
Abstract
We propose that postmortem tissue is an underutilized substrate that may be used to translate genetic and/or preclinical studies, particularly for neuropsychiatric illnesses with complex etiologies. Postmortem brain tissues from subjects with schizophrenia have been extensively studied, and thus serve as a useful vehicle for illustrating the challenges associated with this biological substrate. Schizophrenia is likely caused by a combination of genetic risk and environmental factors that combine to create a disease phenotype that is typically not apparent until late adolescence. The complexity of this illness creates challenges for hypothesis testing aimed at understanding the pathophysiology of the illness, as postmortem brain tissues collected from individuals with schizophrenia reflect neuroplastic changes from a lifetime of severe mental illness, as well as treatment with antipsychotic medications. While there are significant challenges with studying postmortem brain, such as the postmortem interval, it confers a translational element that is difficult to recapitulate in animal models. On the other hand, data derived from animal models typically provide specific mechanistic and behavioral measures that cannot be generated using human subjects. Convergence of these two approaches has led to important insights for understanding molecular deficits and their causes in this illness. In this review, we discuss the problem of schizophrenia, review the common challenges related to postmortem studies, discuss the application of biochemical approaches to this substrate, and present examples of postmortem schizophrenia studies that illustrate the role of the postmortem approach for generating important new leads for understanding the pathophysiology of severe mental illness.
Similar content being viewed by others
INTRODUCTION: THE PROBLEM OF SCHIZOPHRENIA
Schizophrenia affects nearly every person in the world in some way. Based on the lifetime prevalence of the illness, ∼1 in 100 persons has schizophrenia (Bhugra, 2005). Most people know someone or have a family member, however distant, with this diagnosis. The direct costs of this illness are high (Wu et al, 2005). More hospital beds are occupied by persons with schizophrenia than all other psychiatric illnesses combined (Buchanan and Carpenter, 2000). Most staggering are the personal costs: lost opportunities for employment, long-term disability, social isolation, frequent bouts of severe psychopathology, frequent hospitalizations, and a high rate of suicide completion (Knapp et al, 2004).
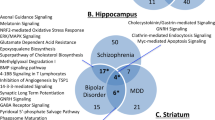
While there are well-established criteria in place for making the diagnosis of schizophrenia, the causes are still unknown. Most recent evidence supports a combination of genetic and environmental factors contributing to the development of the disorder (Marenco and Weinberger, 2000). To date, no one gene, single-nucleotide polymorphism, or mutation has been consistently linked to the illness, and it is likely that multiple susceptibility genes create a predisposition to developing schizophrenia (Fanous et al, 2012; Gershon et al, 2011; Levinson et al, 2012). Genetics studies, which have led the way for diseases such as breast cancer and cystic fibrosis, have been much less impactful in the schizophrenia field. Perhaps the strongest genetic data come from the DISC translocation study (Millar et al, 2000). This Scottish cohort with a high penetrance for schizophrenia has a balanced translocation (t (1;11)); 7/29 carriers in this family have schizophrenia, suggesting that the disrupted genes at these loci may be important in the development of this illness (Blackwood et al, 2001). The authors named the primary disrupted gene ‘Disrupted In Schizophrenia’ or DISC. However, 10/29 of the subjects have a severe mood disorder, suggesting that the disrupted genes have a role in severe mental illness, encompassing several possible phenotypes for the genetic risk conferred by these mutations (Chubb et al, 2008).
The most likely role for genetics in schizophrenia is a polygenic one. A recent genome-wide association study (GWAS) of ∼2500 subjects with ∼700 000 single-nucleotide polymorphisms (SNPs) looked at the relationship between genetic markers and three dimensions, positive, negative/disorganized, and mood, and concluded that schizophrenia is polygenic (Fanous et al, 2012). Another genome-wide analysis identified four SNPs across five disorders in >30 000 patients and ∼30 000 controls, which reached genome-wide significance, suggesting that severe mental illnesses such as schizophrenia, bipolar disorder, and depression share common polygenic risk factors (Smoller et al, 2013). These studies suggest that schizophrenia is a polygenic syndrome.
The onset of schizophrenia can be cruel; unremarkable development until the late teens or early twenties, followed by profound episodes of psychiatric illness, typically including severe thought disorder symptoms (Alda et al, 1996). Paranoid delusions, auditory hallucinations, and cognitive impairment including working memory and decision-making deficits may characterize the initial onset of this illness (Badcock, 2010; Kay, 1990). The psychotic symptoms often respond to high levels of dopamine receptor type 2 (D2) blockade; preterminal and/or presynaptic dopamine receptors modulate glutamate and gamma-aminobutyric acid (GABA) neurotransmission, and it is through these other neurotransmitters that D2 receptor antagonists likely have their effects (Lewis et al, 2004). Unfortunately, only ∼70–80% of patients respond to these drugs, and they may incompletely improve the positive (that is, psychotic) symptoms of the illness (Joyce and Meador-Woodruff, 1997; Laruelle et al, 1999). One of the best pharmacological treatments, clozapine, is associated with several compliance barriers including frequent blood draws and significant adverse effects. The Clinical Antipsychotic Trials of Intervention Effectiveness (CATIE) trial found that ∼75% of all patients are non-compliant with treatment, with either the older typical medications or the newer second generation medications (Lieberman et al, 2005; Meyer et al, 2005). These studies highlight the need for new treatment approaches, not just for the positive symptoms but also for the negative and cognitive symptoms.
One approach to find new drug targets involves identifying pathophysiological substrates of this illness (Figure 1). Synthesizing and injecting insulin to treat diabetes would not be possible without understanding what insulin is, where it comes from, and how it works. Unlocking the pathophysiology of schizophrenia is a seemingly more difficult task: (1) the illness is syndromic, heterogeneous, and has multiple genetic and environmental risk factors; 2) animal models may not fully recapitulate this arguably human condition; for example, locomotor hyperactivity in rodents is often used as a proxy for psychosis, but it does not capture the full complexity of auditory hallucinations or delusions in humans; (3) there are likely many different pathological mechanisms that can result in the schizophrenia phenotype. There are >20 different animal models that exhibit various schizophrenia-related endophenotypes such as abnormal prepulse inhibition, working memory deficits, poor grooming, and locomotor hyperactivity (Table 1); (4) diverse (but connected) parts of the brain are implicated, and current pharmacological treatments bind expressed receptors without cellular or regional specificity. For example, D2 blockade in the striatum may be effective to treat some symptoms, but blocking dopamine receptors in the cortex may impair cognition. The field needs to identify new substrates for drug discovery that come from a deeper understanding of the pathophysiological processes present in the central nervous system (CNS) in schizophrenia.

A role for postmortem studies for investigating severe mental illness. Schizophrenia is caused by a combination of genetic risk and environmental factors that combine to create a disease phenotype that is typically not apparent until late adolescence. The complexity of this illness and its etiology creates challenges for hypothesis testing aimed at understanding the pathophysiology of the illness as well as developing new and more effective treatments. Postmortem brain tissues collected from individuals with schizophrenia reflect neuroplastic changes from a lifetime of severe mental illness (and the life stressors associated with severe medical illness) as well as treatment with antipsychotic medications. While there are significant concerns with studying postmortem brain, such as the postmortem interval, it confers a translational element that is difficult to recapitulate in animal models. Data derived from animal models typically provides specific mechanistic and behavioral measures that cannot be generated in the human condition. Convergence of these two approaches is providing important insights for understanding the molecular deficits and their causes in this often devastating illness.
Given the challenges of studying this and other severe mental illnesses, we propose that postmortem tissue is an underutilized substrate that may be used to translate genetic and/or preclinical studies. In this review, we first discuss the challenges of using the postmortem brain substrate. In the next part we review protein biochemistry approaches and the special challenges of using these methods with postmortem brain. In next section, we review examples of postmortem studies that were driven by findings from clinical genetics studies, were inspired by data from preclinical studies, highlight convergent data from complimentary biochemical techniques, and inform and refine the N-methyl D-aspartate subtype glutamate (NMDA) receptor hypothesis of schizophrenia. In the final section we summarize the role of postmortem brain in the study of severe mental illness and present novel future directions the field may profitably pursue.
THE POSTMORTEM SUBSTRATE: USING BRAIN TISSUES FROM SUBJECTS WITH SCHIZOPHRENIA
The collection of samples and the construction of postmortem brain tissue cohorts have traditionally matched subjects by age, sex, postmortem interval (PMI), and tissue pH. Earlier studies (with some notable exceptions) were primarily focused on measuring transcript expression, and relied on these factors, along with the more recently developed and now widely used mRNA integrity (RIN) value, to determine the suitability of the sample for inclusion in a postmortem study (Fleige and Pfaffl, 2006; Webster, 2006). Other variables, including manner of death, diagnostic certainty, co-morbid substance use, and prior medication treatment status may influence (or confound) dependent measures in postmortem tissue and are considered important factors for matching or exclusion. Despite accounting for these factors, postmortem studies may find inconsistent results for the same dependent variable. For example, expression of mRNA for the GluN1 subunit of the NMDA glutamate receptor in the frontal cortex in schizophrenia was decreased (Sokolov, 1998), increased (Dracheva et al, 2001; Le Corre et al, 2000), and unchanged (Akbarian et al, 1996) in separate studies using tissues from different brain collections. Such disparate findings may be attributed to differences in methodology, variation in dissection protocols, and differences in the biological substrate due to the heterogeneity of the illness being studied. In the remainder of this section we examine each of these factors and discuss how they are generally handled by the field.
Age at Death
Age-related changes have been reported for several gene expression measures (Funk et al, 2012). While protein expression does not always correspond to changes in mRNA levels, the BrainCloud project has demonstrated the complexity of age-related changes in global transcript expression across the lifespan (Colantuoni et al, 2011). It is not surprising that the largest changes in gene expression occur during development and later near end of life, highlighting the need for well-matched postmortem cohorts, as well as having a broad range of ages within a study when possible. For example, divergent changes in vesicular glutamate transporter (VGLUT1) mRNA expression were reported in the hippocampus in schizophrenia. Decreased VLGUT1 mRNA was reported in a middle-aged cohort, whereas no changes in the transcript were found in an elderly cohort (Eastwood and Harrison, 2005; Uezato et al, 2009). The authors of one of the studies attributed this effect to the difference in the ages of the subjects included in the two studies (Eastwood and Harrison, 2005; Uezato et al, 2009). These findings, as well as the conclusions of the BrainCloud project, are consistent with reports of gene expression-specific associations with age of death (Eastwood and Harrison, 2005; Huerta et al, 2006; McCullumsmith et al, 2007; Oni-Orisan et al, 2008). Age-related changes are likely not limited to transcripts. While there are fewer studies demonstrating age-related changes in protein-related measures in postmortem brain, Gα-proteins binding and stimulation, guanylyl cyclase protein levels, and choline acetyltransferase (CAT) activity all decreased with increasing age (Breese et al, 1995; Eastwood and Harrison, 2001; Gonzalez-Maeso et al, 2002; Ibarra et al, 2001; Perry et al, 1977; Spokes et al, 1979). In summary, subjects should be carefully matched for age in postmortem studies, and the effect of age on dependent measures carefully examined.
Manner of Death
Manner of death may impact the integrity of the tissue obtained at autopsy and it may also reflect underlying differences in pathophysiology. For example, specific changes in mRNA and protein levels of nerve growth factor (NGF), neurotrophin (NT)-3 and NT-4/5 were associated with death by suicide but not by natural causes or accident (Dwivedi et al, 2005). The direct physiological effects of the specific manner of death can impact tissue pH, and may affect dependent measures in divergent ways. The phrase ‘agonal status’ derives from the medical term for shallow, infrequent breaths, called agonal respirations. Agonal respirations often follow cardiogenic shock (Rea, 2005), and may lead to a prolonged period of hypoxia due to agonal breathing at the end of life. Several studies have reported diminished mRNA quality associated with prolonged agonal states (Barton et al, 1993; Durrenberger et al, 2010; Harrison et al, 1991, 1995; Kingsbury et al, 1995; Lipska et al, 2006a, 2006b; Tomita et al, 2004a, 2004b; Vawter et al, 2006; Weickert et al, 2010). While subjects known to have a prolonged agonal status are typically excluded from postmortem studies, the effects of agonal status on proteins are not as well established. Low brain pH is associated with prolonged agonal states, and subjects with low tissue pH (<6.0) are typically not included in postmortem studies (Durrenberger et al, 2010; Ervin et al, 2007; Hardy et al, 1985; Harrison et al, 1995; Kingsbury et al, 1995; Tomita et al, 2004a, 2004b). However, changes in brain tissue pH secondary to prolonged agonal status were associated with altered electrophoretic mobility, diminished neurotransmitter-related enzymatic reactions, as well as changes in gene expression for molecules involved in mitochondria respiration, stress response, and proteolytic activities (Butterworth et al, 1985; Cain and Kappy, 1971; Czudek and Reynolds, 1990; Kingsbury et al, 1995; Li et al, 2004; Perry et al, 1977; Spokes et al, 1979). While manner of death impacts measures of gene expression, most brain collections do not include subjects with prolonged agonal status, making this issue more of a theoretical concern.
Diagnosis and Phenotype
Brain collection protocols vary, with some brain collections utilizing prospective characterization of subjects, while others determine diagnosis retrospectively, based on medical records and family interviews. In the case of prospective assessment, there may be a long delay (⩾20 years) between assessment and death. For some illnesses, diagnostic criteria may change as a field evolves, and the clinical profiles of the subjects may change over time. There may also be difficulties in tracking records and participants of prospective donor programs, making the yield for collection very low. For some subjects antemortem assessment is not feasible, as in the case of suicide completers.
The heuristic value of diagnostic criteria may be a limiting factor for the construction of postmortem brain cohorts, because syndromes such as major depressive disorder or schizophrenia can be phenotypically heterogeneous. The identification of behavioral endophenotypes, which bridge the gap between genetic risk factors and symptom presentation, might help reduce the variability in postmortem samples due to the descriptive nature of diagnostic criteria for mental illnesses. Useful endophenotypes might include rapid eye movement (REM) sleep abnormalities or prepulse inhibition deficits, which are present in depression and schizophrenia, respectively (Hasler et al, 2004; Hazlett et al, 2007). Finally, the effects of a lifetime of severe mental illness, and the stress associated with it, may not be disease-specific. Inclusion of a comparison group with a different mental illness permits assessment of the specificity of the findings to a particular disorder (Dorph-Petersen et al, 2009; McCullumsmith and Meador-Woodruff, 2002). In summary, postmortem studies are reliant on descriptive criteria for diagnostic assessment for diseases that are typically quite heterogeneous in their clinical presentation. As the field improves its understanding of severe mental illness, improvement in diagnostic specificity will aid in the construction of well-matched postmortem brain disease cohorts.
Dissection Protocols and Laterality
Divergent results of the same dependent measures in studies using different postmortem brain cohorts may be due to variability in tissue processing and/or dissection protocols. There may be marked variability in gene expression and function depending on the precise boundaries of the sample (Fanselow and Dong, 2010). Assessment of the effects of laterality is often unfeasible in postmortem studies, as most collections typically save half the brain by freezing and half by chemical fixation (Bruder, 1983; Lepage et al, 2006; Webster, 2006; Wible et al, 2009). Laterality may be an underappreciated source of variance in postmortem studies.
Freezer Storage Time
Another factor that may impact postmortem samples is freezer storage time. Postmortem samples are typically collected over months to years due to the relatively slow rate of tissue availability. The effects of freezer storage time on dependent measures have been examined. Storage time did not affect mRNA quality or quantity (Burnet et al, 1997; Harrison et al, 1995; Stockmeier, 1997; Weickert et al, 2010). However, extended freezer storage time affected oligo dT priming of reverse transcription, suggesting that there may be degradation of the polyA tail (Johnston et al, 1997; Leonard et al, 1993). The effects of freezer storage time on protein measures have also been assessed. Storage time for tissue at −70 °C over a period of 1–85 months did not significantly affect immunoreactivity of Gα-proteins (Gonzalez-Maeso et al, 2002). Protocols for tissue processing may also impact protein degradation. Samples held at 1 °C for five different time points (5–50 hours postmortem delay) prior to −80 °C storage had minimal changes in protein expression as measured by western blot (Ferrer et al, 2007). However, left frontal lobe protein levels of α-synuclein, β-synuclein, vacuolar proton ATPase, peroxiredoxin, ATP synthase, fructose-biphosphate aldolase C, superoxide dismutase I, amphiphysin, and α-enolase were reduced when samples were first stored at 4 °C or room temperature for 5–50 hours and then frozen (Ferrer et al, 2007). When possible, freezer storage time should be probed to determine whether there is an impact on the dependent measures in postmortem studies.
Sample Size and Study Design
Postmortem cohorts have increased in size due to increased collection rates of brain samples, making well-matched samples more readily available (Webster, 2006). This has led to studies having more subjects and better matching for tissue pH, postmortem interval, age, and sex (Webster, 2006). Another approach is to utilize dyads or triads of matched subjects, increasing the power to detect an effect (Glantz and Lewis, 1997; Pierri et al, 2001). The field is shifting away from studies with <10 subjects per group, and the increased sample sizes accord increased statistical power to detect significant changes with smaller effect sizes (Carter et al, 2007). However, the use of large subject groups may not be feasible for all approaches. For example, for ultrastructural studies using electron microscopy, where sample size is limited both by the availability of high quality (low PMI) samples and the nature of the technique being labor-intensive, studying fewer brains in more depth is a worthy approach.
Prior Medication Exposure
The treatment of severe mental illness typically includes long-term administration of antipsychotic medications. As discussed in the introduction, these medications powerfully impact brain neurochemistry. A large number of studies have found changes in gene expression following treatment with antidepressants, antipsychotics, mood stabilizers, and other psychotropic medications (Beerpoot et al, 1996; Eggan et al, 2008; Freyberg et al, 2010; Oni-Orisan et al, 2008). Medication effects have been assessed in several ways. Rodent animal models are typically used for these studies and the medication administered for up to 4 weeks (Bennett et al, 2007; Delfs et al, 1995; Toyooka et al, 2002a, 2002b; Uezato et al, 2009). As such a short duration of treatment does not model a lifetime of treatment, longer treatment intervals (such as 6 months) would be more analogous to the treatment course in human subjects (Tamminga et al, 1990; Tamminga et al, 1994). The most analogous treatment models utilize nonhuman primates, but are limited from widespread use by access to primate colonies as well as cost (Akil et al, 2000; Eggan, 2008; Konopaske et al, 2008).
Another approach is based on the availability of subjects ‘off’ medications for a period of time prior to death. This permits secondary analyses of the effects of these medications, an approach limited by the notion that being off antipsychotic medications for weeks to months may not reverse a lifetime of neuroplastic changes (Haroutunian et al, 2006; Oni-Orisan et al, 2008). A third way to probe for medication effects is to perform correlation analyses between the dependent measure and a measure of total lifetime exposure to the drug, typically expressed as chlorpromazine equivalents (Law et al, 2004). This approach has shortcomings as well, including the high rate of treatment noncompliance in patients with severe mental illness (Basil et al, 2006). While antipsychotic treatment is a potential confound, particularly for postmortem schizophrenia studies, changes attributable to antipsychotic treatment are interesting in their own right, and alterations found in postmortem brain are perhaps best thought of occurring as a result of a lifetime of mental illness and antipsychotic treatment.
Postmortem Interval (PMI)
PMI is defined as the time from death to extraction and freezing of the brain tissues at −80 °C. While the effects of PMI on global mRNA integrity are minimal, there may be transcript-specific degradation (Bahn et al, 2001; Barton et al, 1993; Castensson et al, 2000; Ervin et al, 2007; Gilmore et al, 1993; Harrison et al, 1995; Leonard et al, 1993; Oni-Orisan et al, 2008; Pardue et al, 1994; Stan et al, 2006; Trotter et al, 2002). Several studies have specifically examined the effects of PMI on protein measures in the brain. Fountoulakis et al (2001) assessed the effects of PMI in rodent brain over 0–48 hour intervals using 2D electrophoresis and matrix-assisted laser desorption ionization mass spectrometry. The authors determined that changes in levels of some proteins were related to PMI. They propose that proteins that are sensitive to PMI could be excluded from postmortem studies and conclude that most changes occurred ∼24 hours or later. Notably, dihydropyrimidinase-related protein-2, neurofilament, alpha-internexin, synaptosomal-associated protein 25, GFAP, heat shock proteins, and dynamin-1, were decreased at 48 hours PMI. Truncated forms of several other proteins were increased (Fountoulakis et al, 2001).
PMI effects on protein expression may be region-specific. The substantia nigra was found to be exceedingly vulnerable to increased protein oxidation/nitration events with increasing PMI, with increased oxidation/nitration of GFAP and neurofilament protein, while the frontal cortex, cerebellum, and caudate nucleus were relatively preserved (Chandana et al, 2009). The authors posit that the inner regions of the brain may cool more slowly than the outer, accounting for some of their results. Another study extensively examined the effects of PMI on expression of glutamate transporters in the rodent brain (Beckstrøm et al, 1999). They found epitope-specific degradation of rodent glutamate transporters 1 and 2 at 24 hours PMI, compared with 0 and 12 hours PMI. Differential degradation of different epitopes was observed after 24 hours, with selective preservation of the C termini for rodent glutamate transporters 2 and 3 at 72 hours. These effects extended to both immunoreactivity as well as glutamate transport activity (Beckstrøm et al, 1999). The data in this section support the current approach in the field to match samples as closely as possible for PMI and to probe for associations between PMI and each dependent measure.
Sample Quantity and Quality
A final consideration for this section involves sample abundance. There is only so much dorsomedial nucleus of the thalamus; getting more of this region involves more than ordering or breeding more rodents. In addition, some approaches, such as chromatin-immunoprecipitation (ChIP), require large quantities of tissues from each subject (Huang et al, 2006). One approach to get around this issue has been pooling of samples; pooled samples are assayed and help generate specific hypotheses that may be tested with smaller quantities of tissue (English et al, 2009; English et al, 2011; Pennington et al, 2008). One area of postmortem proteomic research that lags behind transcript studies is the assessment of protein quality. PMI, pH, and age are typically the potential covariates that are probed for effects on dependent measures, but there is currently no consensus on what constitutes a high-quality protein sample. Confusing the matter further, one study found a positive correlation between protein immunoreactivity and PMI, while another found increases in ubiquitin-immunoreactivity in relation to the length and intensity of agonal status (Piette et al, 2011). These data suggest that translation of some proteins in dying cells may be selectively upregulated, or that during the postmortem interval changes in protein folding or conformation alter antigenicity of the target epitope, possibly secondary to a decrease in tissue pH (Yoo et al, 1999). Most postmortem studies measuring protein levels assess the impact of age on their dependent measures, and several studies have found correlations between age and levels of some proteins (Eastwood and Harrison, 2005; Oni-Orisan et al, 2008). Additional studies are needed to probe for the effects of factors such as tissue pH, PMI, and freezer storage time. As protein studies become more sophisticated, global oxidation states or other potential measures of protein integrity may be developed as indices of protein quality.
BIOCHEMICAL MEASURES OF PROTEINS IN POSTMORTEM TISSUES
Omics-based approaches have revolutionized the study of human disease states, and led to important new leads for understanding the pathophysiology of severe mental illnesses. However, there is an ongoing role for tried and true techniques, including antibody-based approaches, for the testing of specific hypothesis suggested by genetics and/or animal models, as well as to translate findings generated by novel ‘omics’ approaches. For example, a rare missense variant in the neuregulin 1 gene was associated with schizophrenia in subjects living in the central valley of Costa Rica, leading to a series of predictions for the function of this variant based on its location in the transmembrane domain of the protein (Walss-Bass et al, 2006). In vitro, this variant was found to inhibit dendrite formation, associate with altered pro-inflammatory cytokine expression, and prevent gamma secretase-mediated processing of neuregulin 1 (Chen et al, 2010; Dejaegere et al, 2008; Marballi et al, 2010). Subsequent western blot studies demonstrated abnormalities of protein expression (including gamma secretase subunit PS1) associated with perturbed neuregulin 1 processing in postmortem brain in schizophrenia that were brain region-specific (Marballi et al, 2012). These data highlight the ongoing utility of the western blot approach in postmortem tissues. In this section we discuss biochemical approaches and the special challenges of using these methods with postmortem brain, using examples from the schizophrenia literature.
Western Blot Analyses
The wide availability of commercial primary antibodies, as well as the relatively lower costs of making custom antibodies, has led to a proliferation of antibody-based studies using postmortem tissue. The lack of specificity of some antibodies, as well as the narrow range of detection for western blot assays, is a potentially significant limitation to this technique. Western blot studies typically utilize standard gel electrophoresis approaches, requiring multiple gels to run a sample set. Between-blot variability in increasingly larger study cohorts may introduce unanticipated variability. For example, running, transblotting and probing two to three gels or membranes is straightforward, but scaling up for 5–10 times this number may require various normalization techniques. Normalization to a housekeeping gene or loading a calibrator sample onto each gel may control for some of these issues. Housekeeping genes should be carefully evaluated themselves for changes in expression in various psychiatric diagnoses (Bauer et al, 2009; Stan et al, 2006).
The next generation of imaging devices diminished some of the problems associated with earlier film-based quantification methodologies, but introduced new challenges. For example, greater sensitivity may lead to visualization of bands not seen with older, less sensitive methods. These ‘extra’ bands may represent different protein isoforms or low-level nonspecific cross-reactivity that was not previously detectable. There are several ways to address the specificity of the western blot assay, other than relative migration. The gold standard would be to use tissues from a knockout animal that does not express the protein, and demonstrate a band at the predicted relative migration in the wild-type tissues (Holmseth et al, 2012). For proteins lacking regions of sequence similarity between the rodent and human proteins, knockout studies may not be useful. Preadsorption studies may also be performed with a blocking peptide to show that the extra bands are nonspecific.
Film-based detection of proteins in western blot protocols are diminishing in use, and are generally no longer considered state-of-the-art. Film-based detection methods may easily result in oversaturated images, leading to a ceiling effect and a loss of the ability to detect small changes in protein expression. Newer infrared devices are film-free (and thus cost-less to operate), difficult to saturate, and typically more sensitive than older film-based methods.
Recent work has found cell-specific changes in transcript expression in schizophrenia using laser-capture microdissection (LCM)(Ruzicka et al, 2007; Sodhi et al, 2011). These findings suggest that there are cell type-specific abnormalities of protein expression in this illness, and there are numerous hypotheses that could be tested if the LCM technique could be coupled with a reliable, high-sensitivity protein assay. Based on current western blot technology, it would take a laser-capture of ∼10 000 cells to generate one western blot sample (unpublished observation).
There are several promising methods that have recently been developed that may provide a way to make nanoscale measurement of proteins. Microcapillary electrophoresis facilitates antibody-based measurement of small volume, low concentration samples, as low as 25 cultured cells in 400 nl of buffer (O'Neill et al, 2006). We have piloted this technique in our laboratory, and we found that we can measure moderately abundant proteins at a concentration of 0.2 μg/μl in a sample volume of 10 μl (Figure 2). This would permit ∼30 replicates or runs of an LCM-acquired sample with 1000 captured cells. One limitation of the microcapillary devices on the market is that there are a limited number of commercially available assays for specific proteins, and development of new assays is costly and requires access to the device, which can run into the low six figures. One of the strengths of the newer microcapillary devices is that they can separate based on size or charge, permitting simultaneous interrogation of protein isoforms that are differentially phosphorylated.

Laser-capture microdissection coupled to protein assays. Nanoscale microcapillary electrophoresis (using the Nanopro1000 device, Protein Simple Corp., Santa Clara, CA, USA) analysis of (a) HeLa cell lysates, (b) 600 laser-capture microdissected (LCM) pyramidal neurons from the frontal cortex, or (c) cortical brain homogenate. The HeLa lysate (positive control) expresses all isoforms of ERK detectible by the NP1000 pan-ERK assay. The coefficient of variability for ERK2 in the pyramidal neuron LCM sample (n=6 replicates) was 8.1%.
These microcapillary devices could be made less relevant if protein PCR assays become widely used. This recently developed tool combines an antibody-based approach with QPCR (Fredriksson et al, 2002; Gullberg et al, 2004). Using an oligonucleotide-labeled polyclonal antibody (or two different monoclonal antibodies specific for different epitopes on the same protein), one batch of ‘left primer’ antibody and one batch of ‘right primer’ antibody are biotinylated and coupled to specific ‘left’ and ‘right’ oligonucleotides; when the left and right antibodies are colocalized on the target protein, the left and right oligos may then hybridize with a linking oligo and be amplified using PCR. While not yet in widespread use, this technique is potentially a promising low-cost way to perform multiple nanoscale measurements of specific proteins in small volume and/or low concentration samples such as those generated by LCM.
Assessment of Protein–Protein Interactions in Postmortem Tissues
Studies employing the immunoprecipitation of proteins are no longer just ‘precipitating’ their target of interest; rather, one of several capture systems is employed, although specificity is still dependent on the quality of the primary antibody. Thus, we propose ‘immunoisolation (II)’ as a more accurate term to describe this technique. Popular capture media include protein A, protein G, or protein A/G-linked agarose or magnetic beads, with attachment of the primary antibody to the bead substrate (Kristiansen et al, 2010). The antibody–bead complex is incubated with the sample, and then the beads are isolated. In the case of the agarose beads, isolation is performed by centrifugation. In the case of the magnetic beads, a magnet is used to immobilize the bead–antibody complex, while the supernatant is exchanged or removed.
Coupled with western blots, these approaches have been used to analyze protein–protein interactions in postmortem samples from subjects with schizophrenia. For example, synapse-associated protein 25 (SNAP25) and mammalian uncoordinated-18 (MUNC18) protein levels were decreased following immunoisolation of phosphosyntaxin 1 from the frontal cortex in schizophrenia (Castillo et al, 2010). Another group used a modified enzyme-linked immunosorbent assay (ELISA) to assess the interaction of syntaxin with SNAP25 in the striatum using a SNAP25 capture antibody. Interestingly, there was an increase in SNAP25-associated syntaxin protein levels, whereas total syntaxin and SNAP25 protein levels were decreased in schizophrenia, suggesting that direct measurements of gene expression may fail to detect some types of protein abnormalities (Barakauskas et al, 2010). Another study found higher levels of a protein homolog that co-islolated with presenilin 2 from frontal cortex tissue homogenates, whereas decreased dopamine transporter protein levels were found in samples immunoisolated using anti-D2 dopamine receptor antibodies (Lee et al, 2009; Smith et al, 2004). These data demonstrate the need for more nuanced biochemical studies that move beyond straightforward measurements of gene products in tissue homogenates.
Finally, immunoisolation has been used to capture subcellular organelles from postmortem brain tissue, including endoplasmic reticulum (ER) and endosomes. We used antibodies that facilitated selective capture of early and late endosomes, and analyzed the endosomal contents using western blot analyses. We found increased GluA1 levels in early endosomes from frontal cortex, suggesting an increased rate of turnover for GluA1-containing receptor complexes (Hammond et al, 2010, 2011). In a different study, we used an anti-calnexin antibody to capture ER microsomes from postmortem brain homogenate. We found decreased expression of PSD95 and GluN2B in the ER-enriched fraction (but no change in total levels), suggesting increased forward trafficking of NMDA receptor complexes from the ER (Kristiansen et al, 2010).
Limitations of Bead-Based Capture Systems
We have tested several different capture systems and found significant nonspecific binding of the targeted substrate, including the postsynaptic density, to the capture media (McCullumsmith and Meador-Woodruff, 2011). To overcome these limitations, we have devised several strategies including preclearing samples (Hammond et al, 2010; McCullumsmith and Meador-Woodruff, 2011). Beads used with samples that are not precleared nonspecifically bind proteins from several subcellular compartments including ER, mitochondria, and postsynaptic densities (McCullumsmith and Meador-Woodruff, 2011). Thus, studies employing these methods require rigorous characterization of the immunoisolation preparation, as well as positive and negative controls for each component of the assay, including primary and secondary antibodies, capture media, and detection systems. Without proper controls, dependent measures analyzed using antibody–bead capture systems are difficult to interpret.
Another limitation of using immunoisolation to study the proteome is the association of the target molecules and their binding partners during the postmortem interval and/or tissue processing. For example, if a protein is typically sequestered in the postsynaptic density (PSD), homogenization could increase the interactions between this protein and interacting partners not yet trafficked to the PSD. This theoretical concern raises the issue of whether it is desirable to analyze samples that have been ‘fixed’ at the moment of death. One way to address this issue is using rodent tissues that simulate the postmortem interval, by decapitating the rodents, and harvesting the brain tissues over a range of time intervals to simulate the delays implicit to human brain collection.
In summary, immunoisolation is not being widely used, in part due to the technical challenges associated with this approach. The few studies that have employed this method using postmortem tissues have found abnormalities of protein–protein interactions that most likely reflect altered protein trafficking in schizophrenia.
Biochemical Fractionation of Postmortem Tissues
Biochemical fractionation of fresh tissues or cell lysates is a well-established method to assess various dependent measures in biological samples. However, as postmortem tissue is typically frozen (−80 °C), there may be breakdown of membrane integrity and cross contamination between subcellular compartments. Thus, the need for appropriate control experiments is critical in these types of studies (Bubber et al, 2004; Tsai et al, 1995). Fractionation can often lead to increased protein assay sensitivity due to enrichment of targeted proteins, release from a subcellular organelle, or dissociation from a protein complex. A number of studies have used a relatively straightforward approach by simply purifying membrane and cytosolic fractions, where the membrane fraction contains all the organelles, including nuclei, ER, mitochondria, and plasma membranes (Dean et al, 2008; Hahn et al, 2006). A newer direction for postmortem studies involves a more comprehensive isolation of subcellular fractions. Such studies are typically hypothesis-driven, with focus on a particular subcellular fraction. For example, we optimized a biochemical fractionation protocol to yield a fraction enriched for ER that was free from nuclear, PSD, and golgi markers. We confirmed the purity of this fraction using electron microscopy, as well as western blot analysis using specific markers of each of these fractions (Hammond et al, 2012).
Biochemical Enrichment of Postsynaptic Densities
The PSD is a subcellular microdomain with a number of proteins that contain binding domains that promote protein–protein interactions. Generating a highly purified, enriched PSD fraction from postmortem tissue has been challenging. The traditional approach to isolating the PSD involves using a density gradient to generate synaptosomes. Synaptosomes are small lipid vesicles that form from synapses and astrocytic process following mechanical disruption of brain tissue using a blender or homogenizer (Whittaker et al, 1963). Synaptosomes may contain pre- and postsynaptic elements; this is a potentially confounding problem, as many of the proteins that facilitate receptor localization, anchoring, and signaling may be present in both compartments (Carlin et al, 1980; Morciano et al, 2005). One recent study has made strides to address this problem. Using synaptosomes as a starting point, highly enriched PSDs were isolated using density gradient purification, detergent extraction, and pH-based differential extraction. The authors concluded that a PSD fraction can be isolated from human postmortem brain tissues with a reasonable degree of integrity (Hahn et al, 2009). Another group has used high-speed centrifugation, detergents, and sucrose gradients to isolate a ‘detergent-resistant membrane fraction’ that they refer to as membrane microdomains (Behan et al, 2009; Parkin et al, 1999).
These promising fractionation approaches will permit assessment of protein ‘partitioning’ in postmortem tissue. The dynamic nature of protein trafficking suggests that proteins may be rapidly targeted for transit from one subcellular location to another. For example, AMPA receptors may cycle from the plasma membrane to recycling endosomes, and then back to the plasma membrane. In this example, measuring the total level of AMPA receptors would not reflect shifts in the partitioning of receptors between these two compartments, while an approach using fractions would be informative.
Biochemical Assessment of Post-translational Modification in Schizophrenia
Phosphorylation
Studies of signaling cascades have primarily used phospho-specific antibodies and western blot analysis to indirectly probe for changes in kinase activity. Other studies have measured the levels of kinase or phosphatase enzyme, and a few have directly measured kinase or phosphatase activity (Amar et al, 2008; Carty et al, 2012; Castillo et al, 2010; Funk et al, 2012; Ide et al, 2006; Kozlovsky et al, 2001; Rubio et al, 2012; Swatton et al, 2004; Takahashi et al, 2011). Phosphorylation of some kinases, such as c-Jun n-terminal kinase (JNK), is a well-characterized proxy for kinase activity, such that an increase in phosphorylation of the kinase is directly related to increased activity of that kinase (Minden et al, 1994). However, these situations are more the exception than the rule, and most of the data published on signaling cascades and phosphoprotein targets in schizophrenia at best only indirectly reflect changes in kinase activity (Broadbelt and Jones, 2008; Castillo et al, 2010; Emamian et al, 2004; Funk et al, 2012; Kunii et al, 2011; Kunii et al, 2011; Scott et al, 2003).
Postmortem variables such as PMI may also impact phosphorylation (Funk et al, 2012; Li et al, 1996, 2003, 2005; O’Callaghan and Sriram, 2004; Oka et al, 2011). The often transient nature of phosphorylation is well characterized, and in part led to the development of strategies (such as microwave devices) for rapid ‘fixation’ of brain tissues particularly for proteins with a rapid turnover cycle for phosphorylation/dephosphorylation (O’Callaghan and Sriram, 2004). This approach is of course not possible for collection of human brain tissues, and several studies have examined the effects of PMI and other factors on phosphoproteins using rodent tissues. One particularly well-designed study examined >126 proteins using whole-proteome mass spectrometry analysis (Oka et al, 2011). Adult mice were killed and stored at 25 °C or 4 °C for 0, 3, 12, and 72 hours. The authors found that the most of the phosphoprotein data fit into categories, or clusters, based on the length of the PMI and the storage temperature during this interval. At room temperature, the first group of phosphoproteins did not show changes until after 12 hours. In the second group, there was a constant and slow decrease until 72 hours. In the third, there was a rapid decrease until 12 hours, then a gradual slowing out to 72 hours. Different patterns were also observed when the tissues were stored at 4 °C prior to processing, which is the norm for human brain processing. These data indicate that phosphoproteins are variably sensitive to PMI and depend on how quickly the tissues are stored at 4 °C before processing (Oka et al, 2011).
Measuring Kinase Activity: Kinome Arrays
One limitation of using phospho-specific antibodies and western blot analysis is that in most cases these data only indirectly reflect kinase activity. Another limitation is that it is difficult to comprehensively evaluate entire signaling pathways due to the limited availability of phospho-specific antibodies. Further, the amount of postmortem tissue needed to perform western blots and/or enzyme assays on a large number of targets is prohibitive. Recent developments in cancer biology research have led to a novel technique to evaluate phosphorylation of proteins and kinase activity on an ‘omics’ scale. Called ‘kinome’ profiling, a kinome represents the full complement of protein kinase activity in a cell or organism, while the term sub-kinome refers to a specific subset of these enzymes, such as serine/threonine kinases. This new array platform permits interrogation of kinase activity in small volume, low concentration samples, using a chip preprinted with consensus phosphorylation sequences for specific kinases. Two chips are available, one for tyrosine kinases (STK), and another for serine/threonine kinases (STK) (Hilhorst et al, 2009; Sikkema et al, 2009). Each chip has ∼140 unique kinase targets; thus, the pattern and intensity of phosphorylation on the chip yields activity data for potentially hundreds of protein kinases. Activities of specific kinases may be confirmed by running the array in the presence of specific inhibitors or activators to confirm kinase identity. The PamGene kinome array was first used in the field of cancer biology, but its application to testing hypotheses related to abnormalities of signaling in schizophrenia is a promising new development.
The PamGene kinome array platform has been successfully used in cancer biology to examine kinase activity in multiple cancer types, leading to discovery of novel drug targets and therapeutic agents aimed at repairing alterations in the signaling pathways (Zhang and Daly, 2012). For example, kinase-targeting techniques were used to discover that inhibition of heat shock protein 90 may cause destabilization in the MAPK pathway (Haupt et al, 2012). Kinome-wide assays were also used in the discovery of treatment leads for ependymoma, a chemoresistant brain tumor (Atkinson et al, 2011). Finally, targeting of kinase activity in the MET/HGF (hepatocyte growth factor) pathway has led to Phase 3 studies of MET-targeting agents for the treatment of lung cancer (Feng et al, 2012). While this platform has not yet been comprehensively applied to postmortem brain tissues, these studies highlight the potential for drug discovery related to the elucidation of signal transduction anomalies in biological substrates.
Glycosylation
Glycosylation of proteins is a post-translational modification (PTM) that may significantly impact protein function, trafficking, and localization. Gene products that regulate or facilitate glycosylation may be measured with standard approaches, but do not provide a direct measure of glycosylation in the disease substrate. We recently adapted glycosylation gel-shift assays, which employ endoglycosidases, to evaluate N-glycosylation of several molecules in postmortem tissues from subjects with schizophrenia. The excitatory amino acid transporters, EAAT1 and EAAT2 are predominately localized to astrocytes, and account for up to 90% of glutamate reuptake in most brain regions. Glycosylation of these molecules impacts transporter localization and function (Bauer et al, 2010). We found altered N-glycosylation of both of the molecules in the frontal cortex, in a pattern consistent with increased levels of immature glycosyl groups on the transporters (Bauer et al, 2010). We also found a change in the N-glycosylation of AMPA receptor subunit GluA2, in a pattern that suggests a decrease in immature glycosyl groups (Tucholski et al, 2013).
One limitation of the glycosylation shift assays is that the enzymes used are relatively non-specific, and thus do not provide information regarding the composition of the glycans attached to the protein of interest. One alternative method to evaluate the glycome of a specific protein would be to use mass spectrometry, but this approach generally requires a large amount of starting material and enrichment of the sample for the targeted protein, steps which are technically challenging using the postmortem brain substrate. Another promising alternative involves the use of lectins. Lectins are plant-derived proteins that have binding profiles for specific types of N- and O-linked glycans. Lectins can be used to probe western blots or in place of the capture antibody in an immunoisolation. Data derived from so-called ‘lectin blots’ would permit quantification of specific glycosyl groups (based on lectin binding) and identification of the protein to which they are attached (based on size with comparison to western blotting of an adjacent lane or identical blot). For example, we found an alteration in GluA2 protein levels captured with the lectin concanavalin A (ConA), which selectively binds high mannose glycans (Tucholski et al, 2013). Finally, these methods would permit interrogation of sub-glycomes, where complex samples containing large numbers of glycoproteins could be separated electrophoretically, and then probed for binding to lectins that detect different glycosyl groups, yielding a glycosylation fingerprint that reflect patterns of glycosylation in different disease states.
Ubiquitination
Ubiquitination is a PTM that is central to protein trafficking and degradation. However, few studies have examined this process, directly or indirectly, in schizophrenia (Bousman et al, 2010). Several microarray studies have found changes in transcript expression for genes involved in ubiquitination pathways. These hypotheses-generating experiments found changes in individual genes such as ubiquitin specific protease 14 and ubiquitin C-terminal transferase, as well as changes in canonical pathways such as the ubiquitin proteasome pathway (Altar et al, 2005; Bousman et al, 2010; Middleton et al, 2002; Vawter et al, 2001). There are no published studies that directly measure ubiquitination in postmortem tissues from subjects with schizophrenia. Immunoisolation coupled with western blot analysis or mass spectrometry could be used to interrogate poly- or mono- ubiquitinated proteins in postmortem brain.
Palmitoylation and Nitrosylation
These PTMs have not been assessed in postmortem brain tissues from subjects with schizophrenia. However, given the central roles of these biological processes, these represent high-yield targets for postmortem assessment. In particular, the role of palmitoylation in the trafficking and anchoring of receptor complexes in lipid rafts permits partitioning of proteins into subcellular microdomains, which include specialized compartments that facilitate critical cellular functions such as long-term potentiation and long-term depression (Citri and Malenka, 2008; Shan et al, 2013). One promising biochemical technique has been developed that may permit interrogation of palmitoylation in postmortem samples. The biotin switch assay selectively replaces palmitoyl groups with biotin, which may then be detected with streptavidin conjugated to a reporter molecule (Drisdel and Green, 2004; Kang et al, 2008). A similar strategy may be feasible for the detection of nitrosyl groups, using ascorbate to reduce nitrosylated Cys residues and labeling of reduced residues with biotin (Jaffrey, 2005; Jaffrey et al, 2001; Jaffrey and Snyder, 2001).
In summary, other than phosphorylation, only a few studies have examined post-translational modifications in postmortem schizophrenia brain. While the dozen or so studies of phosphorylation have yielded interesting data, they are difficult to interpret as they typically only indirectly reflect changes in kinase activity. Newer methodologies such as the kinome array and lectin blots may provide new avenues for examining protein changes in postmortem brain.
POSTMORTEM STUDIES IN SCHIZOPHRENIA. WHAT QUESTIONS CAN BE ANSWERED USING POSTMORTEM BRAIN?
Results from postmortem schizophrenia studies are often divergent and difficult to interpret. In this section we discuss several examples of postmortem work that illustrate both the difficulties of working with this substrate as well as the potential for these types of studies to contribute to the field.
Data from DISC1 Postmortem Studies
The DISC1 findings from human genetics studies led to several lines of inquiry that have yielded important leads for understanding the etiology of the schizophrenia phenotype (Blackwood et al, 2001). Several groups have found SNPs or other lesions in the DISC1 gene, and several linkage studies have associated the DISC1 locus with schizophrenia in cohorts that are not enriched for the DISC1 translocation (Callicott et al, 2005; Chakirova et al, 2011; Eastwood et al, 2010; Szeszko et al, 2008; Takahashi et al, 2009) (reviewed by Roberts (2007)). The DISC1 protein has multiple functions that likely underlie cognitive function. Polymorphisms in the gene affect cognition in normal individuals, but large GWAS studies did not find an effect for DISC1 in schizophrenia (Badner and Gershon, 2002; Devon et al, 2001; Hodgkinson et al, 2004). The DISC1 protein has specific interacting partners (reviewed by Kamiya et al (2012)) that are associated with its putative role as a large scaffolding protein that facilitates formation of protein complexes (Lipska et al, 2006a, 2006b). In this role, SNPs or other genetic lesions that affect the expression levels, localization, and/or protein-binding motifs of the DISC1 gene could impact formation of protein complexes facilitated by DISC1 protein. These hypotheses have been partially tested in postmortem tissues. DISC1 mRNA levels were unchanged in the hippocampus and dorsolateral prefrontal cortex region in schizophrenia (Lipska et al, 2006a, 2006b). Another study found evidence that the subcellular distribution of DISC1 is altered in schizophrenia (Sawamura et al, 2005). The authors found an increase in the cytoplasm to nucleus ratio for DISC1, suggesting an alteration in mechanisms that localize DISC1, which may include specific protein–protein interactions (Sawamura et al, 2005). Finally, decreased expression of the DISC1 interacting proteins nuclear distribution element-like (NUDEL), fasciculation and elongation protein zeta-1 (FEZ1), and lissencephaly 1 (LIS1) mRNAs was found in the hippocampus and frontal cortex in schizophrenia; each of these measures was associated with the high-risk DISC1 polymorphism (Lipska et al, 2006a, 2006b). Taken together, these postmortem studies suggest that the localization and function of DISC1 may be altered in subjects with schizophrenia lacking the DISC1 translocation.
The interpretation of these data presents significant challenges. These are region-level studies using blended postmortem tissues, which are an admixture of neurons, astroglia, microglia, and endothelial cells. Even fractionated tissues likely contain organelles from multiple cell types, making cell-specific interpretation of a shift in subcellular location a challenge. In addition, most of these subjects have had a lifetime of severe mental illness and antipsychotic treatment, which alters DISC1 mRNA expression in rodents (Chiba et al, 2006). Atypical antipsychotics increased DISC1 mRNA in the frontal cortex, whereas typical antipsychotics did not, raising the issue that typical antipsychotic treatment might normalize decreased DISC1 expression, masking detection of an effect (Chiba et al, 2006). These brains do not represent the biological lesion that presents during development or at the first break of the illness. Rather, they may represent a common pathophysiological lesion found in the mid to late stages of the illness. Such a lesion, consisting of altered DISC1 localization, and shifts in the composition of the protein complexes that DISC1 facilitates, may provide an important translational data point, which could be utilized to extend the hypotheses related to genetic risk associated with the DISC1 gene. For example, immunoisolation of DISC1, followed by quantification of known DISC1 interacting proteins in nuclear and cytoplasmic fractions generated from postmortem brain tissues, could indicate which of these protein–protein interactions warrant the most attention in preclinical studies where medication treatment, PMI, and other factors associated with postmortem tissues are mitigated. Interestingly, an insoluble protein aggresome containing DISC1 may be formed under pathological conditions in neurons, suggesting examination of DISC1-associated proteins as an important direction for further study (Atkin and Kittler, 2012).
AMPA Subtype of Glutamate Receptor Trafficking
The biology of AMPA receptor trafficking involves complex interactions between receptor-interacting molecules that regulate localization of assembled AMPA receptors to and from synapses. Lateral translocation of this receptor complex to the PSD is the mechanism by which synaptic responses are strengthened during some types of long-term potentiation (Contractor and Heinemann, 2002; Lisman and Zhabotinsky, 2001; McGee and Bredt, 2003)(Figure 3). Colocalization of AMPA receptors with NMDA receptors depolarizes the membrane sufficiently for removal of the Mg2+ block from NMDA receptors, permitting activation of these channels. This movement of AMPA receptors following LTP induction is referred to as the regulated receptor pool (Contractor and Heinemann, 2002)(Figure 4). Following insertion at the synapse, AMPA receptors may cycle between the synapse and an intracellular domain, generating a pool of constitutive receptor complexes available for rapid reinsertion to the plasma membrane (Ashby et al, 2004) (Figure 4).

Assembly and modification of neurotransmitter receptors from the nucleus to the dendrite. Neurotransmitter receptors such as the NMDA and AMPA glutamate receptors are synthesized and assembled in the ER before moving through the Golgi. Assembled receptors such as the AMPA and NMDA subtype glutamate receptors may then be phosphorylated and interact with motor proteins which bind the receptors to the microtubules for transport along the dendrite. The receptors then enter the spine where they interact with other anchoring proteins and are inserted at the synapse. AMPA receptor insertion into the synapse consists of direct insertion from the Golgi or indirect insertion and lateral translocation from the constitutive pool. AMPA receptors may also be localized in the regulated pool. NMDA receptors may be directly inserted and are anchored at the postsynaptic density by myriad proteins or may be internalized following phosphorylation. AMPA, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; CaMKII, Ca2+/calmodulin-dependent protein kinases II; CK2, Casein Kinase 2; DLC, Dynein Light Chain; GKAP, Guanylate-kinase-associated Protein; GRIP/ABP, glutamate receptor AMPAR binding protein; KIF, Kinesin Family Member; mLin, Mammalian Lin; MyoV, Myosin V; NF-L, Neurofilament Light Chain; NMDA, N-Methyl-D-aspartate; nNOS, Neuronal Nitric Oxide Synthase; NSF, N-ethylmalemide sensitive factor; PKA, Protein Kinase A; PKC, Protein Kinase C; PP1, Protein Phosphatase 1; PRKCA 1 PICK1, Protein interacting with Protein Kinase C Alpha; PSD95; Post-Synaptic Density 95; SAP97, Synapse-Associated Protein 97; SAP102, Synapse-Associated Protein 102; Stg, Stargazin; 4.1N, Protein 4.1N.

Insertion of neurotransmitter receptors at the synapse. AMPA and NMDA receptors as examples of the complexity of receptor trafficking in the brain. AMPA receptor insertion into the synapse consists of direct insertion from the Golgi or indirect insertion and lateral translocation from the constitutive pool. AMPA receptors may also be localized in the regulated pool. NMDA receptors may be directly inserted and are anchored at the postsynaptic density by myriad proteins or may be internalized following phosphorylation. AMPA, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; CaMKII, Ca2+/calmodulin-dependent protein kinases II; CK2, Casein Kinase 2; GKAP, Guanylate-kinase-associated Protein; GRIP/ABP, glutamate receptor AMPAR binding protein; NF-L, Neurofilament Light Chain; NMDA, N-Methyl-D-aspartate; nNOS, Neuronal Nitric Oxide Synthase; NSF, N-ethylmalemide sensitive factor; PKA, Protein Kinase A; PKC, Protein Kinase C; PP1, Protein Phosphatase 1; PRKCA 1 PICK1, Protein interacting with Protein Kinase C Alpha; PSD95; Post-Synaptic Density 95; SAP97, Synapse-Associated Protein 97; SAP102, Synapse-Associated Protein 102; Stg, Stargazin; 4.1N, Protein 4.1N.
Numerous protein–protein interactions have been described for the trafficking events related to AMPA receptor mobility in dendritic spines. For example, synapse-associated protein 97 (SAP97) is a member of the MAGUK (membrane-associated guanylate kinase) family of proteins and contains three PDZ (postsynaptic density 95/Discs large/zona occludens-1)-binding domains (Müller et al, 1995). These domains are specialized, regulatable motifs that facilitate colocalization of proteins in specialized microdomains such as the PSD. One of the PDZ domains of SAP97 interacts with the PDZ domain on the C terminus of the AMPA receptor GluA1 subunit in GluA1/GluA2 hetero-oligomers (Cai et al, 2002; Leonard et al, 1998).
SAP97, bound to GluA1/GluA2 hetero-oligomers, interacts with the postsynaptic density (Valtschanoff et al, 2000). This interaction is important for the localization and stabilization of GluA1/GluA2 AMPA receptors to the synaptic area (Cai et al, 2006; Rumbaugh et al, 2003). Protein interacting with C kinase 1 (PICK1) links to the PDZ-binding domain of GluA2 and has a role in the clustering of the subunit and tethering of the receptor to the post-synaptic density and plasma membrane (Cao et al, 2007; Jin et al, 2006; Pan et al, 2007). Stargazin also has a role in the trafficking and localization of AMPA receptors at the cell surface, participating in the lateralization of AMPA receptors from extrasynaptic membrane to the synapse (Bats et al, 2007; Bedoukian et al, 2006; Chen et al, 2000). Through interactions with the PDZ domain of GluA2, GRIP1/ABP has a role in maintaining GluA2 containing receptors at the synapse (Dong et al, 1997; Zhang et al, 2001). NSF, another AMPA-interacting protein is an ATPase that facilitates fusion of vesicles with the plasma membrane (Braithwaite et al, 2002).
Converging evidence led to the hypothesis that AMPA receptor trafficking is abnormal in schizophrenia. The observation that phencyclidine (PCP) yields a schizophreniform psychosis in normal subjects and worsens symptoms in subjects with schizophrenia led to the concept that NMDA receptor function is impaired in this illness, because PCP is an open channel blocker of NMDA receptors. As discussed above, one common mechanism for removing the magnesium block from NMDA receptors is colocalization and activation of AMPA receptors in the PSD. PCP administration also leads to increased levels of glutamate, suggesting that non-NMDA receptors may be chronically exposed to higher baseline glutamate levels in synapses where NMDA receptor function is compromised (Shan et al, 2013). Further, the DISC1 and neuregulin1 genetic lesions, animal models, and postmortem data point towards a global deficit in synaptic transmission, consistent with the central role of AMPA receptors as facilitators of long-term potentiation (Malenka and Nicoll, 1999). Finally, there are several reports (reviewed by Meador-Woodruff et al (2001)) of altered AMPA receptor subunit transcript, protein, and binding site expression in several brain regions in schizophrenia. The pattern of these changes suggests that there may be an alteration in AMPA receptor subunit stoichiometry, which can impact receptor localization and function. Thus, AMPA receptor abnormalities may not be a problem of too much or too little receptor, but an alteration in the trafficking pathways described above. Thus, the discovery of specific mechanisms for AMPA receptor localization taken together with these straightforward measures of gene expression in postmortem brain led to the testing of specific hypotheses centered around the AMPA receptor-interacting proteins.
Preclinical data suggest specific hypotheses, related to AMPA receptor trafficking, that are testable with postmortem brain samples. For example, using epitope tags and expression constructs for truncated AMPA receptor-interacting proteins, the functional consequences of diminished N-ethylmaleimide-sensitive factor (NSF) and glutamate receptor-interacting protein (GRIP)/AMPAR binding protein (ABP) were delineated (Braithwaite et al, 2002). A decrease in NSF-GluA2 binding led to increased endocytosis of GluA2 containing AMPA receptors, whereas a decrease in GRIP/ABP-GluA2 binding destabilized accumulation of GluA2 containing AMPA receptors in an intracellular pool (Braithwaite et al, 2002). These data suggest that measuring levels of GRIP/ABP, NSF and/or other AMPA receptor-interacting proteins in postmortem brain may be an indirect measure of alterations in the AMPA receptor trafficking machinery in schizophrenia.
AMPA receptor-interacting protein expression in schizophrenia
Two separate studies reported decreased NSF mRNA and increased NSF protein expression in the dorsolateral prefrontal cortex (DLPFC) (Mirnics et al, 2000; Prabakaran et al, 2004). PICK1 mRNA expression in the DLPFC and the occipital cortex (OCC) was not significantly altered in schizophrenia (Bauer et al, 2008; Dracheva et al, 2005), whereas a different study found decreased expression of PICK1 and increased stargazin in DLPFC in schizophrenia, where both changes were restricted to large cells of cortical layer III (Morrow et al, 2012). In addition, no changes in SAP97 transcript expression were detected in the DLPFC or OCC (Dracheva et al, 2005), but decreased SAP97 protein was detected in a different cohort in the DLPFC (Toyooka et al, 2002a, 2002b) and similarly divergent results were found for cortical GRIP1 expression (Dracheva et al, 2005). Taken together, the data for these AMPA receptor-interacting proteins are inconsistent at best. While differences in postmortem cohort, medication status, region studied, drug use, and other variables may underlie these differences, the small effect sizes, small sample sizes, and inconsistent findings do not support the hypothesis that AMPA receptor trafficking is altered in schizophrenia. Other approaches generally support this conclusion. We did not find alterations in AMPA receptor subunit protein levels in the ER in schizophrenia, and generally found no changes in AMPA subunit proteins in early or recycling endosomes (Hammond et al, 2011; Hammond et al, 2012).
While the notion that AMPA receptor trafficking is altered in schizophrenia was a promising lead, the current balance of the data does not support this conclusion. Further, modulation of AMPA receptors has been evaluated in schizophrenia. Ampakines are compounds that interact with AMPA receptors to modulate synaptic activity by allosterically binding to the receptors to facilitate synaptic transmission (Wezenberg et al, 2007). Meta-analyses of studies using multiple adjunct therapies found that ampakines were ineffective in reducing the positive symptoms of schizophrenia (Goff et al, 2008; Tuominen et al, 2005; Tuominen et al, 2006). A large, placebo-controlled trial of AMPAkine CX516 as adjunct therapy to clozapine, risperidone, or olanzapine did not improve measures of cognition in these patients, and the placebo group had significantly greater improvement in positive and negative symptoms than the group treated with CX516 (Goff et al, 2008). It may be that modulation of AMPA receptors is ineffective because the effects of these compounds are not enough to overcome profound abnormalities of NMDA receptor complexes in the mid to late stages of the illness. Alternatively, AMPA receptor trafficking could be selectively impaired during brain development, and normalize in the adult brain. It would be interesting to evaluate AMPA trafficking and the effects of AMPAkines in high-risk prodromal subjects to determine whether they may contribute to or help prevent development of the illness.
Data from Post-Synaptic Density 95 (PSD95)
PSD95 is an abundant synaptic protein that facilitates clustering of proteins in the PSD microdomain. PSD95 has three PDZ domains, which may reversibly bind myriad interacting proteins including kinases, phosphatases, molecular chaperones, as well as other structural elements and neurotransmitter receptors (Figures 3 and 4). PSD95 forms specific interactions with the GluN2 subunit of the NMDA receptor (Blandini, 2010). NMDA receptors are trafficked to and anchored at the PSD by numerous proteins including PSD95, neuregulin, and SAP102. (Barnes, 2000; Lionel et al, 2013; Weickert et al, 2004). Similar to the DISC1 and neuregulin1 knockout mice, PSD95 homozygous KO’s are not lethal, and the heterozygous animals share prepulse inhibition, hyperactivity, and enhanced LTP phenotypes (Desbonnet et al, 2009; Dyck et al, 2009; Kato et al, 2010; Le Greves et al, 2006; Yao et al, 2004). Interestingly, alterations in the localization and function of DISC1 and neuregulin1 are linked to abnormalities of the NMDA receptor signaling complex that includes PSD95 (Balu and Coyle, 2011; Geddes et al, 2011; Ma et al, 2013; Schmitt et al, 2011). The well-characterized effects of pharmacologic blockade of NMDA receptors, which yield a schizophreniform phenotype, support the hypothesis that disruption of synaptic plasticity, regardless of the mechanism, contributes to the signs and symptoms of schizophrenia. PSD95 is a good candidate for assessment/evaluation in postmortem brain; it is abundantly expressed, it is relatively easy to process/target for mass spectrometry (unpublished observation), it has several high-quality commercially available antibodies, and it has a central role in the formation of protein complexes that comprise the PSD. Thus, several groups have tested the hypothesis that PSD95 expression is abnormal in schizophrenia.
Altered expression of PSD95 in the thalamus
The thalamus comprises discrete nuclei that comprise relay neurons, interneurons and astrocytes that have dense, paired reciprocal projections with multiple cortical areas. Abnormalities of the dorsomedial nucleus of the thalamus have been reported by studies employing imaging, anatomical, and biochemical approaches (reviewed by Meador-Woodruff et al (2003)). Several studies have measured PSD95 mRNA levels in the thalamus. The first study used in situ hybridization to measure levels of PSD95 and several other PSD proteins in several thalamic nuclei including the dorsomedial nucleus (Clinton et al, 2003), and found increased transcript levels for PSD95. Using a different cohort from the same brain bank, a different group did not find changes in PSD95 in the thalamus using QPCR to examine expression in dissected nuclei (Dracheva et al, 2008). The reasons for this failed replication are not clear, but may be due to differences in methodology. In situ hybridization is less sensitive than QPCR, but has more accurate anatomical precision that does not rely on dissection of discrete nuclei. A third replication study was performed using in situ hybridization and tissues from a different brain collection that were much younger (40–50 s vs 70–80 s). Decreased transcripts for PSD95, and two other NMDA interacting proteins found in the PSD, SAP102 and neurofilament-light (NF-L), were detected using in situ hybridization (Clinton and Meador-Woodruff). There is no simple way to reconcile these three divergent findings. PSD95 mRNA expression may have a disease specific-age effect, with lower expression early in the illness, and higher expression later in the illness. The two studies that found changes were performed by the same laboratory using the same techniques. The younger cohort has well-documented substance use in several of the subjects including alcohol and cocaine. Neither study was well controlled for antipsychotic use. In another follow-up study using a different elderly cohort, PSD95 and GluN2A protein levels were increased in the dorsomedial nucleus of the thalamus (Clinton et al, 2006).
PSD95 expression in the frontal cortex
PSD95 transcripts were unchanged in the DLPFC in schizophrenia in tissues from middle-aged subjects from the Stanley Foundation Neuropathology Consortium (Beneyto and Meador-Woodruff, 2008). In a smaller study (n=6 subjects with schizophrenia, n=10 controls) in situ hybridization analysis showed that the expression of PSD95 was significantly decreased in Brodmann area 9 of the prefrontal cortex but not in the hippocampus (Ohnuma et al, 2000). Using tissues from elderly subjects from the Bronx VA brain collection, PSD95 transcripts were increased, and PSD95 protein was decreased in the anterior cingulate cortex (Kristiansen et al, 2006). This finding was replicated in the ACC by the same group in a cohort with overlapping subjects, whereas no change in PSD95 protein was found in the DLPFC (Funk et al, 2009, 2012).
PSD95 expression in other regions
Using immunoautoradiography, a decrease in PSD95 protein was found in the dentate gyrus molecular layer, but not in the dentate hilus of the hippocampus (Toro and Deakin, 2005). Finally, transcripts for PSD95 were not changed in the striatum in schizophrenia (Kristiansen and Meador-Woodruff, 2005).
Taken together, these studies of PSD95 expression measured using different techniques, examining different levels of gene expression, from several different brain collections, suggest an abnormality of PSD95 levels in schizophrenia. The most consistent findings were reported in the frontal cortex, where PSD95 protein levels were decreased in the ACC. One challenge with the interpretation of this data is that it is difficult to elucidate the meaning of a change in a single descriptive measure. PSD95 is localized to a protein complex that forms a specialized microdomain characterized by dynamic changes in its composition, activity, and ultrastructural localization. Accordingly, hypotheses may be formulated based on the biology of receptor trafficking and the roles of receptor-interacting proteins like PSD95 (Lau and Zukin, 2007). For example, one mechanism of diminished NMDA receptor expression could be retention (and degradation) of NMDA receptor complexes in the ER. Decreased expression of PSD95 and GluN2B was found in an ER-enriched fraction in DLPFC from subjects with schizophrenia (but no change in total levels), suggesting increased forward trafficking of NMDA receptor complexes from the ER (Kristiansen et al, 2010). These data suggest that the rate of turnover of NMDA receptor complexes may be altered, with a compensatory increase in trafficking to the synapse. Interestingly, in a separate study, phosphorylation of PSD95 at S295 was increased in the DLPFC. Phosphorylation at this site promotes targeting of PSD95 to the synapse (Kim et al, 2007), a finding consistent with increased forward trafficking of NMDA receptor complexes to the synapse (Funk et al, 2012). In contrast to the DLPFC, phosphorylation of PSD95 at S295 was decreased in the ACC, possibly secondary to diminished overall levels of PSD95 protein in this region (Funk et al, 2012). Samples from these studies were well matched and the results were not attributable to antipsychotic treatment based on secondary analyses of subjects on/off medications at the time of death. In summary, the postmortem data for PSD95 are promising, and additional studies are warranted to fully elucidate the abnormalities of PSD95 in schizophrenia.
A Mechanism for NMDA Receptor Dysfunction in Schizophrenia
Neuregulin1 modulates erbB signaling and neuregulin1 is believed to suppress NMDA receptor activation via this mechanism (Geddes et al, 2011; Golub et al, 2004; Hahn et al, 2006). In postmortem brain from subjects with schizophrenia, there was increased association between PSD95 and the tyrosine kinase erbB4, and an increase in neuregulin1-mediated suppression of NMDA receptor activity (Hahn et al, 2006). These data directly support the hypothesis that NMDA receptor function is altered in schizophrenia, and highlight the putative importance of multipotent proteins such as PSD95 and neuregulin1 in neuropsychiatric illness. We posit that regardless of the etiology or genetic lesion, pathophysiological processes are likely to impact core constituents of synaptic transmission, yielding a common pathological lesion that may or may not be illness-specific.
The PSD95 data also highlight the utility of postmortem studies. The idea to examine PSD95 in schizophrenia stemmed from its characterization as an NMDA receptor-interacting protein, leading to postmortem studies that found changes in regional expression levels of this molecule in schizophrenia. More sophisticated biochemical studies followed, where changes were found suggesting that the NMDA receptor signaling complex, which contains PSD95, is altered in this illness. Next steps could include (1) generating (or further characterizing) a preclinical model that mirrors these findings; can an animal that goes through development with an NMDA receptor signaling complex deficit be rescued? (Delotterie et al, 2010; Nagai et al, 2011, 2) further elucidating the contribution of genetic risk factors on PSD95 biology, and/or (3) developing pharmacological interventions that are targeted not at the underlying genetic risk factors (such as DISC1 in subjects with the DISC1 translocation) but instead at common pathophysiological factors such as impaired NMDA receptor signaling.
In the sections above, we discussed examples of postmortem studies that yielded different conclusions. In the first example postmortem data for DISC1 is promising but descriptive, and is following (but not driving) current thinking in the field. Studies of DISC1 using postmortem tissues could do better, and several testable hypotheses appear feasible with biochemical approaches. The second example related to the AMPA receptor trafficking proteins is an example of postmortem data rejecting (so far) a hypothesis. These studies include data from biochemical approaches that sought to go beyond straightforward measures of gene expression, including analyses of endosomes and ER contents related to AMPA receptor trafficking. The final example is perhaps one of the more developed stories involving postmortem studies in schizophrenia. The juxtaposition of PSD95 as an integral PSD protein as well as an interacting protein for NMDA receptor complexes makes this a high-yield target for postmortem biochemical studies. The PSD95 studies represent a translational synergy between preclinical and postmortem studies. For example, characterization of the effects of phosphorylating S295 on the PSD95 molecule provided a tool that was used to extend the hypothesis that NMDA receptor complex trafficking may be altered in schizophrenia.
SUMMARY AND CONCLUSIONS
There are a few emerging themes for postmortem schizophrenia studies: (1) postmortem data are often disparate, and may be difficult to evaluate and interpret. Factors such as PMI, tissue pH, and antipsychotic drug treatment highlight the myriad complexities of designing these types of studies; (2) there may be selective alterations in trafficking of receptor complexes in schizophrenia. More work is needed to confirm or extend hypotheses related to NMDA receptors, and to determine whether these changes are receptor-specific or instead reflect an abnormality of an underlying biological process (such as receptor trafficking) or compensatory/pathophysiological changes from an intact receptor trafficking system trying to adjust for other molecular deficits found in the illness, and (3) postmortem tissues are an underutilized resource that may be exploited to test and develop cutting edge hypotheses related to the pathophysiology of complex, polygenic illnesses that are difficult to fully model in animals.
The Schizophrenia Phenotype
The schizophrenia phenotype is a heterogeneous agglutination of signs and symptoms of unclear etiology. Genetic predisposition and environmental risk factors synergize to cause a neurodevelopmental lesion that may be quiescent until young adulthood. Animal models recapitulate elements of this syndrome, but many of the symptoms, such as auditory hallucinations and delusions, are probably uniquely human. Another dimension is that strongly implicated genetic lesions such as the neuregulin1 Val to Leu SNP and the DISC1 translocation are rare and alone cannot account for the high prevalence of this illness. However, these multipotent proteins have roles in multiple biological pathways that affect normal brain development as well as cognitive functions in the matured brain, and may contribute to a common pathophysiological ‘lesion’ that settles in after establishment of the illness. A common theme for DISC1, neuregulin1, and PSD95 is that manipulating these gene products impacts neuroplasticity, a core feature of the schizophrenia syndrome (McCullumsmith et al, 2004; Talbot et al, 2004).
Future Directions
There are many promising biochemical techniques that have not been fully adapted to the postmortem substrate. We have discussed a few of these above, including the kinome array, protein QPCR, and capillary electrophoresis. An important direction for the adaptation of biochemical methods is centered on cell-level studies. The postmortem field needs to move away from blending tissue regions and the loss of information that may be associated with this approach. Cell-level studies employing LCM, fractionation, or advanced immunohistochemical localizing approaches will permit more sophisticated assessment of subcellular and cell-specific hypothesis using postmortem brain tissues.
Fluorescence Activated Cell Sorting (FACS) and Single-Cell Mass Cytometry
These novel approaches to examining pathological alterations in individual cells may be useful in postmortem brain where there are lots of unique cell types heterogeneously distributed across large areas. This approach uses FACS to simultaneously measure dozens of antibody labeled proteins on the surface of individual cells, permitting interrogation of the protein milieu expressed on the plasma membrane (Bendall et al, 2011; Bodenmiller et al, 2012; Doerr, 2011). Adaptation of this method to cell preparations from postmortem brain may be challenging, as important elements found in the CNS (such as dendrites and axons) are difficult to preserve when processing or dissociating brain tissue.
Another promising future direction for postmortem tissues is the characterization of subcellular microdomains. The PSD is perhaps the most well-characterized microdomain in the CNS, and several groups have developed cutting edge approaches to interrogate this complex structure in postmortem brain (Cheng et al, 2010; Fernandez et al, 2009; Hahn et al, 2009; MacDonald et al, 2012). Identification and characterization of novel microdomains is currently feasible using standard biochemical techniques. The extensive anatomic, physiologic, and biochemical interactions between neurons and astroglia involves specialized protein complexes and structural elements that have not been fully examined in postmortem tissues from illnesses such as schizophrenia.
Challenges for Postmortem Protein Studies
There are some notable challenges for postmortem studies. Good measures of protein quality or integrity are needed, and the effects of factors like PMI need to be evaluated on a protein-by-protein basis. Most studies rely on tissue samples from brain regions where all of the cell types and extracellular matrix are blended together into the same sample. Data derived from these samples represent the net effects of changes in the dependent measures from mixtures of different cell types and/or subcellular partitions. While many studies include unmedicated subjects or antipsychotic treated rodents, it is difficult to model a lifetime of severe psychiatric illness and antipsychotic treatment, as both may significantly impact brain neurochemistry. Finally, postmortem studies do not capture the illness at its so-called first break. Instead, brains from afflicted subjects are typically collected years or even decades after onset of the illness, and represent a “matured” phase of the illness. Despite these limitations, the postmortem approach has a direct translational nature that is difficult to replicate in an animal model. Data generated from these translational approaches has begun to illuminate putative common pathophysiological pathways in schizophrenia that may lead to the identification of new substrates for drug discovery.
FUNDING AND DISCLOSURE
The authors have nothing to disclose. We gratefully acknowledge support from MH53327(JMW), MH88752 (JMW), MH074016 (REM), MH094445 (REM), and Doris Duke Clinical Scientist Award (REM). The authors declare no conflict of interest.
References
Abekawa T, Ito K, Nakagawa S, Nakato Y, Koyama T (2008). Olanzapine and risperidone block a high dose of methamphetamine-induced schizophrenia-like behavioral abnormalities and accompanied apoptosis in the medial prefrontal cortex. Schizophr Res 101: 84–94.
Akbarian S, Sucher NJ, Bradley D, Tafazzoli A, Trinh D, Hetrick WP et al (1996). Selective alterations in gene expression for NMDA receptor subunits in prefrontal cortex of schizophrenics. J Neurosci 16: 19–30.
Akil M, Edgar CL, Pierri JN, Casali S, Lewis DA (2000). Decreased density of tyrosine hydroxylase-immunoreactive axons in the entorhinal cortex of schizophrenic subjects. Biol Psychiatry 47: 361–370.
Alda M, Ahrens B, Lit W, Dvorakova M, Labelle A, Zvolsky P et al (1996). Age of onset in familial and sporadic schizophrenia. Acta Psychiatr Scand 93: 447–450.
Altar CA, Jurata LW, Charles V, Lemire A, Liu P, Bukhman Y et al (2005). Deficient hippocampal neuron expression of proteasome, ubiquitin, and mitochondrial genes in multiple schizophrenia cohorts. Biol Psychiatry 58: 85–96.
Amar S, Shaltiel G, Mann L, Shamir A, Dean B, Scarr E et al (2008). Possible involvement of post-dopamine D2 receptor signalling components in the pathophysiology of schizophrenia. Int J Neuropsychopharmacol 11: 197–205.
Ashby M, De La Rue S, Ralph G, Uney J, Collingridge G, Henley J (2004). Removal of AMPA receptors (AMPARs) from synapses is preceded by transient endocytosis of extrasynaptic AMPARs. J Neurosci 24: 5172–5176.
Atkin T, Kittler J (2012). DISC1 and the aggresome: A disruption to cellular function? Autophagy 8: 851–852.
Atkinson JM, Shelat AA, Carcaboso AM, Kranenburg TA, Arnold LA, Boulos N et al (2011). An integrated in vitro and in vivo high-throughput screen identifies treatment leads for ependymoma. Cancer Cell 20: 384–399 This study suggests that kinome arrays may be adapted to myriad biological substrates.
Badcock JC (2010). The cognitive neuropsychology of auditory hallucinations: a parallel auditory pathways framework. Schizophr Bull 36: 576–584.
Badner JA, Gershon ES (2002). Meta-analysis of whole-genome linkage scans of bipolar disorder and schizophrenia. Mol Psychiatry 7: 405–411.
Bahn S, Augood SJ, Ryan M, Standaert DG, Starkey M, Emson PC (2001). Gene expression profiling in the post-mortem human brain — no cause for dismay. J Chem Neuroanat 22: 79–94.
Balu DT, Coyle JT (2011). Neuroplasticity signaling pathways linked to the pathophysiology of schizophrenia. Neurosci Biobehav Rev 35: 848–870.
Bannerman DM, Deacon RM, Brady S, Bruce A, Sprengel R, Seeburg PH et al (2004). A comparison of GluR-A-deficient and wild-type mice on a test battery assessing sensorimotor, affective, and cognitive behaviors. Behav Neurosci 118: 643–647.
Barakauskas VE, Beasley CL, Barr AM, Ypsilanti AR, Li HY, Thornton AE et al (2010). A novel mechanism and treatment target for presynaptic abnormalities in specific striatal regions in schizophrenia. Neuropsychopharmacology 35: 1226–1238.
Barnes EM Jr. (2000). Intracellular trafficking of GABA(A) receptors. Life Sci 66: 1063–1070.
Barton AJ, Pearson RC, Najlerahim A, Harrison PJ (1993). Pre- and postmortem influences on brain RNA. J Neurochem 61: 1–11.
Basil B, Adetunji B, Mathews M, Budur K (2006). Trial of risperidone in India--concerns. Br J Psychiatry 188: 489–490.
Bats C, Groc L, Choquet D (2007). The interaction between Stargazin and PSD-95 regulates AMPA receptor surface trafficking. Neuron 53: 719–734 This work demonstrates that specific protein-protein interactions are critical for normal synaptic function.
Bauer D, Haroutunian V, McCullumsmith R, Meador-Woodruff J (2009). Expression of four housekeeping proteins in elderly patients with schizophrenia. J Neural Transm 116: 487–491.
Bauer D, Haroutunian V, Meador-Woodruff JH, McCullumsmith RE (2010). Abnormal glycosylation of EAAT1 and EAAT2 in prefrontal cortex of elderly patients with schizophrenia. Schizophr Res 117: 92–98 The first evidence that glycosylation is altered in schizophrenia in postmortem brain.
Bauer D, McCullumsmith RE, Meador-Woodruff JH (2008). A role for glutamate receptors, transporters, and interacting proteins in cortical dysfunction in schizophrenia. P. O'Donnell (eds) Cortical Deficits in Schizophrenia: From Genes to Function. Springer Science+Business Media, LLC: New York, NY, USA. pp 113–148.
Beckstrøm H, Julsrud L, Haugeto Ø, Dewar D, Graham DI, Lehre KP et al (1999). Interindividual differences in the levels of the glutamate transporters GLAST and GLT, but no clear correlation with Alzheimer's disease. J Neurosci Res 55: 218–229.
Bedoukian M, Weeks A, Partin K (2006). Different domains of the AMPA receptor direct stargazin-mediated trafficking and stargazin-mediated modulation of kinetics. J Biol Chem 281: 23908–23921.
Beerpoot LJ, Lipska BK, Weinberger DR (1996). Neurobiology of treatment-resistant schizophrenia: new insights and new models. Eur Neuropsychopharmacol 6 (Suppl 2): S27–S34.
Behan AT, Byrne C, Dunn MJ, Cagney G, Cotter DR (2009). Proteomic analysis of membrane microdomain-associated proteins in the dorsolateral prefrontal cortex in schizophrenia and bipolar disorder reveals alterations in LAMP, STXBP1 and BASP1 protein expression. Mol Psychiatry 14: 601–613.
Bendall SC, Simonds EF, Qiu P, Amir el-AD, Krutzik PO, Finck R et al (2011). Single-cell mass cytometry of differential immune and drug responses across a human hematopoietic continuum. Science 332: 687–696.
Beneyto M, Meador-Woodruff JH (2008). Lamina-specific abnormalities of NMDA receptor-associated postsynaptic protein transcripts in the prefrontal cortex in schizophrenia and bipolar disorder. Neuropsychopharmacology 33: 2175–2186.
Bennett BM, Reynolds JN, Prusky GT, Douglas RM, Sutherland RJ, Thatcher GR (2007). Cognitive deficits in rats after forebrain cholinergic depletion are reversed by a novel NO mimetic nitrate ester. Neuropsychopharmacology 32: 505–513.
Bhugra D (2005). The global prevalence of schizophrenia. PLoS Med 2: e151.
Blackwood DH, Fordyce A, Walker MT, St Clair DM, Porteous DJ, Muir WJ (2001). Schizophrenia and affective disorders—cosegregation with a translocation at chromosome 1q42 that directly disrupts brain-expressed genes: clinical and P300 findings in a family. Am J Hum Genet 69: 428–433.
Blandini F (2010). An update on the potential role of excitotoxicity in the pathogenesis of Parkinson's disease. Funct Neurol 25: 65–71.
Bodenmiller B, Zunder ER, Finck R, Chen TJ, Savig ES, Bruggner RV et al (2012). Multiplexed mass cytometry profiling of cellular states perturbed by small-molecule regulators. Nat Biotechnol 30: 858–867 Represents a potentially high-yield approach for future studies using postmortem brain tissue.
Borrell J, Vela JM, Arevalo-Martin A, Molina-Holgado E, Guaza C (2002). Prenatal immune challenge disrupts sensorimotor gating in adult rats. Implications for the etiopathogenesis of schizophrenia. Neuropsychopharmacology 26: 204–215.
Bousman CA, Chana G, Glatt SJ, Chandler SD, Lucero GR, Tatro E et al (2010). Preliminary evidence of ubiquitin proteasome system dysregulation in schizophrenia and bipolar disorder: convergent pathway analysis findings from two independent samples. Am J Med Genet B Neuropsychiatr Genet 153B: 494–502.
Bousman CA, Chana G, Glatt SJ, Chandler SD, May T, Lohr J et al (2010). Positive symptoms of psychosis correlate with expression of ubiquitin proteasome genes in peripheral blood. Am J Med Genet B Neuropsychiatr Genet 153B: 1336–1341.
Braithwaite SP, Xia H, Malenka RC (2002). Differential roles for NSF and GRIP/ABP in AMPA receptor cycling. Proc Natl Acad Sci USA 99: 7096–7101.
Breese CR, Freedman R, Leonard SS (1995). Glutamate receptor subtype expression in human postmortem brain tissue from schizophrenics and alcohol abusers. Brain Research 674: 82–90.
Broadbelt K, Jones LB (2008). Evidence of altered calmodulin immunoreactivity in areas 9 and 32 of schizophrenic prefrontal cortex. J Psychiatr Res 42: 612–621.
Bruder GE (1983). Cerebral laterality and psychopathology: a review of dichotic listening studies. Schizophr Bull 9: 134–151.
Bubber P, Tang J, Haroutunian V, Xu H, Davis KL, Blass JP et al (2004). Mitochondrial enzymes in schizophrenia. J Mol Neurosci 24: 315–321.
Buchanan RW, Carpenter WT (2000). Schizophrenia: Introduction and overview. In: Sadock BJ, Sadock VA (eds). Comprehensive Textbook of Psychiatry, Vol 1. Lippincott, Williams, and Wilkins: Philadelphia, PA, USA. pp 1096–1110.
Burnet PW, Eastwood SL, Harrison PJ (1997). [3H]WAY-100635 for 5-HT1A receptor autoradiography in human brain: a comparison with [3H]8-OH-DPAT and demonstration of increased binding in the frontal cortex in schizophrenia. Neurochem Int 30: 565–574.
Butterworth J, Yates CM, Reynolds GP (1985). Distribution of phosphate-activated glutaminase, succinic dehydrogenase, pyruvate dehydrogenase and γ-glutamyl transpeptidase in post-mortem brain from Huntington's disease and agonal cases. J Neurol Sci 67: 161–171.
Cai C, Coleman SK, Niemi K, Keinänen K (2002). Selective binding of synapse-associated protein 97 to GluR-A alpha-amino-5-hydroxy-3-methyl-4-isoxazole propionate receptor subunit is determined by a novel sequence motif. J Biol Chem 277: 31484–31490.
Cai C, Li H, Rivera C, Keinänen K (2006). Interaction between SAP97 and PSD-95, two Maguk proteins involved in synaptic trafficking of AMPA receptors. J Biol Chem 281: 4267–4273.
Cain DF, Kappy MS (1971). Post mortem changes in extractable protein and in tissue pH in the guinea pig brain. J Neurochem 18: 115–121.
Callicott JH, Straub RE, Pezawas L, Egan MF, Mattay VS, Hariri AR et al (2005). Variation in DISC1 affects hippocampal structure and function and increases risk for schizophrenia. Proc Natl Acad Sci USA 102: 8627–8632 Combines fMRI and genetics to formulate hypotheses related to the role(s) of DISC1 variants in severe mental illness.
Cao M, Xu J, Shen C, Kam C, Huganir R, Xia J (2007). PICK1-ICA69 heteromeric BAR domain complex regulates synaptic targeting and surface expression of AMPA receptors. J Neurosci 27: 12945–12956.
Carlin RK, Grab DJ, Cohen RS, Siekevitz P (1980). Isolation and characterization of postsynaptic densities from various brain regions: enrichment of different types of postsynaptic densities. J Cell Biol 86: 831–845 Demonstrates the feasibility of isolating discrete subcellular microdomains in CNS tissues.
Carter MD, Weaver DF, Joudrey HR, Carter AO, Rockwood K (2007). Epilepsy and antiepileptic drug use in elderly people as risk factors for dementia. J Neurol Sci 252: 169–172.
Carty NC, Xu J, Kurup P, Brouillette J, Goebel-Goody SM, Austin DR et al (2012). The tyrosine phosphatase STEP: implications in schizophrenia and the molecular mechanism underlying antipsychotic medications. Transl Psychiatry 2: e137.
Castensson A, Emilsson L, Preece P, Jazin EE (2000). High-resolution quantification of specific mRNA levels in human brain autopsies and biopsies. Genome Res 10: 1219–1229.
Castillo MA, Ghose S, Tamminga CA, Ulery-Reynolds PG (2010). Deficits in syntaxin 1 phosphorylation in schizophrenia prefrontal cortex. Biol Psychiatry 67: 208–216.
Chakirova G, Whalley HC, Thomson PA, Hennah W, Moorhead TW, Welch KA et al (2011). The effects of DISC1 risk variants on brain activation in controls, patients with bipolar disorder and patients with schizophrenia. Psychiatry Res 192: 20–28.
Chandana R, Mythri RB, Mahadevan A, Shankar SK, Srinivas Bharath MM (2009). Biochemical analysis of protein stability in human brain collected at different post-mortem intervals. Indian J Med Res 129: 189–199.
Chen L, Chetkovich D, Petralia R, Sweeney N, Kawasaki Y, Wenthold R et al (2000). Stargazin regulates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature 408: 936–943.
Chen Y, Hancock ML, Role LW, Talmage DA (2010). Intramembranous valine linked to schizophrenia is required for neuregulin 1 regulation of the morphological development of cortical neurons. J Neurosci 30: 9199–9208.
Cheng MC, Lu CL, Luu SU, Tsai HM, Hsu SH, Chen TT et al (2010). Genetic and functional analysis of the DLG4 gene encoding the post-synaptic density protein 95 in schizophrenia. PLoS One 5: e15107.
Chiba S, Hashimoto R, Hattori S, Yohda M, Lipska B, Weinberger DR et al (2006). Effect of antipsychotic drugs on DISC1 and dysbindin expression in mouse frontal cortex and hippocampus. J Neural Trans 113: 1337–1346.
Chubb JE, Bradshaw NJ, Soares DC, Porteous DJ, Millar JK (2008). The DISC locus in psychiatric illness. Mol Psychiatry 13: 36–64.
Citri A, Malenka RC (2008). Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology 33: 18–41.
Clinton SM, Haroutunian V, Davis KL, Meador-Woodruff JH (2003). Altered transcript expression of NMDA receptor-associated postsynaptic proteins in the thalamus of subjects with schizophrenia. Am J Psychiatry 160: 1100–1109.
Clinton SM, Haroutunian V, Meador-Woodruff JH (2006). Up-regulation of NMDA receptor subunit and post-synaptic density protein expression in the thalamus of elderly patients with schizophrenia. J Neurochem 98: 1114–1125.
Clinton SM, Meador-Woodruff JH (2004). Abnormalities of the NMDA Receptor and Associated Intracellular Molecules in the Thalamus in Schizophrenia and Bipolar Disorder. Neuropsychopharmacology 29: 1353–1362.
Colantuoni C, Lipska BK, Ye T, Hyde TM, Tao R, Leek JT et al (2011). Temporal dynamics and genetic control of transcription in the human prefrontal cortex. Nature 478: 519–523.
Contractor A, Heinemann SF (2002). Glutamate receptor trafficking in synaptic plasticity. Sci STKE 2002: re14.
Czudek C, Reynolds GP (1990). [3H]nipecotic acid binding to gamma-aminobutyric acid uptake sites in postmortem human brain. J Neurochem 55: 165–168.
Dean B, Crossland N, Boer S, Scarr E (2008). Evidence for altered post-receptor modulation of the serotonin 2a receptor in schizophrenia. Schizophr Res 104: 185–197.
Dejaegere T, Serneels L, Schafer MK, Van Biervliet J, Horre K, Depboylu C et al (2008). Deficiency of Aph1B/C-gamma-secretase disturbs Nrg1 cleavage and sensorimotor gating that can be reversed with antipsychotic treatment. Proc Natl Acad Sci USA 105: 9775–9780.
Delfs JM, Anegawa NJ, Chesselet MF (1995). Glutamate decarboxylase messenger RNA in rat pallidum: comparison of the effects of haloperidol, clozapine and combined haloperidol-scopolamine treatments. Neuroscience 66: 67–80.
Delotterie D, Ruiz G, Brocard J, Schweitzer A, Roucard C, Roche Y et al (2010). Chronic administration of atypical antipsychotics improves behavioral and synaptic defects of STOP null mice. Psychopharmacology 208: 131–141.
Desbonnet L, Waddington JL, O'Tuathaigh CM (2009). Mutant models for genes associated with schizophrenia. Biochem Soc Trans 37: 308–312.
Devon RS, Anderson S, Teague PW, Burgess P, Kipari TM, Semple CA et al (2001). Identification of polymorphisms within Disrupted in Schizophrenia 1 and Disrupted in Schizophrenia 2, and an investigation of their association with schizophrenia and bipolar affective disorder. Psychiatr Genet 11: 71–78.
Doerr A (2011). A flow cytometry revolution. Nat Methods 8: 531. Discusses the putative applications of "mass cytometry," an approach which may be adaptable to postmortem brain.
Dong H, O'Brien R, Fung E, Lanahan A, Worley P, Huganir R (1997). GRIP: a synaptic PDZ domain-containing protein that interacts with AMPA receptors. Nature 386: 279–284.
Dorph-Petersen KA, Caric D, Saghafi R, Zhang W, Sampson AR, Lewis DA (2009). Volume and neuron number of the lateral geniculate nucleus in schizophrenia and mood disorders. Acta Neuropathol 117: 369–384.
Dracheva S, Byne W, Chin B, Haroutunian V (2008). Ionotropic glutamate receptor mRNA expression in the human thalamus: absence of change in schizophrenia. Brain Res 1214: 23–34.
Dracheva S, Marras SA, Elhakem SL, Kramer FR, Davis KL, Haroutunian V (2001). N-methyl-D-aspartic acid receptor expression in the dorsolateral prefrontal cortex of elderly patients with schizophrenia. Am J Psychiatry 158: 1400–1410.
Dracheva S, McGurk S, Haroutunian V (2005). mRNA expression of AMPA receptors and AMPA receptor binding proteins in the cerebral cortex of elderly schizophrenics. J Neurosci Res 79: 868–878.
Drisdel RC, Green WN (2004). Labeling and quantifying sites of protein palmitoylation. Biotechniques 36: 276–285.
Durrenberger PF, Fernando S, Kashefi SN, Ferrer I, Hauw JJ, Seilhean D et al (2010). Effects of antemortem and postmortem variables on human brain mRNA quality: a BrainNet Europe study. J Neuropathol Exp Neurol 69: 70–81.
Dwivedi Y, Mondal AC, Rizavi HS, Conley RR (2005). Suicide Brain Is Associated with Decreased Expression of Neurotrophins. Biological Psychiatry 58: 315–324.
Dyck BA, Skoblenick KJ, Castellano JM, Ki K, Thomas N, Mishra RK (2009). Behavioral abnormalities in synapsin II knockout mice implicate a causal factor in schizophrenia. Synapse 63: 662–672.
Eastwood SL, Harrison PJ (2001). Synaptic pathology in the anterior cingulate cortex in schizophrenia and mood disorders. A review and a Western blot study of synaptophysin, GAP-43 and the complexins. Brain Res Bull 55: 569–578.
Eastwood SL, Harrison PJ (2005). Decreased expression of vesicular glutamate transporter 1 and complexin II mRNAs in schizophrenia: further evidence for a synaptic pathology affecting glutamate neurons. Schizophr Res 73: 159–172.
Eastwood SL, Walker M, Hyde TM, Kleinman JE, Harrison PJ (2010). The DISC1 Ser704Cys substitution affects centrosomal localization of its binding partner PCM1 in glia in human brain. Hum Mol Genet 19: 2487–2496.
Eggan K (2008). Using stem cells and reprogramming to understand disease. Regen Med 3: 799–801.
Eggan SM, Hashimoto T, Lewis DA (2008). Reduced cortical cannabinoid 1 receptor messenger RNA and protein expression in schizophrenia. Arch Gen Psychiatry 65: 772–784.
Emamian ES, Karayiorgou M, Gogos JA (2004). Decreased phosphorylation of NMDA receptor type 1 at serine 897 in brains of patients with Schizophrenia. J Neurosci 24: 1561–1564.
English JA, Dicker P, Focking M, Dunn MJ, Cotter DR (2009). 2-D DIGE analysis implicates cytoskeletal abnormalities in psychiatric disease. Proteomics 9: 3368–3382 Adapts 2D electrophoresis and mass spectrometry to assess protein expression in postmortem brain in schizophrenia.
English JA, Pennington K, Dunn MJ, Cotter DR (2011). The neuroproteomics of schizophrenia. Biol Psychiatry 69: 163–172.
Ervin JF, Heinzen EL, Cronin KD, Goldstein D, Szymanski MH, Burke JR et al (2007). Postmortem delay has minimal effect on brain RNA integrity. J Neuropathol Exp Neurol 66: 1093–1099.
Fanous AH, Zhou B, Aggen SH, Bergen SE, Amdur RL, Duan J et al (2012). Genome-wide association study of clinical dimensions of schizophrenia: polygenic effect on disorganized symptoms. Am J Psychiatry 169: 1309–1317.
Fanselow MS, Dong HW (2010). Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 65: 7–19.
Featherstone RE, Rizos Z, Nobrega JN, Kapur S, Fletcher PJ (2007). Gestational methylazoxymethanol acetate treatment impairs select cognitive functions: parallels to schizophrenia. Neuropsychopharmacology 32: 483–492.
Feng Y, Thiagarajan PS, Ma PC (2012). MET signaling: novel targeted inhibition and its clinical development in lung cancer. J Thorac Oncol 7: 459–467.
Fernandez E, Collins MO, Uren RT, Kopanitsa MV, Komiyama NH, Croning MD et al (2009). Targeted tandem affinity purification of PSD-95 recovers core postsynaptic complexes and schizophrenia susceptibility proteins. Mol Syst Biol 5: 269.
Ferrer I, Santpere G, Arzberger T, Bell J, Blanco R, Boluda S et al (2007). Brain protein preservation largely depends on the postmortem storage temperature: implications for study of proteins in human neurologic diseases and management of brain banks: a BrainNet Europe Study. J Neuropathol Exp Neurol 66: 35–46.
Fleige S, Pfaffl MW (2006). RNA integrity and the effect on the real-time qRT-PCR performance. Mol Aspects Med 27: 126–139.
Fountoulakis M, Hardmeier R, Hoger H, Lubec G (2001). Postmortem changes in the level of brain proteins. Exp Neurol 167: 86–94.
Fredriksson S, Gullberg M, Jarvius J, Olsson C, Pietras K, Gustafsdottir SM et al (2002). Protein detection using proximity-dependent DNA ligation assays. Nat Biotechnol 20: 473–477 Basis for novel protein PCR assays now commercially available for measuring low abundance proteins.
Freyberg Z, Ferrando SJ, Javitch JA (2010). Roles of the Akt/GSK-3 and Wnt signaling pathways in schizophrenia and antipsychotic drug action. Am J Psychiatry 167: 388–396.
Funk A, Rumbaugh G, Harotunian V, McCullumsmith R, Meador-Woodruff J (2009). Decreased expression of NMDA receptor-associated proteins in frontal cortex of elderly patients with schizophrenia. Neuroreport 20: 1019–1022.
Funk AJ, McCullumsmith RE, Haroutunian V, Meador-Woodruff JH (2012). Abnormal activity of the MAPK- and cAMP-associated signaling pathways in frontal cortical areas in postmortem brain in schizophrenia. Neuropsychopharmacology 37: 896–905.
Geddes AE, Huang X-F, Newell KA (2011). Reciprocal signalling between NR2 subunits of the NMDA receptor and neuregulin1 and their role in schizophrenia. Prog Neuro-Psychopharmacol Biol Psychiatry 35: 896–904.
Gershon ES, Alliey-Rodriguez N, Liu C (2011). After GWAS: searching for genetic risk for schizophrenia and bipolar disorder. Am J Psychiatry 168: 253–256.
Gilmore JH, Lawler CP, Eaton AM, Mailman RB (1993). Postmortem stability of dopamine D1 receptor mRNA and D1 receptors. Brain Res Mol Brain Res 18: 290–296.
Glantz LA, Lewis DA (1997). Reduction of synaptophysin immunoreactivity in the prefrontal cortex of subjects with schizophrenia. Regional and diagnostic specificity. Arch Gen Psychiatry 54: 943–952.
Goff D, Lamberti J, Leon A, Green M, Miller A, Patel J et al (2008). A placebo-controlled add-on trial of the Ampakine, CX516, for cognitive deficits in schizophrenia. Neuropsychopharmacology 33: 465–472.
Golub MS, Germann SL, Lloyd KC (2004). Behavioral characteristics of a nervous system-specific erbB4 knock-out mouse. Behav Brain Res 153: 159–170.
Gonzalez-Maeso J, Torre I, Rodriguez-Puertas R, Garcia-Sevilla JA, Guimon J, Meana JJ (2002). Effects of age, postmortem delay and storage time on receptor-mediated activation of G-proteins in human brain. Neuropsychopharmacology 26: 468–478.
Gullberg M, Gustafsdottir SM, Schallmeiner E, Jarvius J, Bjarnegard M, Betsholtz C et al (2004). Cytokine detection by antibody-based proximity ligation. Proc Natl Acad Sci USA 101: 8420–8424 Application of protein PCR to small, soluble proteins in complex biological samples.
Guo X, Hamilton P, Reish N, Sweatt J, Miller C, Rumbaugh G (2009). Reduced Expression of the NMDA Receptor-Interacting Protein SynGAP Causes Behavioral Abnormalities that Model Symptoms of Schizophrenia. Neuropsychopharmacology 34: 1659–1672.
Hahn CG, Banerjee A, Macdonald ML, Cho DS, Kamins J, Nie Z et al (2009). The post-synaptic density of human postmortem brain tissues: an experimental study paradigm for neuropsychiatric illnesses. PLoS One 4: e5251 Develops a novel approach to isolate and characterize PSDs in postmortem brain.
Hahn CG, Wang HY, Cho DS, Talbot K, Gur RE, Berrettini WH et al (2006). Altered neuregulin 1-erbB4 signaling contributes to NMDA receptor hypofunction in schizophrenia. Nat Med 12: 824–828 Uses standard biochemical approaches with postmortem brain to demonstrate abnormalities of protein-protein interactions in schizophrenia.
Haller J, Szirmai M, Varga B, Ledent C, Freund TF (2005). Cannabinoid CB1 receptor dependent effects of the NMDA antagonist phencyclidine in the social withdrawal model of schizophrenia. Behav Pharmacol 16: 415–422.
Hammond J, McCullumsmith RE, Haroutunian V, Meador-Woodruff JH (2011). Endosomal trafficking of AMPA receptors in frontal cortex of elderly patients with schizophrenia. Schizophr Res 130: 260–265.
Hammond JC, McCullumsmith RE, Funk AJ, Haroutunian V, Meador-Woodruff JH (2010). Evidence for abnormal forward trafficking of AMPA receptors in frontal cortex of elderly patients with schizophrenia. Neuropsychopharmacology 35: 2110–2119 First postmortem study to isolate and characterize endosomes.
Hammond JC, McCullumsmith RE, Haroutunian V, Meador-Woodruff JH (2011). Endosomal trafficking of AMPA receptors in frontal cortex of elderly patients with schizophrenia. Schizophr Res 130: 260–265.
Hammond JC, Meador-Woodruff JH, Haroutunian V, McCullumsmith RE (2012). AMPA receptor subunit expression in the endoplasmic reticulum in frontal cortex of elderly patients with schizophrenia. PLoS One 7: e39190 Adapts standard biochemical approaches to postmortem brain to isolate ER microsomes in tissues from subjects with schizophrenia.
Hanlon FM, Sutherland RJ (2000). Changes in adult brain and behavior caused by neonatal limbic damage: implications for the etiology of schizophrenia. Behav Brain Res 107: 71–83.
Hardy JA, Wester P, Winblad B, Gezelius C, Bring G, Eriksson A (1985). The patients dying after long terminal phase have acidotic brains; implications for biochemical measurements on autopsy tissue. J Neural Transm 61: 253–264.
Haroutunian V, Katsel P, Dracheva S, Davis KL (2006). The human homolog of the QKI gene affected in the severe dysmyelination "quaking" mouse phenotype: downregulated in multiple brain regions in schizophrenia. Am J Psychiatry 163: 1834–1837.
Harrison PJ, Heath PR, Eastwood SL, Burnet PW, McDonald B, Pearson RC (1995). The relative importance of premortem acidosis and postmortem interval for human brain gene expression studies: selective mRNA vulnerability and comparison with their encoded proteins. Neurosci Lett 200: 151–154.
Harrison PJ, Procter AW, Barton AJ, Lowe SL, Najlerahim A, Bertolucci PH et al (1991). Terminal coma affects messenger RNA detection in post mortem human temporal cortex. Brain Res Mol Brain Res 9: 161–164.
Hasler G, Drevets WC, Manji HK, Charney DS (2004). Discovering endophenotypes for major depression. Neuropsychopharmacology 29: 1765–1781.
Haupt A, Joberty G, Bantscheff M, Frohlich H, Stehr H, Schweiger MR et al (2012). Hsp90 inhibition differentially destabilises MAP kinase and TGF-beta signalling components in cancer cells revealed by kinase-targeted chemoproteomics. BMC Cancer 12: 38.
Hazlett EA, Speiser LJ, Goodman M, Roy M, Carrizal M, Wynn JK et al (2007). Exaggerated affect-modulated startle during unpleasant stimuli in borderline personality disorder. Biol Psychiatry 62: 250–255.
Hikida T, Jaaro-Peled H, Seshadri S, Oishi K, Hookway C, Kong S et al (2007). Dominant-negative DISC1 transgenic mice display schizophrenia-associated phenotypes detected by measures translatable to humans. Proc Natl Acad Sci USA 104: 14501–14506.
Hilhorst R, Houkes L, van den Berg A, Ruijtenbeek R (2009). Peptide microarrays for detailed, high-throughput substrate identification, kinetic characterization, and inhibition studies on protein kinase A. Anal Biochem 387: 150–161 Uses kinome arrays to characterize/validate protein kinase A function.
Hodgkinson CA, Goldman D, Jaeger J, Persaud S, Kane JM, Lipsky RH et al (2004). Disrupted in schizophrenia 1 (DISC1): association with schizophrenia, schizoaffective disorder, and bipolar disorder. Am J Hum Genet 75: 862–872.
Holmseth S, Dehnes Y, Huang YH, Follin-Arbelet VV, Grutle NJ, Mylonakou MN et al (2012). The density of EAAC1 (EAAT3) glutamate transporters expressed by neurons in the mammalian CNS. J Neurosci 32: 6000–6013.
Huang H-S, Matevossian A, Jiang Y, Akbarian S (2006). Chromatin immunoprecipitation in postmortem brain. J Neurosci Methods 156: 284–292.
Huerta I, McCullumsmith RE, Haroutunian V, Gimenez-Amaya JM, Meador-Woodruff JH (2006). Expression of excitatory amino acid transporter interacting protein transcripts in the thalamus in schizophrenia. Synapse 59: 394–402.
Huotari M, Garcia-Horsman JA, Karayiorgou M, Gogos JA, Mannisto PT (2004). D-amphetamine responses in catechol-O-methyltransferase (COMT) disrupted mice. Psychopharmacology (Berl) 172: 1–10.
Ibarra C, Nedvetsky PI, Gerlach M, Riederer P, Schmidt HHHW (2001). Regional and age-dependent expression of the nitric oxide receptor, soluble guanylyl cyclase, in the human brain. Brain Res 907: 54–60.
Ide M, Ohnishi T, Murayama M, Matsumoto I, Yamada K, Iwayama Y et al (2006). Failure to support a genetic contribution of AKT1 polymorphisms and altered AKT signaling in schizophrenia. J Neurochem 99: 277–287.
Jaffrey SR (2005). Detection and characterization of protein nitrosothiols. Methods Enzymol 396: 105–118.
Jaffrey SR, Erdjument-Bromage H, Ferris CD, Tempst P, Snyder SH (2001). Protein S-nitrosylation: a physiological signal for neuronal nitric oxide. Nat Cell Biol 3: 193–197.
Jaffrey SR, Snyder SH (2001). The biotin switch method for the detection of S-nitrosylated proteins. Sci STKE 2001: pl1.
Jentsch JD, Roth RH (1999). The neuropsychopharmacology of phencyclidine: from NMDA receptor hypofunction to the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 20: 201–225.
Jin W, Ge W, Xu J, Cao M, Peng L, Yung W et al (2006). Lipid binding regulates synaptic targeting of PICK1, AMPA receptor trafficking, and synaptic plasticity. J Neurosci 26: 2380–2390.
Johnston NL, Cervenak J, Shore AD, Torrey EF, Yolken RH (1997). Multivariate analysis of RNA levels from postmortem human brains as measured by three different methods of RT-PCR. Stanley Neuropathology Consortium. J Neurosci Methods 77: 83–92.
Joyce JN, Meador-Woodruff JH (1997). Linking the family of D2 receptors to neuronal circuits in human brain: insights into schizophrenia. Neuropsychopharmacology 16: 375–384.
Kamiya A, Sedlak TW, Pletnikov MV (2012). DISC1 pathway in brain development: exploring therapeutic targets for major psychiatric disorders. Front Psychiatry 3: 25.
Kang R, Wan J, Arstikaitis P, Takahashi H, Huang K, Bailey AO et al (2008). Neural palmitoyl-proteomics reveals dynamic synaptic palmitoylation. Nature 456: 904–909.
Kato T, Kasai A, Mizuno M, Fengyi L, Shintani N, Maeda S et al (2010). Phenotypic characterization of transgenic mice overexpressing Neuregulin-1. PLoS One 5: e14185.
Kay SR (1990). Significance of the positive-negative distinction in schizophrenia. Schizophr Bull 16: 635–652.
Kim MJ, Futai K, Jo J, Hayashi Y, Cho K, Sheng M (2007). Synaptic accumulation of PSD-95 and synaptic function regulated by phosphorylation of serine-295 of PSD-95. Neuron 56: 488–502 Characterizes the role(s) of serine-295 phosphorylation of PSD95, providing a tool that may be adapted to postmortem studies..
Kingsbury AE, Foster OJ, Nisbet AP, Cairns N, Bray L, Eve DJ et al (1995). Tissue pH as an indicator of mRNA preservation in human post-mortem brain. Brain Res Mol Brain Res 28: 311–318.
Knapp M, Mangalore R, Simon J (2004). The global costs of schizophrenia. Schizophr Bull 30: 279–293.
Konopaske GT, Dorph-Petersen KA, Sweet RA, Pierri JN, Zhang W, Sampson AR et al (2008). Effect of chronic antipsychotic exposure on astrocyte and oligodendrocyte numbers in macaque monkeys. Biol Psychiatry 63: 759–765.
Kozlovsky N, Belmaker RH, Agam G (2001). Low GSK-3 activity in frontal cortex of schizophrenic patients. Schizophr Res 52: 101–105.
Kristiansen L, Beneyto M, Haroutunian V, Meador-Woodruff J (2006). Changes in NMDA receptor subunits and interacting PSD proteins in dorsolateral prefrontal and anterior cingulate cortex indicate abnormal regional expression in schizophrenia. Mol Psychiatry 11: 737–747 705.
Kristiansen L, Patel S, Haroutunian V, Meador-Woodruff J (2010). Expression of the NR2B-NMDA receptor subunit and its Tbr-1/CINAP regulatory proteins in postmortem brain suggest altered receptor processing in schizophrenia. Synapse 64: 495–502.
Kristiansen LV, Meador-Woodruff JH (2005). Abnormal striatal expression of transcripts encoding NMDA interacting PSD proteins in schizophrenia, bipolar disorder and major depression. Schizophr Res 78: 87–93.
Kunii Y, Ikemoto K, Wada A, Yang Q, Kusakabe T, Suzuki T et al (2011). Detailed DARPP-32 expression profiles in postmortem brains from patients with schizophrenia: an immunohistochemical study. Med Mol Morphol 44: 190–199.
Kunii Y, Yabe H, Wada A, Yang Q, Nishiura K, Niwa S (2011). Altered DARPP-32 expression in the superior temporal gyrus in schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry 35: 1139–1143.
Laruelle M, Abi-Dargham A, Gil R, Kegeles L, Innis R (1999). Increased dopamine transmission in schizophrenia: relationship to illness phases. Biol Psychiatry 46: 56–72.
Lau CG, Zukin RS (2007). NMDA receptor trafficking in synaptic plasticity and neuropsychiatric disorders. Nat Rev Neurosci 8: 413–426.
Law AJ, Weickert CS, Hyde TM, Kleinman JE, Harrison PJ (2004). Reduced spinophilin but not microtubule-associated protein 2 expression in the hippocampal formation in schizophrenia and mood disorders: molecular evidence for a pathology of dendritic spines. Am J Psychiatry 161: 1848–1855.
Le Corre S, Harper CG, Lopez P, Ward P, Catts S (2000). Increased levels of expression of an NMDARI splice variant in the superior temporal gyrus in schizophrenia. Neuroreport 11: 983–986.
Le Greves M, Zhou Q, Berg M, Le Greves P, Fholenhag K, Meyerson B et al (2006). Growth hormone replacement in hypophysectomized rats affects spatial performance and hippocampal levels of NMDA receptor subunit and PSD-95 gene transcript levels. Exp Brain Res 173: 267–273.
Lee FJ, Pei L, Liu F (2009). Disruption of the dopamine transporter-dopamine D2 receptor interaction in schizophrenia. Synapse 63: 710–712.
Lee PR, Brady DL, Shapiro RA, Dorsa DM, Koenig JI (2005). Social interaction deficits caused by chronic phencyclidine administration are reversed by oxytocin. Neuropsychopharmacology 30: 1883–1894.
Lee PR, Brady DL, Shapiro RA, Dorsa DM, Koenig JI (2007). Prenatal stress generates deficits in rat social behavior: Reversal by oxytocin. Brain Res 1156: 152–167.
Leonard AS, Davare MA, Horne MC, Garner CC, Hell JW (1998). SAP97 is associated with the alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor GluR1 subunit. J Biol Chem 273: 19518–19524.
Leonard S, Logel J, Luthman D, Casanova M, Kirch D, Freedman R (1993). Biological stability of mRNA isolated from human postmortem brain collections. Biol Psychiatry 33: 456–466.
Lepage M, Montoya A, Pelletier M, Achim AM, Menear M, Lal S (2006). Associative memory encoding and recognition in schizophrenia: an event-related fMRI study. Biol Psychiatry 60: 1215–1223.
Levinson DF, Shi J, Wang K, Oh S, Riley B, Pulver AE et al (2012). Genome-wide association study of multiplex schizophrenia pedigrees. Am J Psychiatry 169: 963–973.
Lewis DA, Volk DW, Hashimoto T (2004). Selective alterations in prefrontal cortical GABA neurotransmission in schizophrenia: a novel target for the treatment of working memory dysfunction. Psychopharmacology (Berl) 174: 143–150 Develops the idea that selective modulation of GABA receptors may restore working memory in limbic circuits.
Li J, Gould TD, Yuan P, Manji HK, Chen G (2003). Post-mortem interval effects on the phosphorylation of signaling proteins. Neuropsychopharmacology 28: 1017–1025.
Li JZ, Vawter MP, Walsh DM, Tomita H, Evans SJ, Choudary PV et al (2004). Systematic changes in gene expression in postmortem human brains associated with tissue pH and terminal medical conditions. Hum Mol Genet 13: 609–616.
Li X, Friedman AB, Roh M-S, Jope RS (2005). Anesthesia and post-mortem interval profoundly influence the regulatory serine phosphorylation of glycogen synthase kinase-3 in mouse brain. J Neurochem 92: 701–704.
Li X, Greenwood AF, Powers R, Jope RS (1996). Effects of postmortem interval, age, and Alzheimer's disease on G-proteins in human brain. Neurobiol Aging 17: 115–122.
Lieberman JA, Stroup TS, McEvoy JP, Swartz MS, Rosenheck RA, Perkins DO et al (2005). Effectiveness of antipsychotic drugs in patients with chronic schizophrenia. N Engl J Med 353: 1209–1223.
Lijam N, Paylor R, McDonald MP, Crawley JN, Deng CX, Herrup K et al (1997). Social interaction and sensorimotor gating abnormalities in mice lacking Dvl1. Cell 90: 895–905.
Lionel AC, Vaags AK, Sato D, Gazzellone MJ, Mitchell EB, Chen HY et al (2013). Rare exonic deletions implicate the synaptic organizer Gephyrin (GPHN) in risk for autism, schizophrenia and seizures. Hum Mol Genet 22: 2055–2066.
Lipska BK, Deep-Soboslay A, Weickert CS, Hyde TM, Martin CE, Herman MM et al (2006a). Critical factors in gene expression in postmortem human brain: Focus on studies in schizophrenia. Biol Psychiatry 60: 650–658.
Lipska BK, Peters T, Hyde TM, Halim N, Horowitz C, Mitkus S et al (2006b). Expression of DISC1 binding partners is reduced in schizophrenia and associated with DISC1 SNPs. Hum Mol Genet 15: 1245–1258.
Lisman JE, Zhabotinsky AM (2001). A model of synaptic memory: a CaMKII/PP1 switch that potentiates transmission by organizing an AMPA receptor anchoring assembly. Neuron 31: 191–201.
Ma TM, Abazyan S, Abazyan B, Nomura J, Yang C, Seshadri S et al (2013). Pathogenic disruption of DISC1-serine racemase binding elicits schizophrenia-like behavior via D-serine depletion. Mol Psychiatry 18: 557–567.
MacDonald ML, Ciccimaro E, Prakash A, Banerjee A, Seeholzer SH, Blair IA et al (2012). Biochemical fractionation and stable isotope dilution liquid chromatography-mass spectrometry for targeted and microdomain-specific protein quantification in human postmortem brain tissue. Mol Cell Proteomics 11: 1670–1681.
Malenka RC, Nicoll RA (1999). Long-term potentiation--a decade of progress? Science 285: 1870–1874.
Marballi K, Cruz D, Thompson P, Walss-Bass C (2012). Differential neuregulin 1 cleavage in the prefrontal cortex and hippocampus in schizophrenia and bipolar disorder: preliminary findings. PLoS One 7: e36431 Develops an important hypothesis using postmortem brain that neuregulin1 cleavage may have a role in the pathophysiology of schizophrenia.
Marballi K, Quinones MP, Jimenez F, Escamilla MA, Raventos H, Soto-Bernardini MC et al (2010). In vivo and in vitro genetic evidence of involvement of neuregulin 1 in immune system dysregulation. J Mol Med (Berl) 88: 1133–1141.
Marenco S, Weinberger DR (2000). The neurodevelopmental hypothesis of schizophrenia: following a trail of evidence from cradle to grave. Dev Psychopathol 12: 501–527.
McCullumsmith R, Clinton S, Meador-Woodruff J (2004). Schizophrenia as a disorder of neuroplasticity. Int Rev Neurobiol 59: 19–45 One of the first reviews to argue for schizophrenia as a disorder of neuroplasticity.
McCullumsmith RE, Kristiansen LV, Beneyto M, Scarr E, Dean B, Meador-Woodruff JH (2007). Decreased NR1, NR2A, and SAP102 transcript expression in the hippocampus in bipolar disorder. Brain Res 1127: 108–118.
McCullumsmith RE, Meador-Woodruff JH (2002). Striatal excitatory amino acid transporter transcript expression in schizophrenia, bipolar disorder, and major depressive disorder. Neuropsychopharmacology 26: 368–375.
McCullumsmith RE, Meador-Woodruff JH (2011). Novel approaches to the study of postmortem brain in psychiatric illness: old limitations and new challenges. Biol Psychiatry 69: 127–133.
McGee AW, Bredt DS (2003). Assembly and plasticity of the glutamatergic postsynaptic specialization. Curr Opin Neurobiol 13: 111–118.
Meador-Woodruff J, Clinton S, Beneyto M, McCullumsmith R (2003). Molecular abnormalities of the glutamate synapse in the thalamus in schizophrenia. Ann NY Acad Sci 1003: 75–93.
Meador-Woodruff JH, Hogg AJ Jr., Smith RE (2001). Striatal ionotropic glutamate receptor expression in schizophrenia, bipolar disorder, and major depressive disorder. Brain Res Bull 55: 631–640.
Meyer JM, Nasrallah HA, McEvoy JP, Goff DC, Davis SM, Chakos M et al (2005). The Clinical Antipsychotic Trials Of Intervention Effectiveness (CATIE) Schizophrenia Trial: clinical comparison of subgroups with and without the metabolic syndrome. Schizophr Res 80: 9–18.
Middleton FA, Mirnics K, Pierri JN, Lewis DA, Levitt P (2002). Gene expression profiling reveals alterations of specific metabolic pathways in schizophrenia. J Neurosci 22: 2718–2729.
Millar JK, Wilson-Annan JC, Anderson S, Christie S, Taylor MS, Semple CA et al (2000). Disruption of two novel genes by a translocation co-segregating with schizophrenia. Hum Mol Genet 9: 1415–1423.
Minden A, Lin A, Smeal T, Derijard B, Cobb M, Davis R et al (1994). c-Jun N-terminal phosphorylation correlates with activation of the JNK subgroup but not the ERK subgroup of mitogen-activated protein kinases. Mol Cell Biol 14: 6683–6688.
Miner LA, Ostrander M, Sarter M (1997). Effects of ibotenic acid-induced loss of neurons in the medial prefrontal cortex of rats on behavioral vigilance: evidence for executive dysfunction. J Psychopharmacol 11: 169–178.
Mirnics K, Middleton F, Marquez A, Lewis D, Levitt P (2000). Molecular characterization of schizophrenia viewed by microarray analysis of gene expression in prefrontal cortex. Neuron 28: 53–67.
Mohn AR, Gainetdinov RR, Caron MG, Koller BH (1999). Mice with reduced NMDA receptor expression display behaviors related to schizophrenia. Cell 98: 427–436.
Morciano M, Burre J, Corvey C, Karas M, Zimmermann H, Volknandt W (2005). Immunoisolation of two synaptic vesicle pools from synaptosomes: a proteomics analysis. J Neurochem 95: 1732–1745.
Morrow JA, Maclean JKF, Jamieson C (2012). AMPA Receptor Positive Modulators Albert JS, Wood MW, Targets and Emerging Therapies for Schizophrenia. John Wiley & Sons: Hoboken, NJ, USA, 187–218.
Müller BM, Kistner U, Veh RW, Cases-Langhoff C, Becker B, Gundelfinger ED et al (1995). Molecular characterization and spatial distribution of SAP97, a novel presynaptic protein homologous to SAP90 and the Drosophila discs-large tumor suppressor protein. J Neurosci 15: 2354–2366.
Nagai T, Kitahara Y, Ibi D, Nabeshima T, Sawa A, Yamada K (2011). Effects of antipsychotics on the behavioral deficits in human dominant-negative DISC1 transgenic mice with neonatal polyI:C treatment. Behav Brain Res 225: 305–310.
O'Neill RA, Bhamidipati A, Bi X, Deb-Basu D, Cahill L, Ferrante J et al (2006). Isoelectric focusing technology quantifies protein signaling in 25 cells. Proc Natl Acad Sci USA 103: 16153–16158 Demonstrates that capillary electrophoresis may have sufficient sensitivity to be combined with laser-capture microdissection.
O’Callaghan JP, Sriram K (2004). Focused microwave irradiation of the brain preserves in vivo protein phosphorylation: comparison with other methods of sacrifice and analysis of multiple phosphoproteins. J Neurosci Methods 135: 159–168.
Ohnuma T, Kato H, Arai H, Faull RL, McKenna PJ, Emson PC (2000). Gene expression of PSD95 in prefrontal cortex and hippocampus in schizophrenia. Neuroreport 11: 3133–3137.
Oka T, Tagawa K, Ito H, Okazawa H (2011). Dynamic changes of the phosphoproteome in postmortem mouse brains. PLoS One 6: e21405.
Oni-Orisan A, Kristiansen L, Haroutunian V, Meador-Woodruff J, McCullumsmith R (2008). Altered vesicular glutamate transporter expression in the anterior cingulate cortex in schizophrenia. Biol Psychiatry 63: 766–775.
Oni-Orisan A, Kristiansen LV, Haroutunian V, Meador-Woodruff JH, McCullumsmith RE (2008). Altered vesicular glutamate transporter expression in the anterior cingulate cortex in schizophrenia. Biol Psychiatry 63: 766–775.
Pan L, Wu H, Shen C, Shi Y, Jin W, Xia J et al (2007). Clustering and synaptic targeting of PICK1 requires direct interaction between the PDZ domain and lipid membranes. EMBO J 26: 4576–4587.
Pardue S, Zimmerman AL, Morrison-Bogorad M (1994). Selective postmortem degradation of inducible heat shock protein 70 (hsp70) mRNAs in rat brain. Cell Mol Neurobiol 14: 341–357.
Parkin ET, Hussain I, Karran EH, Turner AJ, Hooper NM (1999). Characterization of detergent-insoluble complexes containing the familial Alzheimer's disease-associated presenilins. J Neurochem 72: 1534–1543.
Pennington K, Beasley CL, Dicker P, Fagan A, English J, Pariante CM et al (2008). Prominent synaptic and metabolic abnormalities revealed by proteomic analysis of the dorsolateral prefrontal cortex in schizophrenia and bipolar disorder. Mol Psychiatry 13: 1102–1117.
Perry EK, Gibson PH, Blessed G, Perry RH, Tomlinson BE (1977). Neurotransmitter enzyme abnormalities in senile dementia: Choline acetyltransferase and glutamic acid decarboxylase activities in necropsy brain tissue. J Neurol Sci 34: 247–265.
Pierri JN, Volk CL, Auh S, Sampson A, Lewis DA (2001). Decreased somal size of deep layer 3 pyramidal neurons in the prefrontal cortex of subjects with schizophrenia. Arch Gen Psychiatry 58: 466–473.
Piette MA, Pieters SP, Letter E (2011). Evaluation of the agonal stress: can immunohistochemical detection of ubiquitin in the locus coeruleus be useful? Int J Legal Med 125: 333–340.
Prabakaran S, Swatton JE, Ryan MM, Huffaker SJ, Huang JT, Griffin JL et al (2004). Mitochondrial dysfunction in schizophrenia: evidence for compromised brain metabolism and oxidative stress. Mol Psychiatry 9: 684–697 643.
Rea TD (2005). Agonal respirations during cardiac arrest. Curr Opin Crit Care 11: 188–191.
Roberts RC (2007). Schizophrenia in translation: disrupted in schizophrenia (DISC1): integrating clinical and basic findings. Schizophr Bull 33: 11–15 Highlights the relevance of using postmortem tissues to extend and confirm hypothesis generated by preclinical studies.
Rubio MD, Haroutunian V, Meador-Woodruff JH (2012). Abnormalities of the Duo/Ras-related C3 botulinum toxin substrate 1/p21-activated kinase 1 pathway drive myosin light chain phosphorylation in frontal cortex in schizophrenia. Biol Psychiatry 71: 906–914.
Rumbaugh G, Sia G, Garner C, Huganir R (2003). Synapse-associated protein-97 isoform-specific regulation of surface AMPA receptors and synaptic function in cultured neurons. J Neurosci 23: 4567–4576.
Ruzicka WB, Zhubi A, Veldic M, Grayson DR, Costa E, Guidotti A (2007). Selective epigenetic alteration of layer I GABAergic neurons isolated from prefrontal cortex of schizophrenia patients using laser-assisted microdissection. Mol Psychiatry 12: 385–397.
Sawamura N, Sawamura-Yamamoto T, Ozeki Y, Ross CA, Sawa A (2005). A form of DISC1 enriched in nucleus: altered subcellular distribution in orbitofrontal cortex in psychosis and substance/alcohol abuse. Proc Natl Acad Sci USA 102: 1187–1192.
Schmitt A, Hasan A, Gruber O, Falkai P (2011). Schizophrenia as a disorder of disconnectivity. Eur Arch Psychiatry Clin Neurosci 261 (Suppl 2): S150–S154.
Scott DB, Blanpied TA, Ehlers MD (2003). Coordinated PKA and PKC phosphorylation suppresses RXR-mediated ER retention and regulates the surface delivery of NMDA receptors. Neuropharmacology 45: 755–767.
Shan D, Yates SD, Roberts RC, McCullumsmith RE (2013). Update on the neurobiology of schizophrenia: a role for extracellular microdomains. Minerva Psichiatrica 53: 233–249.
Sikkema AH, Diks SH, den Dunnen WF, ter Elst A, Scherpen FJ, Hoving EW et al (2009). Kinome profiling in pediatric brain tumors as a new approach for target discovery. Cancer Res 69: 5987–5995.
Smith MJ, Sharples RA, Evin G, McLean CA, Dean B, Pavey G et al (2004). Expression of truncated presenilin 2 splice variant in Alzheimer's disease, bipolar disorder, and schizophrenia brain cortex. Brain Res Mol Brain Res 127: 128–135.
Smoller JW, Craddock N, Kendler K, Lee PH, Neale BM, Nurnberger JI et al (2013). Identification of risk loci with shared effects on five major psychiatric disorders: a genome-wide analysis. Lancet 381: 1371–1379.
Sodhi MS, Simmons M, McCullumsmith R, Haroutunian V, Meador-Woodruff JH (2011). Glutamatergic gene expression is specifically reduced in thalamocortical projecting relay neurons in schizophrenia. Biol Psychiatry 70: 646–654 The first study to characterize and employ cell-level laser capture microdissection in postmortem tissue in severe mental illness.
Sokolov B (1998). Expression of NMDAR1, GluR1, GluR7, and KA1 glutamate receptor mRNAs is decreased in frontal cortex of "neuroleptic-free" schizophrenics: evidence on reversible up-regulation by typical neuroleptics. J Neurochem 71: 2454–2464.
Spokes EGS, Garrett NJ, Iversen LL (1979). Differential effects of agonal status on measurements of gaba and glutamate decarboxylase in human post-mortem brain tissue from control and Huntington's chorea subjects. J Neurochem 33: 773–778.
Stan AD, Ghose S, Gao XM, Roberts RC, Lewis-Amezcua K, Hatanpaa KJ et al (2006). Human postmortem tissue: what quality markers matter? Brain Res 1123: 1–11.
Stefansson H, Sigurdsson E, Steinthorsdottir V, Bjornsdottir S, Sigmundsson T, Ghosh S et al (2002). Neuregulin 1 and susceptibility to schizophrenia. Am J Hum Genet 71: 877–892.
Stockmeier CA (1997). Neurobiology of serotonin in depression and suicide. Ann NY Acad Sci 836: 220–232.
Swatton JE, Sellers LA, Faull RL, Holland A, Iritani S, Bahn S (2004). Increased MAP kinase activity in Alzheimer's and Down syndrome but not in schizophrenia human brain. Eur J Neurosci 19: 2711–2719.
Szeszko PR, Hodgkinson CA, Robinson DG, Derosse P, Bilder RM, Lencz T et al (2008). DISC1 is associated with prefrontal cortical gray matter and positive symptoms in schizophrenia. Biol Psychol 79: 103–110.
Szumlinski KK, Lominac KD, Kleschen MJ, Oleson EB, Dehoff MH, Schwarz MK et al (2005). Behavioral and neurochemical phenotyping of Homer1 mutant mice: possible relevance to schizophrenia. Genes Brain Behav 4: 273–288.
Takahashi N, Nielsen KS, Aleksic B, Petersen S, Ikeda M, Kushima I et al (2011). Loss of function studies in mice and genetic association link receptor protein tyrosine phosphatase alpha to schizophrenia. Biol Psychiatry 70: 626–635.
Takahashi T, Suzuki M, Tsunoda M, Maeno N, Kawasaki Y, Zhou SY et al (2009). The Disrupted-in-Schizophrenia-1 Ser704Cys polymorphism and brain morphology in schizophrenia. Psychiatry Res 172: 128–135.
Talbot K, Eidem WL, Tinsley CL, Benson MA, Thompson EW, Smith RJ et al (2004). Dysbindin-1 is reduced in intrinsic, glutamatergic terminals of the hippocampal formation in schizophrenia. J Clin Invest 113: 1353–1363.
Tamminga CA, Dale JM, Goodman L, Kaneda H, Kaneda N (1990). Neuroleptic-induced vacuous chewing movements as an animal model of tardive dyskinesia: a study in three rat strains. Psychopharmacology (Berl) 102: 474–478.
Tamminga CA, Thaker GK, Moran M, Kakigi T, Gao XM (1994). Clozapine in tardive dyskinesia: observations from human and animal model studies. J Clin Psychiatry 55 (Suppl B): 102–106.
Tomita H, Vawter MP, Walsh DM, Evans SJ, Choudary PV, Li J et al (2004a). Effect of agonal and postmortem factors on gene expression profile: quality control in microarray analyses of postmortem human brain. Biol Psychiatry 55: 346–352.
Tomita Y, Nihira M, Ohno Y, Sato S (2004b). Ultrastructural changes during in situ early postmortem autolysis in kidney, pancreas, liver, heart and skeletal muscle of rats. Leg Med (Tokyo) 6: 25–31.
Toro C, Deakin JF (2005). NMDA receptor subunit NRI and postsynaptic protein PSD-95 in hippocampus and orbitofrontal cortex in schizophrenia and mood disorder. Schizophr Res 80: 323–330.
Toyooka K, Iritani S, Makifuchi T, Shirakawa O, Kitamura N, Maeda K et al (2002a). Selective reduction of a PDZ protein, SAP-97, in the prefrontal cortex of patients with chronic schizophrenia. J Neurochem 83: 797–806.
Toyooka K, Usui M, Washiyama K, Kumanishi T, Takahashi Y (2002b). Gene expression profiles in the brain from phencyclidine-treated mouse by using DNA microarray. Ann NY Acad Sci 965: 10–20.
Trinh JV, Nehrenberg DL, Jacobsen JP, Caron MG, Wetsel WC (2003). Differential psychostimulant-induced activation of neural circuits in dopamine transporter knockout and wild type mice. Neuroscience 118: 297–310.
Trotter SA, Brill LB 2nd, Bennett JP Jr. (2002). Stability of gene expression in postmortem brain revealed by cDNA gene array analysis. Brain Res 942: 120–123.
Tsai G, Passani LA, Slusher BS, Carter R, Baer L, Kleinman JE et al (1995). Abnormal excitatory neurotransmitter metabolism in schizophrenic brains. Arch Gen Psychiatry 52: 829–836.
Tsai G, Ralph-Williams RJ, Martina M, Bergeron R, Berger-Sweeney J, Dunham KS et al (2004). Gene knockout of glycine transporter 1: characterization of the behavioral phenotype. Proc Natl Acad Sci USA 101: 8485–8490.
Tucholski J, Simmons MS, Pinner AL, Haroutunian V, McCullumsmith RE, Meador-Woodruff JH (2013). Abnormal N-linked glycosylation of cortical AMPA receptor subunits in schizophrenia. Schizophr Res 146: 177–183.
Tuominen H, Tiihonen J, Wahlbeck K (2005). Glutamatergic drugs for schizophrenia: a systematic review and meta-analysis. Schizophr Res 72: 225–234.
Tuominen H, Tiihonen J, Wahlbeck K (2006). Glutamatergic drugs for schizophrenia. Cochrane Database Syst Rev 2: 1–40.
Uezato A, Meador-Woodruff JH, McCullumsmith RE (2009). Vesicular glutamate transporter mRNA expression in the medial temporal lobe in major depressive disorder, bipolar disorder, and schizophrenia. Bipolar Disord 11: 711–725.
Valtschanoff JG, Burette A, Davare MA, Leonard AS, Hell JW, Weinberg RJ (2000). SAP97 concentrates at the postsynaptic density in cerebral cortex. Eur J Neurosci 12: 3605–3614.
Vawter MP, Barrett T, Cheadle C, Sokolov BP, Wood WH 3rd, Donovan DM et al (2001). Application of cDNA microarrays to examine gene expression differences in schizophrenia. Brain Res Bull 55: 641–650.
Vawter MP, Tomita H, Meng F, Bolstad B, Li J, Evans S et al (2006). Mitochondrial-related gene expression changes are sensitive to agonal-pH state: implications for brain disorders. Mol Psychiatry 11: 663–679.
Walss-Bass C, Liu W, Lew DF, Villegas R, Montero P, Dassori A et al (2006). A novel missense mutation in the transmembrane domain of neuregulin 1 is associated with schizophrenia. Biol Psychiatry 60: 548–553 Establishes that the multipotent neuregulin1 gene has a causative role in schizophrenia.
Webster MJ (2006). Tissue preparation and banking. Prog Brain Res 158: 3–14.
Weickert CS, Sheedy D, Rothmond DA, Dedova I, Fung S, Garrick T et al (2010). Selection of reference gene expression in a schizophrenia brain cohort. Aust NZ J Psychiatry 44: 59–70.
Weickert CS, Straub RE, McClintock BW, Matsumoto M, Hashimoto R, Hyde TM et al (2004). Human dysbindin (DTNBP1) gene expression in normal brain and in schizophrenic prefrontal cortex and midbrain. Arch Gen Psychiatry 61: 544–555.
Weiss IC, Di Iorio L, Feldon J, Domeney AM (2000). Strain differences in the isolation-induced effects on prepulse inhibition of the acoustic startle response and on locomotor activity. Behav Neurosci 114: 364–373.
Wezenberg E, Verkes R, Ruigt G, Hulstijn W, Sabbe B (2007). Acute effects of the ampakine farampator on memory and information processing in healthy elderly volunteers. Neuropsychopharmacology 32: 1272–1283.
Whittaker VP, Michaelson IA, Kirkland RJ (1963). The separation of synaptic vesicles from disrupted nervending particles. Biochem Pharmacol 12: 300–302.
Wible CG, Preus AP, Hashimoto R (2009). A cognitive neuroscience view of schizophrenic symptoms: abnormal activation of a system for social perception and communication. Brain Imaging Behav 3: 85–110.
Wu EQ, Birnbaum HG, Shi L, Ball DE, Kessler RC, Moulis M et al (2005). The economic burden of schizophrenia in the United States in 2002. J Clin Psychiatry 66: 1122–1129.
Yao W-D, Gainetdinov RR, Arbuckle MI, Sotnikova TD, Cyr M, Beaulieu J-M et al (2004). Identification of PSD-95 as a regulator of dopamine-mediated synaptic and behavioral plasticity. Neuron 41: 625–638.
Yee BK, Keist R, von Boehmer L, Studer R, Benke D, Hagenbuch N et al (2005). A schizophrenia-related sensorimotor deficit links alpha 3-containing GABAA receptors to a dopamine hyperfunction. Proc Natl Acad Sci USA 102: 17154–17159.
Yoo BC, Seidl R, Cairns N, Lubec G (1999). Heat-shock protein 70 levels in brain of patients with Down syndrome and Alzheimer's disease. J Neural Transm Suppl 57: 315–322.
Zhang L, Daly RJ (2012). Targeting the human kinome for cancer therapy: current perspectives. Crit Rev Oncog 17: 233–246.
Zhang Q, Fan J, Zhang M (2001). Interdomain chaperoning between PSD-95, Dlg, and Zo-1 (PDZ) domains of glutamate receptor-interacting proteins. J Biol Chem 276: 43216–43220 Characterizes the role(s) of PDZ domains for glutamate receptor trafficking.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
McCullumsmith, R., Hammond, J., Shan, D. et al. Postmortem Brain: An Underutilized Substrate for Studying Severe Mental Illness. Neuropsychopharmacol 39, 65–87 (2014). https://doi.org/10.1038/npp.2013.239
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2013.239
Keywords
This article is cited by
-
The synaptic hypothesis of schizophrenia version III: a master mechanism
Molecular Psychiatry (2023)
-
Identification of novel targets and pathways to distinguish suicide dependent or independent on depression diagnosis
Scientific Reports (2023)
-
Isolation and Differentiation of Neurons and Glial Cells from Olfactory Epithelium in Living Subjects
Molecular Neurobiology (2023)
-
Glutamatergic dysfunction in Schizophrenia
Translational Psychiatry (2022)
-
A qualitative study exploring the process of postmortem brain tissue donation after suicide
Scientific Reports (2022)
