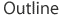

Selection is a powerful force in evolution, and it works in many ways. In the end, however, selection constantly sorts through the variation that is produced by mutations to select the fit and remove the unfit, while ignoring neutral changes. A few distinct types of selection are illustrated in Figure 1.
 and thus the trait values that will be positively selected. All other trait values are under negative selection.", "true", "All rights reserved.", '1682', '580', 'http://www.nature.com/nature_education');)
 Figure 1
Figure 1- Stabilizing selection keeps the population at one stable optimal value
- Directional selection transforms the value of a trait by increasing the frequency of individuals closer to a distant optimum
- Disruptive selection increases the frequency of large and small values of a trait at the expense of intermediate values
- Balancing selection selects the optimal compromise among several constraints
At their core, all forms of selection involve individuals with inherited differences in fitness competing within the same population. This competition is about fitness as measured by survival rates, fecundity, or some other trait that correlates with fitness. The "winner" of the competition is positively selected, and its genotype increases in frequency; on the other hand, the "loser" is negatively selected, and the frequency of its genotype decreases. Thus, negative selection and positive selection cannot be separated. To make communication easier, however, scientists talk about positive selection when the focus of a particular study is on an increase in rare variants that improve optimal fitness, and they speak of negative selection when the focus is on the removal of harmful variants.
Causes of Negative Selection
;)
© 2004 The Royal Society Hall, J. P. W., Robbins, R. K. & Harvey, D. J. Extinction and biogeography in the Caribbean: new evidence from a fossil riodinid butterfly in Dominican amber. Proceedings of the Royal Society B 271, 797–801 (2004). All rights reserved. 
More short-term negative selection is also widespread, especially due to ecological reasons. Many structures in biology are only conditionally optimal, because they depend upon the details of other structures or circumstances to perform their function. If such other structures are within the same organism, this relationship is termed epistasis. For example, two proteins could interact epistatically in such a way that a deleterious mutation in one protein could be either compensated for or aggravated by a mutation in the other protein (Burch & Chao, 1999). Frequently, ecological circumstances also play a role in determining mutational effects. For instance, if the niche of a species stays the same, some mutations that would be beneficial in other niches will be under negative selection. If the niche changes, however, some traits that were previously under negative selection may suddenly be beneficial and have a greater fitness than the majority of the previously favored genotypes.
If environmental interactions include other rapidly evolving species, then the pressure to change may never stop, and the evolutionary optimum will always remain some steps ahead. Host-parasite interactions are a famous example of this sort of situation. Here, the host immune system evolves to recognize a special structure on the parasite and allow its removal. This in turn induces negative selection on the current form of the parasite while leading to positive selection of variants that cannot be recognized by the host. Furthermore, if such variant parasites exist, they will increase in frequency and in turn induce negative selection of the current variant of the host, which will lead to the positive selection of hosts that can again recognize the parasite, and so on. Negative (and positive) selection in such a system never rests, which is why one hypothesis describing these systems was named after the Red Queen in the book Alice in Wonderland, who famously stated that it takes a great amount of running to stay in the same place.