as long as it has other bees in attendance. Remarkably, the foragers that attend the dance account for movement of the sun when they set out in search of the food source (Dyer 2002).
© 2012 Nature Education All rights reserved. 
Honey bees provide an intriguing example of how animals exhibit extraordinary abilities to navigate their worlds. Some of these behaviors remind us of our own abilities, whereas others extend beyond human cognition. Either way, these amazing behaviors beg a number of questions. Do honey bees possess a mental map of their environment? What about other animals? What kind of cognitive capacities do they need to process information they receive from their environment?
Studying Animal Cognition
Studying the animal mind poses unique challenges because we project ourselves onto animals, as seen in the example of Clever Hans. Hans was a German horse who became a global phenomenon in the early 1900s because of his ability to answer mathematical questions (Figure 2). His ability to add, subtract, multiply and divide (by stomping the correct number of times) was examined by a number of professionals and determined to be real.

Figure 2: Clever Hans.
Clever Hans stomping out his response to a mathematical question.
© 2012 Nature Education All rights reserved.
Although many people were thoroughly convinced of Hans's mathematical prowess, some remained skeptical. Oskar Pfungst made careful observations of Hans's behavior and discovered that although Hans could correctly answer questions from a variety of people, he could do so only if the questioner were visible. Pfungst discovered that when people asked Hans a question, they slightly moved their heads when the correct answer was presented. Hans was indeed clever: he was attuned to the subtle subconscious body language of the people around him (Pfungst 1911).
Clever Hans taught comparative psychologists an important lesson. Avoiding pitfalls such as this requires carefully controlling experimental studies and following Lloyd Morgan's canon: accept the simplest explanation for a behavior in favor of a more complex cognitive process.
Cognitive Capacities
The physical world poses a number of problems for animals to solve. On a daily basis, animals must find food, avoid predators, and seek shelter. Solving these problems requires cognitive capacities. Cognition involves processing information, from sensing the environment to making decisions based on available information. Such cognitive capacities include, among others, the ability to navigate through space, account for the passage of time, determine quantity, and remember events and locations. Here, we explore a sample of the cognitive capacities animals use in their daily lives.
Where am I?
Most animal species move about in their habitat, which requires navigating between locations. Navigation occurs over different spatial scales, from centimetres to thousands of kilometres, and different mechanisms are used at different scales. At small scales, in which animals navigate around their home territory, they can use dead reckoning, landmarks, and cognitive maps to navigate.
Dead reckoning involves estimating the distance and direction one has traveled. For instance, desert ants (Cataglyphis spp.) track how far away and in what direction they have traveled from home in order to return home after searching for food (Wehner 2003; Figure 3a).
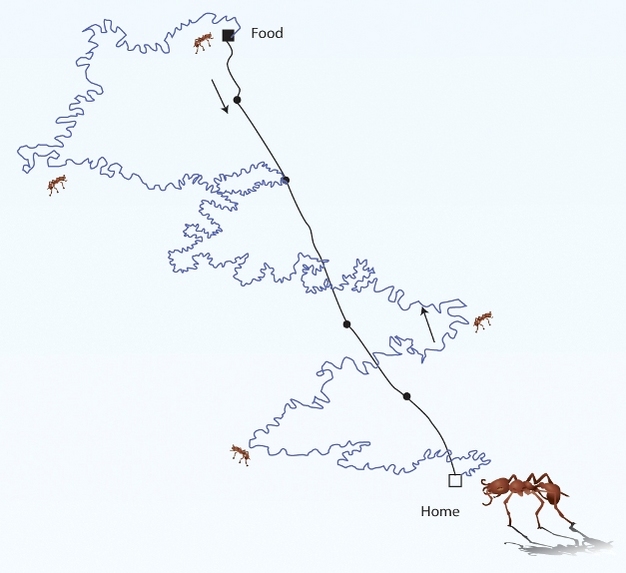
Figure 3a: Navigation.
Ants use dead reckoning to locate their nests after convoluted foraging trips.
© 2012 Nature Education All rights reserved.
Other species use landmarks to guide their movement. Animals can learn the relationships among landmarks such as rocks, trees, or other large objects to triangulate their position. Landmarks are often the primary cues that animals use to locate their nests. For example, after digger wasps (Philanthus triangulum) leave their nests they circle around the entrance to orient themselves to local landmarks. When the landmarks are moved several centimetres away, the returning wasps land where the nest entrance should be relative to the landmarks and have difficulty finding their nests (Tinbergen 1951; Figure 3b).

Figure 3b: Navigation.
In Tinbergen's (1951) experiments, digger wasps were unable to locate their nest entrances after he moved the landmarks surrounding the nests.
© 2012 Nature Education All rights reserved.
Finally, some animals may use a cognitive map to navigate. A cognitive map involves a mental map-like representation of the environment. Though controversial and difficult to demonstrate, honey bees show some evidence of using cognitive maps; when they are physically displaced to a new foraging location, they return home via a direct route. That is, they take a shortcut, suggesting that they possess a cognitive map of their territory (Menzel et al. 2005).
Many migrating species navigate over long distances. Arctic terns (Sterna paradisaea) travel nearly 80,000 km a year between feeding and mating areas (Egevang et al. 2010). How do Arctic terns and other migrating species navigate such enormous distances? Many species use something similar to global positioning systems that are based on a sun compass or the earth's magnetic field. A sun compass is the ability to use the sun's position in the sky to determine direction, accounting for both daily and seasonal changes in the sun's position (Alerstam et al. 2001). Honey bees appear to use a sun compass when navigating to their foraging sites (Figure 1). Birds, reptiles, amphibians, and molluscs have also been shown to orient themselves based on the earth's magnetic field (Lohmann & Lohmann 1993; Figure 4). The precise mechanisms enabling such navigation are still under investigation.
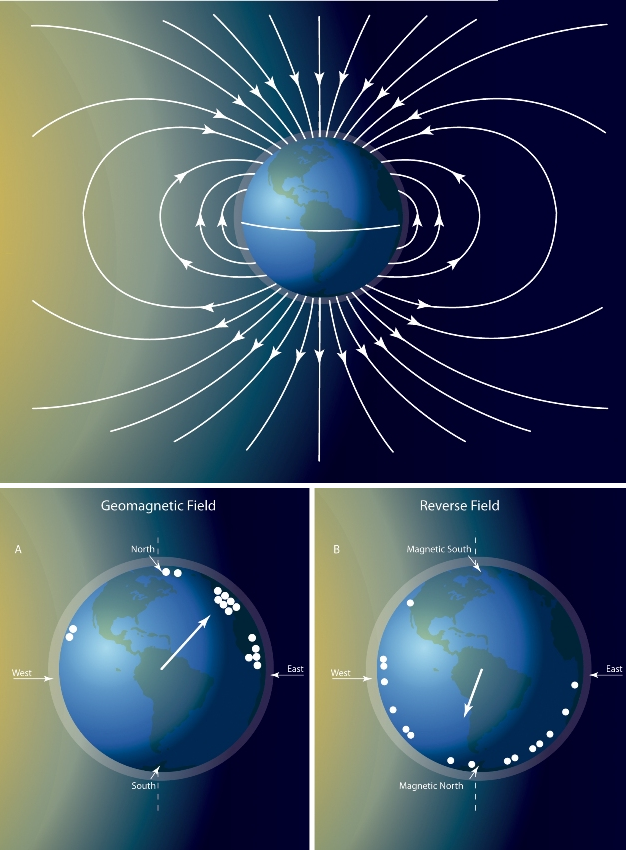
Figure 4: Detection of Earth's magnetic field.
(a) Earth's magnetic field generally runs in a north-south direction, providing a cue animals can use to orient their bodies during migration. (b) To test the ability to perceive the magnetic field, animals, such as leatherback sea turtles, are placed in a magnetic coil that reverses the polarity. These turtles reverse their direction of travel in the coils, suggesting that they use magnetic fields to navigate.
© 2012 Nature Education All rights reserved.
Telling Time
Time influences an animal's environment over periods ranging from milliseconds to decades. Annual cycles, in particular, are important for migration, hibernation, caching, mating, and raising young. Though temperature may influence the timing of these activities, photoperiod provides a more accurate cue and plays a large role in initiating or stopping seasonal behaviors. Photoperiod is so important in regulating behavior such as caching that researchers artificially manipulate the photoperiod for animals in captivity to induce this behavior (Pravosudov et al. 2010).
The day-night cycle also plays a key role in animal behavior. Some species are active during daylight, others at night, and still others only at dawn or dusk. Activity corresponds to diurnal variation in the availability of food sources, temperature requirements, and the presence or absence of major predators. Even without the cues of light and dark (e.g., in an all-light or all-dark environment), animals maintain a circadian rhythm approximately 24 h long (Roberts 1965), which suggests the existence of an internal circadian clock used to regulate daily activities.
Conditions also change over finer time scales, requiring another internal clock that works over seconds and minutes. Timing over the short term is particularly important for foraging. To forage efficiently, animals must be able to estimate time periods. This is particularly true for species that consume resources that refresh over time. For example, long-tailed hermit hummingbirds (Phaethornis superciliosus) forage on nectar, and birds must wait for flowers to refill before they return for another meal. Returning too soon would be a waste of time and energy for the hummingbird; waiting too long might mean losing the nectar to a competitor. So the birds learn to return within a few minutes of the time required for the flower to refill (Gill 1988). Experiments on timing in rats show that they can estimate short time intervals fairly precisely, but as the interval increases, their accuracy decreases (Gibbon 1977).
More or Less?
Many aspects of an animal's environment vary visibly in size and quantity. Peahens (Pavo spp.), for instance, use the number of eye spots on a male's tail as a cue when selecting a mate (Petrie & Halliday 1994). To use this cue, females must have some way to assess the quantity of eye spots. The ability to discriminate numbers is also important for group-living, territorial animals. For example, black howler monkeys (Alouatta pigra), which are highly territorial, can assess relative group size based on the number of males howling in a rival troop. This ability allows the monkeys to avoid potentially injurious encounters with larger troops (Kitchen 2006).
How accurate are animals at discriminating quantities? One rather accurate method would be to count the number of items. Hauser and colleagues (2000) tested free-ranging rhesus macaques (Macaca mulatta) to determine whether animals can precisely distinguish between small numbers. They placed apple slices into each of two boxes in full view of a monkey (Figure 5). They then allowed the monkey to choose a box from which to feed. When the number of slices put into a box was four or less, the monkeys accurately chose the box that contained more slices, but when the number of slices exceeded four for both options, they chose randomly.

Figure 5: Number discrimination.
Hauser et al. (2000) placed apple slices in boxes in full view of rhesus monkeys; the monkeys preferred boxes with more slices, provided the total number of slices in a box was less than four.
© 2012 Nature Education All rights reserved.
When the number of items is larger than three or four, the ability to distinguish precisely between amounts becomes more difficult. Yet in many situations, determining which of two options is "more" is important to an individual's fitness, so animals must use another mechanism to assess quantity. For example, many fish species group together in shoals; a larger shoal should provide greater benefits by decreasing predation risk. Agrillo and colleagues (2007) tested mosquitofish (Gambusia holbrooki) and found that the fish could distinguish between shoals that varied by a 1:2 ratio (1:2, 2:4, 4:8, and 8:16), but they were unable to discriminate a ratio of 2:3 (Figure 6). Many species show this effect of reduced precision as the ratio increases, and, like timing, animals are less accurate at quantifying as the magnitude increases (Brannon & Roitman 2003).

Figure 6: Number discrimination.
Fish chose larger shoals when one shoal was at least twice as large as the other; at smaller ratios, they were unable to determine which was the larger option (Agrillo et al. 2007).
© 2012 Nature Education All rights reserved.
Memory
Where did you put your keys? What is the capital of Germany? With whom did you speak at your last social gathering? How do you ride a bicycle? What was the first question asked here? Each of these questions represents a different type of memory, from short-term to long-term and including memory about places, facts, and experiences. Storing and retrieving information we have encountered previously can be useful when making predictions about the future. For animals as well, it is often beneficial to remember past information, and some animals seem to have enhanced memory for tasks that they face repeatedly in their natural environments.
Several species of birds cache seeds for the winter. For this to be an effective strategy, they must be able to remember the location of their caches months later, when they need the food. For example, captive black-capped chickadees (Parus atricapillus) are capable of recovering caches up to 28 d after caching (Hitchcock and Sherry 1990). Caching may have strong influences on at least two types of memory. First, caching species may have superior spatial memory. Clark's nutcrackers (Nucifraga columbiana) are corvids that cache up to 30,000 seeds each year (Vander Wall & Balda 1977). These seeds are an important winter food source for nutcrackers. Compared with non-caching corvid species, nutcrackers excel at remembering the locations of food when these species are tested in spatial memory tasks (Balda & Kamil 2006; Figure 7).
Caching may also impart an advantage to episodic memory. Episodic memory is the memory we use to recall experiences: the who, what, when, and where that we recall from specific episodes in our past. Though difficult to test in animals, there is evidence that some species have "episodic-like memory." Scrub jays (Aphelecoma californica), a relative of nutcrackers, also cache food (Figure 8). In experiments, they were allowed to cache nuts (a stable food source) and waxworms (a decaying food source). After both a 4-h and a 5-d delay, the jays were allowed to recover whichever food they wanted. Because they prefer waxworms, the jays retrieved more waxworms than nuts after the 4-h delay. After 5 d, however, the waxworms had decayed, so the jays retrieved more nuts than waxworms. The jays remembered what they cached, where, and when (either 4 h or 5 d ago) — these are the hallmarks of episodic-like memory (Clayton & Dickinson 1998).
Summary
Animals exhibit a range of fascinating cognitive abilities. Some species amaze us with their human-like abilities. Others amaze us because their abilities are so different from our own. Each species is unique in its constellation of cognitive capacities because evolution has tailored these capacities for solving the problems animals face in their physical and social worlds.
Glossary
caching: The act of storing food in a cache for later recovery
circadian: Daily; circadian rhythms refer to behaviors that occur at approximately the same time in a 24-h cycle
Corvid: A member of the family Corvidae, a group of birds that includes crows, jays, ravens, magpies, jackdaws and rooks
diurnal: Active primarily in the daytime
fitness: An individual's ability to pass on its genes; typically measured by the individual's lifetime reproductive success, fitness reflects both survival and reproduction
peahen: Female peafowl; counterpart of a peacock
photoperiod: Time period in a 24-h cycle during which organisms are exposed to light
shoal: A group of fish
spatial memory: Ability to recall the location of objects in space
References and Recommended Reading
Agrillo, C. et al. Quantity discrimination in female mosquitofish. Animal Cognition 10, 63-70 (2007).
Alerstam, T. et al. Migration along orthodromic sun compass routes by Arctic birds. Science 291, 300-303 (2001).
Balda, R. P. & Kamil, A. C. Linking life zones, life history traits, ecology, and spatial cognition in four allopatric Southwestern seed caching corvids. In Animal Spatial Cognition: Comparative, Neural, and Computational Approaches. M.F. Brown and R.G. Cook, eds. ([On-line]: 2006): Available at http://www.pigeon.psy.tufts.edu/asc/Balda/Default.htm
Brannon, E. A. & Roitman, J. D. Nonverbal representations of time and number in non-human animals and human infants. In Functional and Neural Mechanisms of Interval Timing. W.H. Meck, ed. (New York: CRC Press, 2003): 143-182.
Clayton, N. S. & Dickinson, A. Episodic-like memory during cache recovery by scrub jays. Nature 395, 272-274 (1998).
Dyer, F. C. The biology of the dance language. Annual Review of Entomology 47, 917-949 (2002).
Egevang, C. et al. Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proceedings of the National Academy of Sciences (USA) 107, 2078 -2081 (2010).
Gibbon, J. Scalar expectancy theory and Weber's law in animal timing. Psychological Review 84, 279-325 (1977).
Gill, F. B. Trapline foraging by hermit hummingbirds: competition for an undefended, renewable resource. Ecology 69, 1933-1942 (1988).
Hauser, M. D. et al. Spontaneous number representation in semi-free-ranging rhesus monkeys. Proceedings of the Royal Society of London, Series B 267, 829-833 (2000).
Hitchcock, C. L. & D. F. Sherry. Long-term memory for cache sites in the black-capped chickadee. Animal Behaviour 40, 701-712 (1990).
Kitchen, D. M. Experimental test of female black howler monkey (Alouatta pigra) responses to loud calls from potentially infanticidal males: effects of numeric odds, vulnerable offspring, and companion behavior. American Journal of Physical Anthropology 131, 73-83 (2006).
Lohmann, K. J. & Lohmann, C. A light-independent magnetic compass in the leatherback sea turtle. Biological Bulletin 185, 149-151 (1993).
Menzel, R. et al. Honey bees navigate according to a map-like spatial memory. Proceedings of the National Academy of Sciences (USA) 102, 3040 -3045 (2005).
Petrie, M. & Halliday, T. Experimental and natural changes in the peacock's (Pavo cristatus) train can affect mating success. Behavioral Ecology and Sociobiology 35, 213-217 (1994).
Pfungst, O. Clever Hans: (The Horse of Mr. Von Osten.) A Contribution to Experimental Animal and Human Psychology. (New York: Henry Holt, 1911).
Pravosudov, V. V., Roth, T. C. & LaDage, L. D. Chickadees are selfish group members when it comes to food caching. Animal Behaviour 80, 175-180 (2010).
Roberts, S. K. Photoreception and entrainment of cockroach activity rhythms. Science 148, 958 -959 (1965).
Tinbergen, N. The Study of Instinct. (Oxford: Clarendon Press, 1951).
Vander Wall, S. B. & Balda, R. P. Coadaptations of the Clark's Nutcracker and the pinon pine for efficient seed harvest and dispersal. Ecological Monographs 47, 89-111 (1977).
von Frisch, K. V. Decoding the language of the bee. Science 185, 663-668 (1974).
Wehner, R. Desert ant navigation: how miniature brains solve complex tasks. Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology 189, 579-588 (2003).
Recommended Reading
Brown, M. F. & Cook, R. G. Animal spatial cognition: comparative, neural, and computational approaches. ([On-line]: 2006): Available at http://www.pigeon.psy.tufts.edu/asc/toc.htm
Clayton, N. S. et al. The prospective cognition of food caching and recovery by western scrub-jays (Alphelocoma californica). Comparative Cognition and Behavior Reviews 1, 1-11 (2006).
Emmerton, J. Birds' judgments of
number and quantity. In Avian Visual Cognition. R.G.
Cook, ed. ([On-line]:
2001): Available at http://www.pigeon.psy.tufts.edu/avc/emmerton/
Meck, W. H. Functional and Neural Mechanisms of Interval Timing. (Boca Raton, Fla.: CRC Press, 2003).
Pearce, J. M. Animal Learning and Cognition: An Introduction. (New York: Psychology Press, 2008).
Shettleworth, S. J. Cognition, Evolution, and Behavior. (Oxford: Oxford University Press, 2010).
Wynne, C. D. L. Animal Cognition: The Mental Lives of Animals. (New York: Palgrave, 2001).










