
Abstract
Approximately 30–40% of estrogen receptor α (ERα)-positive breast tumors express high levels of the cyclooxygenase-2 (COX-2) protein, and these high levels have been associated with a poorer prognosis in breast cancer patients. We speculate that high levels of COX-2 induce drug resistance in ERα-positive breast tumors, thus reducing the survival rate of patients with such tumors. Human breast cancer cell lines that express high levels of COX-2 are generally ERα negative. To determine whether COX-2 induces drug resistance, plasmids encoding the COX-2 gene were stably transfected into ERα-positive MCF-7 human breast cancer cells (MCF-7/COX-2). MCF-7/COX-2 cells were resistant to the selective estrogen receptor modulator tamoxifen but not to its analog, raloxifene. MCF-7/COX-2 cells were also resistant to the retinoid N-(4-hydroxyphenyl)retinamide (4-HPR) but not to its analog, all-trans retinoic acid. In contrast, the sensitivities of MCF-7/COX-2 cells to doxorubicin and paclitaxel were similar to those of the parental MCF-7 cells. We then determined which COX-2 product, prostaglandin E2 (PGE2) or prostaglandin F2α is involved in the COX-2-mediated drug resistance. PGE2, but not PGF2α, blocked the antiproliferative effects of tamoxifen and 4-HPR. Agonists that activate PGE2 receptors and their downstream kinase effectors, protein kinases A and C, also blocked the growth inhibitory effects of these drugs. Increased levels of Bcl-2 and Bcl-XL proteins have been reported in mammary tumors of COX-2 transgenic mice and in human colon cancer cell lines that have high levels of COX-2. However, we did not observe any changes in Bcl-2, Bcl-XL, or Bax expression induced by COX-2 or PGE2. Here we report the novel findings that COX-2 uses PGE2 to stimulate the activities of protein kinases A and C to induce selectively tamoxifen and 4-HPR resistance in ERα-positive breast cancer cells.
Similar content being viewed by others

Main
The cyclooxygenase-2 (COX-2) protein is the inducible form of the COX enzymes. Functionally, the COX-2-derived prostaglandins (PGs) have been shown to block apoptosis, induce invasion, and promote angiogenesis. High levels of COX-2 and its products prostaglandin E2 (PGE2) and prostaglandin F2α (PGF2α) have been found in human mammary tumor tissues.1, 2, 3, 4, 5, 6, 7, 8, 9 Elevated COX-2 expression had been associated with breast tumors of an estrogen receptor α (ERα)-negative status, large tumor size, high histological grade, high proliferation rate, high p53 expression, and the presence of the amplification of the HER2/neu oncogene.2, 4, 7 High expression of COX-2 was associated with reduced disease-free survival in breast cancer patients with ERα-negative tumors.10
High levels of COX-2 have been identified in 30–40% of ERα-positive breast tumors,2 and been associated with the development of ERα-positive tumors. Terry et al11 showed that women who were newly diagnosed with in situ or invasive breast cancer had reduced breast cancer risk when they used aspirin at least once per week for 6 months or longer. The reduction in breast cancer risk was seen among women with ERα-positive, but not ERα-negative, breast cancers.11 Reduction in breast cancer risk was also seen in women who used ibuprofen but not acetaminophen. Since aspirin and ibuprofen, not acetaminophen, can inhibit COX-2 activity, it was speculated that among the numerous mechanisms of these two drugs, it is their anti-COX-2 activities that reduce breast cancer risk. In a retrospective study of 1576 invasive breast tumors, Ristimaki et al2 found that elevated COX-2 expression was associated with a lower survival rate in patients with ERα-positive breast tumors. This association was particularly significant for ERα-positive, COX-2-overexpressing breast tumors that did not have p53 expression or HER2/neu amplification.2 Women whose invasive breast tumors were ERα positive but had low levels of COX-2 had an 86% chance of 5-year distant disease-free survival, whereas women whose tumors were ERα positive but had high levels of COX-2 had a 76% chance of 5-year distant disease-free survival.2
Breast cancer patients who have ERα-positive breast tumors are typically treated with selective estrogen receptor modulators (SERMs) and/or chemotherapy. We speculate that high levels of the COX-2 protein cause ERα-positive breast tumors to be resistant to SERMs and/or chemotherapy, thus reducing the survival rate of patients with these tumors. Human breast cancer cell lines that express high levels of COX-2 are generally ERα negative, or generally contain HER2/neu gene amplification even if ERα positive. To determine whether high levels of COX-2 could affect drug sensitivity of ERα-positive breast cancer cells, stable transfection of plasmids encoding the COX-2 gene into ERα-positive MCF-7 human breast cancer cells, which do not have HER2/neu gene amplification, was performed. We evaluated whether COX-2 overexpression causes MCF-7 cells to be resistant to the SERMs tamoxifen and raloxifene, and to the chemotherapeutic drugs doxorubicin and paclitaxel. We also evaluated whether COX-2 overexpression causes breast cancer cells to be resistant to the potential chemopreventive agents, all-trans retinoic acid (ATRA) and N-(4-hydroxyphenyl) retinamide (4-HPR). Finally, we evaluated whether PGE2 and PGF2α are involved in COX-2-induced resistance.
Materials and methods
Reagents
Tamoxifen citrate, raloxifene, ATRA, and 4-HPR were purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA). PGE2, PGF2α, 17-phenyl trinor PGE2 (an EP1 receptor agonist), and butaprost (an EP2 receptor agonist) were purchased from Cayman Chemical (Ann Arbor, MI, USA). Forskolin and phorbol myristate acetate (PMA) were purchased from EMD Biosciences (La Jolla, CA, USA). Doxorubicin and paclitaxel were purchased from our institutional pharmacy. Stock solutions (10 mM) of tamoxifen, raloxifene, ATRA, 4-HPR, PGE2, PGF2α, EP1 receptor agonist, EP2 receptor agonist, forkskolin, and PMA were prepared in DMSO and stored at –20°C. All reagents were diluted in culture medium to the indicated final concentration.
Cell Lines and Culture Conditions
MCF-7 human breast cancer cells were obtained from American Type Culture Collection (Manassas, VA, USA). The cells were cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 medium supplemented with 5% heat-inactivated fetal bovine serum (FBS) at 37°C under 5% CO2 in a humidified incubator. MCF-7/HER2 cells, generously provided by Dr Mien-Chie Hung of our institution, were grown in DMEM/F12 medium containing 5% FBS and 500 μg/ml G418.
COX-2 cDNA Transfection
The pSG5-COX-2 plasmid, which contains a full-length human COX-2 cDNA in the pSG expression vector,12 was used for COX-2 transfection. MCF-7 cells were plated at 1 × 105 cells/well in six-well plates in DMEM/F12 medium supplemented with 5% FBS. After overnight attachment, the medium was changed to OpTI-MEM*I reduced serum medium (Invitrogen, Carlsbad, CA, USA). FuGENE 6 (Roche Diagnostics Corporation, Indianapolis, IN, USA) was mixed with DNA plasmids (1 μg pcDNA3 vector to 5 μg pSG5-COX-2 plasmid) at a ratio of 3 μl/μg DNA for 20 min before being added to cell cultures. After 5 h of transfection, the medium was changed to DMEM/F12 medium. After 48 h, G418 was added to cells at a concentration of 500 μg/ml. Cell lines were obtained from individual colonies using cloning cylinders. Seven vector (MCF-7/V) and 13 COX-2-overexpressing (MCF-7/COX-2) clones were obtained. MCF-7/V and MCF-7/COX-2 cells were continuously cultured in DMEM/F12 medium containing 5% FBS and 500 μg/ml G418.
Celltiter 96 Aqueous Nonradioactive Proliferation Assay
To determine the effects of COX-2 overexpression on the sensitivity of breast cancer cells to tamoxifen and raloxifene, breast cancer cells were plated at 1000 cells/well in 96-well plates in 0.1 ml of DMEM/F12 medium supplemented with 5% FBS. The next day, the medium was changed to DMEM/F12 medium supplemented with 5% charcoal-stripped serum (CSS). After 24 h, cells were treated with various concentrations of tamoxifen or raloxifene for 5 days. At the end of the incubation, cell proliferation was determined by the Promega (Madison, WI, USA) Celltiter 96 Aqueous nonradioactive proliferation (MTS) assay and was expressed as the percentage of proliferating cells relative to untreated cells.
For experiments involving doxorubicin, paclitaxel, ATRA, and 4-HPR, breast cancer cells were plated at 1000 cells/well in 96-well plates in 0.1 ml of DMEM/F12 medium supplemented with 5% FBS. The next day, the cells were treated with various concentrations of doxorubicin, paclitaxel, ATRA, or 4-HPR for 5 days. At the end of the incubation, cell proliferation was determined by the MTS assay and was expressed as the percentage of proliferating cells relative to untreated cells.
Cell Counting
The effects of PGE2 and PGF2α on tamoxifen sensitivity were determined. Parental MCF-7 (MCF-7/WT) cells were plated at 1.0 × 105 cells/well in six-well plates in 2 ml of DMEM/F12 medium supplemented with 5% FBS. The next day, the medium was changed to DMEM/F12 medium supplemented with 5% CSS. After 24 h, cells were treated with tamoxifen (1 μM) in the presence or absence of exogenous PGE2 or PGF2α (10 μM) for 5 days. At the end of the incubation, the number of live cells was counted using Trypan blue exclusion.
The effects of PGE2 and PGF2α on 4-HPR sensitivity of MCF-7/WT cells were determined in DMEM/F12 medium supplemented with 5% FBS. MCF-7/WT cells were plated at 0.5 × 105 cells/well in six-well plates and were incubated with 4-HPR (1 μM) in the presence and absence of exogenous PGE2 or PGF2α (10 μM). After 5 days of incubation, the number of live cells was counted using Trypan blue exclusion.
Western Blot
Protein lysates (50 μg) from untreated exponentially growing breast cancer cells were loaded on 12% polyacrylamide gels to determine the COX-2 status. Proteins were electrophoresed and electrotransferred as described by Simeone et al.13 Membranes were incubated with mouse monoclonal COX-2 antibody (Cayman Chemical). β-Actin (Sigma Chemical Co.) was used as a loading control. Protein bands were visualized by enhanced chemiluminescence (Kirkgaard & Perry Laboratories, Gaithersburg, MD, USA). Images were scanned using an Alpha Imager application program (Alpha Innotech, San Leadro, CA, USA).
To determine the effects of PGE2 and PGF2α on the expression levels of the Bcl-2, Bcl-XL, and Bax proteins, MCF-7/WT cells were plated at 0.5 × 105 cells/well in six-well plates in 2 ml of DMEM/F12 medium supplemented with 5% FBS. After 24 h, cells were treated with PGE2 or PGF2α (10 μM). After 4 days of incubation, cells were harvested, and Western blotting was performed. Membranes were incubated with mouse monoclonal Bcl-2, Bcl-XL, and Bax antibodies (Cell Signaling, Beverly, MA, USA).
Flow Cytometric Assay
The effects of tamoxifen or 4-HPR on MCF-7/WT and MCF-7/COX-2 cells were analyzed by flow cytometry as described by Simeone et al.13 Approximately 1 × 106 breast cancer cells were trypsinized, collected by centrifugation at 1500 r.p.m. for 5 min, washed in PBS, and resuspended in 1 ml of PBS. The cell suspension was added to 1 ml of cold 70% ethanol and incubated overnight at −20°C. Cells were centrifuged at 1500 r.p.m. for 10 min at 4°C and then washed twice in PBS, and the pellet was left loose. Approximately 0.5–1 ml of PBS containing RNase (20 μg/ml) and propidium iodide (50 μg/ml) was added to each cell pellet, followed by 20 min of incubation at room temperature. Flow cytometric analysis was performed using a Coulter Epics Profile 488 laser. The percent values reported were the means of two experiments.
RNA Extraction
Total RNA was extracted from breast cancer cells using a modified protocol of the RNeasy® Mini Kit (Qiagen Inc., Valencia, CA, USA). Briefly, the cells were exposed to 0.05% trypsin for dislodgement, and then centrifuged to remove trypsin. Cell pellets were mixed with 350 μl of RLT lysis buffer and 350 μl of 70% ethanol. The cell mixture was homogenized by passing through 28.5-G needles with 0.5 ml syringe 10–12 times, and then passed through the RNeasy® Mini filter cartridges. Filter cartridges were washed once with 700 μl of buffer RW1, twice with 500 μl of buffer RPE, and the filtrate from these washes was discarded. After eluting twice into a fresh collection tube with 40 μl of RNase-free water, the RNA was stored at −80°C. RNA concentration was measured by NanoDrop® ND-1000 UV-Vis spectrophotometer (NanoDrop Technologies Inc., Rockland, DE, USA).
Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
RT-PCR was performed using an ABI 7900 sequence detector (Applied Biosystems, Foster City, CA, USA). The specific primers and probes for each gene were purchased through Assays-on-Demand from Applied Biosystems. cDNA was synthesized using Superscript II reverse transcriptase (Invitrogen Life Technology). Synthesized cDNAs were mixed with primers/probes in the 2 × Taqman universal PCR buffer, and then assayed on an ABI 7900. The data obtained from assays were analyzed with SDS 2.1 software (Applied Biosystems). The amount of total RNA of each sample was normalized to GAPDH transcript levels.
Results
COX-2 Expression in MCF-7 Breast Cancer Cells
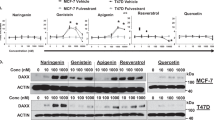
In all, 13 clones of MCF-7/COX-2 cells were found. Western blotting was used to confirm that these clones have higher levels of COX-2 than the parental MCF-7/WT cells. As expected, different MCF-7/COX-2 clones expressed the COX-2 protein at different levels (Figure 1). Clones 13 and 12 expressed higher levels of COX-2 than clones 5 and 1. Compared to the parental MCF-7/WT cells, MCF-7/COX-2 clones 13, 12, 5 and 1 expressed 3.6-, 3.1-, 2.1-, and 1.4-fold higher levels of COX-2, respectively. Clones 12 and 13 were used for the subsequent experiments. The levels of COX-2 were similar between the vector-transfected (MCF-7/V) and the MCF-7WT cells (Figure 1).

COX-2 expression in MCF-7 cells. Western blots were carried out to demonstrate whether higher levels of COX-2 protein are expressed in MCF-7/COX-2 clones than in MCF-7/WT and MCF-7/V cells. In all, 50 μg of proteins were obtained from exponentially growing cells and were loaded on 12% SDS-PAGE, and Western blots using monoclonal COX-2 antibodies were done. β-Actin was used as a loading control.
COX-2 Selectively Suppresses the Inhibitory Effects of Tamoxifen and 4-HPR
The inhibitory effects of tamoxifen were determined in MCF-7/WT and MCF-7/COX-2 cells. MCF-7/WT cells were sensitive to tamoxifen. In the presence of 5% CSS, the IC50 value of tamoxifen in MCF-7/WT cells was approximately 1 μM (Figure 2a). On the other hand, the IC50 values of tamoxifen in MCF-7/COX-2 clones 12 and 13 were >5 μM (Figure 2a).

COX-2 selectively induces resistance to tamoxifen and 4-HPR in MCF-7 cells. MCF-7/WT (□), MCF-7/COX-2 clone 12 (▴), and MCF-7/COX-2 clone 13 (▪) cells were plated at 1000 cells/well in 96-well plates in DMEM/F12 medium containing 5% FBS. The next day, the medium was changed to DMEM/F12 medium containing 5% CSS. Cells were treated with 0–9 μM of (a) tamoxifen or (b) raloxifene for 5 days. An MTS assay was carried out to determine the proliferation of tamoxifen- and raoloxifene-treated cells. Values represent the means of five wells±s.d. and are expressed as percentages relative to untreated cells. Statistical analysis of tamoxifen-induced inhibitory effects induced in MCF-7/WT cells and MCF-7/COX-2 clones 12 and 13 was performed using a two-tailed paired t-test (*P<0.001). After overnight attachment, cells were treated with (c) 0–5 μM retinoids (4-HPR, filled lines; ATRA, dashed lines) or (d) 0–100 ng/ml doxorubicin for 5 days. MCF-7/HER2 (⋄) cells were used as a doxorubicin-resistant control. An MTS assay was carried out to determine the proliferation of retinoid- and doxorubicin-treated cells. Values represent the means of three wells±s.d. and are expressed as percentages relative to untreated cells. Statistical analysis of 4-HPR-induced inhibitory effects induced in MCF-7/WT and MCF-7/COX-2 clone 12 cells were performed using a two-tailed paired t-test (*P<0.001).
Since COX-2 was found to decrease the sensitivity of MCF-7 breast cancer cells to tamoxifen, the effects of COX-2 on another SERM, raloxifene, were studied. Contrary to tamoxifen, raloxifene was equally effective in inducing growth inhibition in MCF-7/WT and MCF-7/COX-2 cells (Figure 2b). Similar data were obtained when the experiments were performed in the presence of 5% FBS (data not shown). Thus, COX-2 suppresses the effects of tamoxifen, but not raloxifene, in breast cancer cells.
We previously demonstrated that the HER2/neu oncogene uses COX-2 to suppress the apoptotic effects of the retinoid 4-HPR.13 However, whether COX-2 by itself can suppress 4-HPR's effects is not known. Thus, we determined the proliferation of COX-2-overexpressing cells treated with 4-HPR. We also determined whether COX-2 affects the sensitivity of ATRA, a 4-HPR analog. MCF-7/COX-2 clone 12 cells were resistant to 4-HPR (Figure 2c). At a 1-μM concentration, 4-HPR reduced the proliferation of MCF-7/WT cells by 71% (Figure 2c), whereas the same concentration of 4-HPR reduced the proliferation of MCF-7/COX-2 cells by only 9% (Figure 2c). However, MCF-7/COX-2 clone 12 cells were sensitive to ATRA. At a 1-μM concentration, ATRA reduced the proliferation of MCF-7/WT cells by 59% (Figure 2c). Under the same conditions, the proliferation of MCF-7/COX-2 cells was reduced by 42%, which was similar to that in MCF-7/WT cells (Figure 2c). These data indicate that COX-2 induces 4-HPR, but not ATRA, resistance in breast cancer cells.
The inhibitory effects of doxorubicin were also determined. MCF-7/HER2 cells were used as a positive control. Doxorubicin decreased the proliferation of MCF-7/WT and MCF-7/COX-2 clone 12 cells to about the same extent (Figure 2d). At a concentration of 2.5 μg/ml, doxorubicin decreased MCF-7/WT and MCF-7/COX-2 cell proliferation by 63 and 74%, respectively (Figure 2d), whereas under the same conditions, doxorubicin decreased the viability of MCF-7/HER2 cells by only 23% (Figure 2d). The sensitivity of MCF-7/COX-2 clone 12 cells to paclitaxel was similar to that of MCF-7/WT cells (data not shown). These data indicate that COX-2 does not affect the sensitivities of MCF-7 cells to doxorubicin or paclitaxel.
COX-2 Suppresses the Cell Cycle-Arresting Effects of Tamoxifen and the Apoptotic Effects of 4-HPR
Tamoxifen and 4-HPR have been shown to induce cell cycle arrest and apoptosis in breast cancer cells.13, 14 Flow cytometry was used to determine whether COX-2 disrupts the growth arrest or the apoptotic effects of these drugs. In the presence of 5% CSS, a 1-μM concentration of tamoxifen inhibits cell proliferation by arresting cells at the G1 phase. Tamoxifen increases the percentage of MCF-7/WT cells in the G1 phase from 66.7% (untreated cells) to 80.6%, and decreases the percentage of cells in the S phase from 21.3% (untreated cells) to 10.3% (Figure 3, top panel). However, the same concentration of tamoxifen was ineffective in arresting MCF-7/COX-2 clone 12 cells in the G1 phase. The percentages of MCF-7/COX-2 cells in the G1 phase were 60.0 and 57.7% for untreated and tamoxifen-treated cells, respectively; the percentages of MCF-7/COX-2 cells in the S phase were 32.4 and 29.4%, respectively (Figure 3, top panel). COX-2 suppresses the inhibitory effects of tamoxifen by preventing it from inducing G1 cell cycle arrest.

COX-2 blocks the G1-arresting effects of tamoxifen and the apoptotic effects of 4-HPR. MCF-7/WT and MCF-7/COX-2 cells were plated between 1 and 3 × 105 cells in T-25 flasks in DMEM/F12 medium containing 5% FBS. Cells were treated with (top) 1 μM of tamoxifen in 5% CSS or (bottom) 1 μM of 4-HPR in 5% FBS. After a 4-day incubation, cells were harvested and processed for propidium iodide staining, followed by flow cytometry.
Flow cytometry showed that 4-HPR, at a 2.5-μM concentration, increased the percentage of apoptotic cells from 1.0% (untreated cells) to 56.9% in MCF-7/WT cells (Figure 3, bottom panel). However, 4-HPR increased the percentage of apoptotic cells from 1.3% to only 10.5% in MCF-7/COX-2 cells (Figure 3, bottom panel). Thus, COX-2 suppresses the apoptotic effects of 4-HPR.
Effects of COX-2 on the RNA Levels of ER Target Genes
Previously we showed that the Bcl-2 and the carbonic anhydrase XII (CA12) genes are regulated by ER.15, 16 We used quantitative RT-PCR to determine whether overexpression of COX-2 affects the expression of these ER target genes. The Bcl-2 RNA levels are very similar between the untreated MCF-7/WT and MCF-7/COX-2 cells (Table 1). After incubating with 1-μM concentration of tamoxifen for 5 days, the Bcl-2 levels in MCF-7/WT and MCF-7/COX-2 cells were decreased to a similar extent (Table 1). On the other hand, the RNA levels of the CA12 gene were about two-fold higher in MCF-7/COX-2 than MCF-7/WT cells (Table 1). Tamoxifen was able to decrease the CA12 levels by about 50–60% in MCF-7/WT and MCF-7/COX-2 cells (Table 1). Thus, COX-2 could affect the basal levels of some ER target genes.
PGE2, but not PGF2α, Suppresses Tamoxifen- and 4-HPR-Induced Growth Inhibition
Both PGE2 and PGF2α have been found in high levels in human breast tumor samples8, 9 and are implicated in COX-2-mediated mammary tumorigenesis. PGE2 suppressed the inhibitory effects of tamoxifen in MCF-7/WT cells. When MCF-7/WT cells were incubated with 1 μM tamoxifen in 5% CSS, cell numbers were reduced by 49% (Figure 4a). In the presence of 10 μM PGE2, tamoxifen only reduced cell numbers by 6% (Figure 4a). However, in the presence of PGF2α, tamoxifen was equally effective in decreasing cell counts (Figure 4a). 4-HPR reduced cell numbers by 55 and 20% in the absence and presence of PGE2, respectively (Figure 4b). However, 4-HPR was equally effective in decreasing cell counts in the presence of PGF2α (Figure 4b). These data indicate that PGE2, but not PGF2α, mediates the suppressive effects of COX-2 on tamoxifen and 4-HPR.

PGE2, but not PGF2α, suppresses the effects of tamoxifen and 4-HPR. MCF-7/WT cells were plated between 0.5 and 1.0 × 105 cells/well in six-well plates. Cells were treated with (a) 1 μM of tamoxifen for 5 days in 5% CSS, or (b) 1 μM of 4-HPR for 5 days in 5% FBS. Exogenous PGE2 or PGF2α at a 10-μM concentration was added to some wells. Cells were counted with the aid of a hemacytometer and Trypan blue dye. Values represent the means of three wells±s.d. Statistical analysis of tamoxifen- or 4-HPR-induced inhibitory effects in the presence and absence of PGE2 was performed using a two-tailed paired t-test (*P<0.001).
PGE2 does not Affect Levels of Bcl-2, Bcl-XL and Bax Proteins
Deregulation of Bcl-2, Bcl-XL, and Bax proteins has been reported in the mammary tumors of COX-2 transgenic mice17 and in colon cancer cells that have high levels of COX-2.12 Thus, we determined whether PGE2 suppresses the growth inhibitory effects of tamoxifen and 4-HPR by deregulating the expression of these proteins. Neither PGE2 nor PGF2α at 10-μM concentrations affected the levels of Bcl-2, Bcl-XL, or Bax proteins (Figure 5). The expression levels of these proteins also did not differ between MCF-7/WT and MCF-7/COX-2 cells (data not shown).

PGE2 does not change the levels of Bcl-2, Bcl-XL, or Bax proteins in MCF-7 cells. MCF-7/WT cells were treated with 10 μM PGE2 or PGF2α for 4 days. Cells were harvested, and 40 μg of proteins were loaded on a 15% SDS-PAGE. Western blots were done to compare the levels of Bcl-2, Bcl-XL, and Bax proteins in treated and untreated MCF-7 cells. β-Actin was used as a loading control.
COX-2 Uses Protein Kinases A and C to Suppress the Inhibitory Effects of Tamoxifen and 4-HPR
PGE2 exerts its effects through interaction with its cell surface receptors. We determined whether agonists of EP1 or EP2 receptors mediate the suppressive effects of COX-2 on tamoxifen and 4-HPR. The EP1 and the EP2 receptor agonists were not toxic to cells at 10-μM or lower concentrations (Figure 6a). In the absence of agonists, 1-μM concentration of tamoxifen reduced the proliferation of MCF-7/WT cells by 40–50% (Figure 6b). However, in the presence of the 10-μM concentration of EP1 and EP2 agonists, tamoxifen reduced MCF-7/WT cell proliferation by 15 and 9%, respectively (Figure 6b). Similarly, EP1 and EP2 agonists were also able to suppress the antiproliferative effects of 4-HPR (Figure 6c). A 1-μM concentration of 4-HPR reduced cell growth by 44%, but in the presence of EP1 and EP2 agonists, cell growth inhibition was 11 and 19%, respectively (Figure 6c).

Activation of EP1 receptor, EP2 receptor, PKA, or PKC decreases the sensitivity of MCF-7 cells to tamoxifen and 4-HPR. MCF-7/WT cells were plated at 1000 cells/well in 96-well plates. Cells were treated with (a) EP1 receptor agonist, EP2 receptor agonist, forskolin, or PMA alone. The concentrations used for EP1 receptor agonist, EP2 receptor agonist, and forskolin were: 0 μM (□), 1 μM (▨), 10 μM (▪), and 100 μM (⊞). The concentrations used for PMA were: 0 μM (□), 0.1 μM (░), 1 μM (▨), and 10 μM (▪). Cells were treated with (b) 1 μM of tamoxifen in 5% CSS, or (c) 1 μM of 4-HPR in 5% FBS in the presence and absence of the various agonists. EP1 receptor agonist, EP2 receptor agonist, and forskolin were used at: 0 μM (□), 1 μM (▨), and 10 μM (▪) concentrations. PMA was used at: 0 μM (□), 0.1 μM (░), and 1 μM (▨) concentrations. After a 5-day incubation, an MTS assay was carried out to determine the proliferation of tamoxifen- and 4-HPR-treated cells. Values represent the means of five wells ±s.d. and are expressed as percentages relative to untreated cells. Statistical analysis of tamoxifen- or 4-HPR-induced inhibitory effects in the presence and absence of agonists was performed using a two-tailed paired t-test (*P<0.001, +P<0.05).
Activation of EP1 and EP2 receptors is known to stimulate the activities of protein kinase A (PKA) and protein kinase C (PKC). Forskolin was used to stimulate PKA activity, and PMA was used to stimulate PKC activity. Forskolin was not toxic to cells at <10-μM concentration, and PMA was not toxic to cells at <1-μM concentration (Figure 6a). Both forskolin and PMA were able to block the inhibitory effects of tamoxifen (Figure 6b) and 4-HPR (Figure 6c). In the presence of forskolin (10 μM) and PMA (1 μM), a 1-μM concentration of tamoxifen reduced MCF-7/WT cell proliferation by 3 and 30%, respectively (Figure 6b). In the presence of forskolin and PMA, a 1-μM concentration of 4-HPR reduced cell growth by 21 and 6%, respectively (Figure 6c). These data indicate that, by stimulating PKA and PKC activities, EP1 and EP2 receptors are involved in the PGE2-induced resistance to tamoxifen and 4-HPR.
Discussion
Our data indicate that COX-2 selectively suppresses the antiproliferative effects of tamoxifen and 4-HPR in MCF-7 breast cancer cells. We previously demonstrated that the COX-2 protein is used by the HER2/neu oncogene to suppress 4-HPR-induced apoptosis.13 Here we show that, when stably transfected into ERα-positive MCF-7 breast cancer cells, the COX-2 protein by itself is sufficient to suppress 4-HPR-induced apoptosis. We also report the novel findings that COX-2 suppresses the growth inhibitory effects of tamoxifen in breast cancer cells. The effects of COX-2 are selective as the protein does not suppress the effects of raloxifene, ATRA, doxorubicin, or paclitaxel. Our data are in agreement with those of Mizutani et al,18 who found that COX-2 suppresses the effects of 5-fluorouracil, but not doxorubicin, in bladder cancer cell lines.
4-HPR is a synthetic retinamide analog of ATRA. However their mechanisms of action are very different. ATRA mediates its effects via retinoid receptor mechanisms,19 whereas 4-HPR mediates its effects via retinoid receptor-independent mechanisms.20, 21, 22, 23 We demonstrated that both retinoids can induce nitric oxide production, but nitric oxide is only essential for 4-HPR to induce its inhibitory effects.24 Both tamoxifen and raloxifene display antagonistic actions towards ERα and can inhibit the expression of ER target genes in breast cancer cells;15, 25 however, raloxifene is more effective than tamoxifen in promoting ER binding to the N-CoR corepressor.26 Microarray profiling demonstrated that tamoxifen and raloxifene induced distinct gene regulation patterns in MCF-7 breast cancer cells. In the endometrium, raloxifene acts as an antagonist but tamoxifen acts as an agonist and recruits coactivators instead of corepressors to ER target genes.27 The difference in the activities of these two SERMs may be due to their ability in inducing distinctly different ER–ligand complex conformations.28, 29
Increased COX-2 activity is known to increase the production of PGs, such as PGE2 and PGF2α. High levels of PGE2 and PGF2α have been found in human breast tumors and in transgenic mouse mammary glands that were engineered to produce high levels of COX-2.17, 30 Timoshenko et al31 showed that PGE2 is vital for breast cancer invasion, but they did not determine whether other PGs, such as PGF2α, are also involved. Previously we demonstrated that PGE2 is vital for HER2/neu to suppress 4-HPR-induced apoptosis; however, we did not determine whether other PGs were involved in suppressing 4-HPR-induced apoptosis.13 Here we show that it is PGE2, not PGF2α, which mediates the inhibitory effects of COX-2 on tamoxifen and 4-HPR. To our knowledge, this is the first report demonstrating the divergent role of these two PGs in breast cancer.
PGE2 exerts its effects through interaction with specific cell surface receptors. Here, we show that both EP1 receptors and EP2 receptors mediate PGE2 inhibitory effects on tamoxifen and 4-HPR. Activation of the EP1 receptor leads to increased intracellular Ca2+ levels and PKC activation, whereas activation of the EP2 receptor results in increased cAMP levels and PKA activation. To our knowledge, this is the first time that PKA and PKC activation are shown to be involved in 4-HPR resistance. Our results confirm previous reports that both PKA and PKC are involved in tamoxifen resistance. Stable transfection of PKCα in T-47D breast cancer cells resulted in tamoxifen-resistant tumor growth.32 Tonetti et al33 reported that PKCα overexpression is more frequent in primary tumors of patients who experience disease recurrence after tamoxifen treatment than in tumors of patients who remain disease free. Fujimoto and Katzenellenbogen34 showed that increasing the intracellular concentration of cAMP, or transfecting PKA catalytic subunits, leads to tamoxifen resistance in breast cancer cell lines. In clinical samples, Michalides et al35 found that downregulation of a negative regulator of PKA was associated with tamoxifen resistance, whereas Miller et al36, 37 correlated high levels of tumor camp-binding proteins with disease recurrence and poorer overall survival. However, it is not known what causes the deregulation of these two protein kinase pathways. We propose that COX-2, by activating EP1 and EP2 receptors via increased production of PGE2, is one link that could cause the deregulation of PKA and PKC signaling, and tamoxifen resistance in breast tumors.
We previously demonstrated that nitric oxide is essential for 4-HPR to induce apoptosis in breast cancer cells, as inhibitors of the nitric oxide synthases are able to block the 4-HPR effects.24 However, we do not believe nitric oxide is involved in the COX-2-mediated tamoxifen resistance because nitric oxide inhibitors were not able to suppress the growth inhibitory effects of tamoxifen (data not shown). Deregulated expression of the Bcl-2 family has been reported in COX-2 transgenic mice and in COX-2-overexpressing colon cancer cell lines.12, 17 However, we do not believe these proteins are involved in the COX-2-mediated resistance because the basal levels of Bcl-2, Bcl-XL, and Bax proteins were not affected by PGE2 treatment or COX-2 transfection. Bcl-2 is an ER target gene, but COX-2 did not affect the ability of tamoxifen to reduce Bcl-2 expression. COX-2 also did not affect the ability of tamoxifen to reduce the expression of CA12, another ER target gene. However, CA12 was expressed at higher levels in COX-2-overexpressing cells. One potential mechanism by which COX-2 induces tamoxifen resistance is by upregulating the basal levels of selective ER target genes so that the levels of these genes are still too high for tamoxifen to decrease and deliver its antiproliferative effects. We are currently using microarray and proteomic technologies to investigate what genes and proteins are up- or downregulated by COX-2. These technologies may enable us to further identify the downstream mechanisms used by COX-2/PGE2/PKA and PKC to induce tamoxifen and 4-HPR resistance.
COX-2 has emerged as a potential breast cancer therapeutic and chemopreventive target because it is expressed at high levels in breast tumors but not in normal breast tissues. Selective inhibitors of COX-2 have been shown to suppress carcinogen-induced and genetically induced breast cancer in animal models;38 these COX-2 inhibitors are currently being tested in clinical trials. Inhibitors of PGE2 production and/or its downstream signaling may also be useful for breast cancer therapy and chemoprevention. Indeed, a selective EP1 antagonist has been shown to reduce breast cancer incidence and multiplicity in a carcinogen-induced rat model.39 Elevated levels of COX-2 have been associated with the development of ERα-positive tumors.2, 11 Here, we demonstrate that COX-2 uses PGE2 to inhibit the antiproliferative and the apoptotic effects induced by tamoxifen and 4-HPR in ERα-positive breast cancer cells. These data suggest that breast cancer patients who have ERα-positive and COX-2-overexpressing tumors may not benefit from tamoxifen as much as patients who have low levels of COX-2 in their ERα-positive breast tumors. On the other hand, these patients may gain more clinical benefit from chemotherapy treatment or from raloxifene chemoprevention.
References
Half E, Tang XM, Gwyn K, et al. Cyclooxygenase-2 expression in human breast cancers and adjacent ductal carcinoma in situ. Cancer Res 2002;62:1676–1681.
Ristimaki A, Sivula A, Lundin J, et al. Prognostic significance of elevated cyclooxygenase-2 expression in breast cancer. Cancer Res 2002;62:63263–63265.
Shim V, Gauthier ML, Sudilovsky D, et al. Cyclooxygenase-2 expression is related to nuclear grade in ductal carcinoma in situ and is increased in its normal adjacent epithelium. Cancer Res 2003;63:2347–2350.
Boland GP, Butt IS, Prasad R, et al. COX-2 expression is associated with an aggressive phenotype in ductal carcinoma in situ. Br J Cancer 2004;90:423–429.
Davies G, Salter J, Hills M, et al. Correlation between cyclooxygenase-2 expression and angiogenesis in human breast cancer. Clin Cancer Res 2003;9:2651–2656.
Subbaramaiah K, Telang N, Ramonetti JT, et al. Transcription of cyclooxygenase-2 is enhanced in transformed mammary epithelial cells. Cancer Res 1996;56:4424–4429.
Denkert C, Winzer KJ, Muller BM, et al. Elevated expression of cyclooxygenase-2 is a negative prognostic factor for disease free survival and overall survival in patients with breast carcinoma. Cancer 2003;97:2978–2987.
Bennett A, Charlier EM, McDonald AM, et al. Prostaglandins and breast cancer. Lancet 1977;2:624–626.
Rolland P, Martin P, Jacquemier J, et al. Prostaglandin in human breast cancer: evidence suggesting that an elevated prostaglandin production is a marker of high metastatic potential for neoplastic cells. J Natl Cancer Inst 1980;64:1061–1070.
Witton CJ, Hawe SJ, Cooke TG, et al. Cyclooxygenase 2 (COX-2) expression is associated with poor outcome in ER-negative, but not ER-positive, breast cancer. Histopathology 2004;45:47–54.
Terry MB, Gammon MD, Zhang FF, et al. Association of frequency and duration of aspirin use and hormone receptor status with breast cancer risk. JAMA 2004;291:2433–2440.
Sun Y, Tang XM, Half E, et al. Cyclooxygenase-2 overexpression reduces apoptotic susceptibility by inhibiting the cytochrome c-dependent apoptotic pathway in human colon cancer cells. Cancer Res 2002;62:6323–6328.
Simeone AM, Li YJ, Broemeling LD, et al. Cyclooxygenase-2 is essential for HER2/neu to suppress N-(4-hydroxyphenyl)retinamide apoptotic effects in breast cancer cells. Cancer Res 2004;64:1224–1228.
Mandlekar S, Kong AN . Mechanisms of tamoxifen-induced apoptosis. Apoptosis 2001;6:469–477.
Zapata-Benavides P, Tuna M, Lopez-Berestein G, et al. Downregulation of Wilms' tumor 1 protein inhibits breast cancer proliferation. Biochem Biophys Res Commun 2002;295:784–790.
Pusztai L, Ayers M, Stec J, et al. Gene expression profiles obtained from fine-needle aspirations of breast cancer reliably identify routine prognostic markers and reveal large-scale molecular differences between estrogen-negative and estrogen-positive tumors. Clin Cancer Res 2003;9:2406–2415.
Liu C, Chang S, Narko K, et al. Overexpression of cyclooxygenase-2 is sufficient to induce tumorigenesis in transgenic mice. J Biol Chem 2001;267:18563–18569.
Mizutani Y, Kamoi K, Ukimura O, et al. Synergistic cytotoxicity and apoptosis of JTE-522, a selective cyclooxygenase-2 inhibitor, and 5-fluorouracil against bladder cancer. J Urol 2002;168:2650–2654.
Mangelsdorf D, Umesono K, Evans R . The retinoid receptors. In: Sporn M, Roberts A, Goodman D (eds). The Retinoids: Biology, Chemistry and Medicine, 2nd edn. Raven Press: New York, 1994, pp 319–350.
Sheikh M, Shao Z-M, Li X-S, et al. N-(4-hydroxyphenyl)retinamide (4-HPR)-mediated biological actions involve retinoid receptor-independent pathways in human breast carcinoma. Carcinogenesis 1995;16:2477–2486.
Fanjul A, Delia D, Pierotti M, et al. 4-Hydroxyphenyl retinamide is a highly selective activator of retinoid receptors. J Biol Chem 1996;271:22441–22446.
Kazmi S, Plante R, Visconti V, et al. Comparison of N-(4-hydroxyphenyl) retinamide and all-trans retinoic acid in the regulation of retinoid receptor-mediated gene expression in human breast cancer cell lines. Cancer Res 1996;56:1056–1062.
Dmitrovsky E . N-(4-hydroxyphenyl)retinamide activation of a distinct pathway signaling apoptosis. J Natl Cancer Inst 1997;89:1179–1181.
Simeone AM, Broemeling L, Rosenblum J, et al. Her2/neu reduces the sensitivity of breast cancer cells to N-(4-hydroxyphenyl)retinamide. Oncogene 2003;22:6739–6747.
Frasor J, Stossi F, Danes JM, et al. Selective estrogen receptor modulators: discrimination of agonistic versus antagonistic activities by gene expression profiling in breast cancer cells. Cancer Res 2004;64:1522–1533.
Webb P, Nguyen P, Kushner PJ . Differential SERM effects on corepressor binding dictate ERalpha activity in vivo. J Biol Chem 2003;278:6912–6920.
Shang Y, Brown M . Molecular determinants for the tissue specificity of SERMs. Science 2002;295:2465–2468.
Brzozowski AM, Pike AC, Dauter Z, et al. Molecular basis of agonism and antagonism in the oestrogen receptor. Nature 1997;389:753–758.
Shiau AK, Barstad D, Loria PM, et al. The structural basis of estrogen receptor/coactivator recognition and the antagonism of this interaction by tamoxifen. Cell 1998;95:927–937.
Chang SH, Liu CH, Conway R, et al. Role of prostaglandin E2-dependent angiogenic switch in cyclooxygenase 2-induced breast cancer progression. Proc Natl Acad Sci USA 2004;101:591–596.
Timoshenko AV, Xu G, Chakrabarti S, et al. Role of prostaglandin E2 receptors in migration of murine and human breast cancer cells. Exp Cell Res 2003;289:265–274.
Tonetti DA, Chisamore MJ, Grdina W, et al. Stable transfection of protein kinase C alpha cDNA in hormone-dependent breast cancer cell lines. Br J Cancer 2000;83:782–791.
Tonetti DA, Morrow M, Kidwai N, et al. Elevated protein kinase C alpha expression may be predictive of tamoxifen treatment failure. Br J Cancer 2003;88:1400–1402.
Fujimoto N, Katzenellenbogen BS . Alteration in the agonist/antagonist balance of antiestrogens by activation of protein kinase A signaling pathways in breast cancer cells: antiestrogen selectivity and promoter dependence. Mol Endocrinol 1994;8:296–304.
Michalides R, Griekspoor A, Balkenende A, et al. Tamoxifen resistance by a conformational arrest of the estrogen receptor alpha after PKA activation in breast cancer. Cancer Cell 2004;5:597–605.
Miller WR, Elton RA, Dixon JM, et al. Cyclic AMP binding proteins and prognosis in breast cancer. Br J Cancer 1990;61:263–266.
Miller WR, Watson DM, Jack W, et al. Tumour cyclic AMP binding proteins: an independent prognostic factor for disease recurrence and survival in breast cancer. Breast Cancer Res Treat 1993;26:89–94.
Howe L, Subbaramaiah K, Patel J, et al. Celecoxib, a selective cyclooxygenase 2 inhibitor, protects against human epidermal growth factor receptor 2 (HER-2)/neu-induced breast cancer. Cancer Res 2002;62:5405–5407.
Kawamori T, Uchiya N, Nakatsugi S, et al. Chemopreventive effects of ONO-8711, a selective prostaglandin E receptor EP(1) antagonist, on breast cancer development. Carcinogenesis 2001;22:2001–2004.
Acknowledgements
We thank Karen M Ramirez and The University of Texas MD Anderson Cancer Center, Department of Immunology Flow Cytometry Core Laboratory for their technical assistance. This research was supported in part by the instiutiton's NCI Core Grant (CA 16672), the Cancer Research and Prevention Foundation (to A-M Simeone) and the Susan G Komen Breast Cancer Foundation (to AM Tari).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tari, A., Simeone, AM., Li, YJ. et al. Cyclooxygenase-2 protein reduces tamoxifen and N-(4-hydroxyphenyl)retinamide inhibitory effects in breast cancer cells. Lab Invest 85, 1357–1367 (2005). https://doi.org/10.1038/labinvest.3700339
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.3700339
Keywords
This article is cited by
-
Antitumor activity of the selective cyclooxygenase-2 inhibitor, celecoxib, on breast cancer in Vitro and in Vivo
Cancer Cell International (2012)
-
Programmed Cell Death 4 inhibits breast cancer cell invasion by increasing Tissue Inhibitor of Metalloproteinases-2 expression
Breast Cancer Research and Treatment (2009)
-
TIMP-2 mediates the anti-invasive effects of the nitric oxide-releasing prodrug JS-K in breast cancer cells
Breast Cancer Research (2008)
