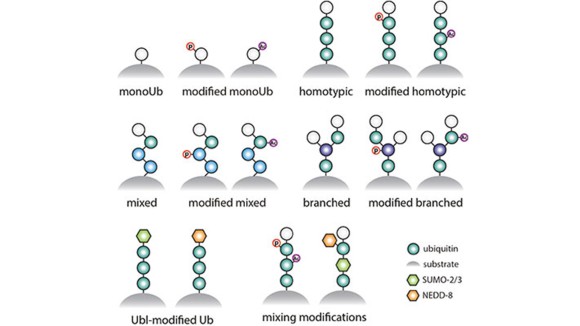
Ubiquitin and Ubl signaling
The April 2016 special issue on ubiquitin and Ubl signalling brings together a series of authoritative reviews from outstanding experts in the field. The current understanding, as well as recent advances in various aspects of ubiquitin and Ubl signalling, challenges in the fast growing field, and potentially important directions for future study, are presented. Further background information on this important topic is available through the accompanying web focus which links to related articles from across the Nature Publishing Group.