
Abstract
Background:
We have shown that a sodium ionophore monensin inhibits prostate cancer cell growth. A structurally related compound to monensin, salinomycin, was recently identified as a putative cancer stem cell inhibitor.
Methods:
The growth inhibitory potential of salinomycin was studied in a panel of prostate cells. To get insights into the mechanism of action, a variety of assays such as gene expression and steroid profiling were performed in salinomycin-exposed prostate cancer cells.
Results:
Salinomycin inhibited the growth of prostate cancer cells, but did not affect non-malignant prostate epithelial cells. Salinomycin impacted on prostate cancer stem cell functions as evidenced by reduced aldehyde dehydrogenase activity and the fraction of CD44+ cells. Moreover, salinomycin reduced the expression of MYC, AR and ERG, induced oxidative stress as well as inhibited nuclear factor-κB activity and cell migration. Furthermore, profiling steroid metabolites revealed increased levels of oxidative stress-inducing steroids 7-ketocholesterol and aldosterone and decreased levels of antioxidative steroids progesterone and pregnenolone in salinomycin-exposed prostate cancer cells.
Conclusion:
Our results indicate that salinomycin inhibits prostate cancer cell growth and migration by reducing the expression of key prostate cancer oncogenes, inducing oxidative stress, decreasing the antioxidative capacity and cancer stem cell fraction.
Similar content being viewed by others
Main
Redox regulation has an important role in controlling cancer cell behaviour. Cancer cells may potentially benefit from oxidative stress induction and production of reactive oxygen species (ROS), which are known to increase the rate of mutations (Auten and Davis, 2009; Acharya et al, 2010; Cairns et al, 2011). However, as cancer cells have a higher level of oxidative stress than non-malignant cells, cancer cells are dependent on an active antioxidant defence system. Thus, cancer cells are vulnerable to acute induction of oxidative stress caused by agents inducing ROS or reducing antioxidative capacity (Khandrika et al, 2009; Trachootham et al, 2009; Acharya et al, 2010; Cairns et al, 2011).
Interestingly, most key genes involved in prostate cancer modulate the antioxidative capacity. For example, the decrease in androgen receptor (AR) signalling has been shown to reduce the antioxidative capacity and increase ROS production (Tam et al, 2003; Pinthus et al, 2007), MYC expression is known to protect cells against oxidative stress (Benassi et al, 2006) and our recent results indicate that transmembrane protease, serine 2 (TMPRSS2)–the v-ets erythroblastosis virus E26 oncogene homolog (ERG) fusion-positive VCaP prostate cancer cells are vulnerable to oxidative stress induction (Iljin et al, 2009; Ketola et al, 2010; Vainio et al, 2011). Moreover, recent studies have shown that the enzymes that are used as cancer stem cell and tumour-initiating cell markers have antioxidative properties. Aldehyde dehydrogenase (ALDH) activity, used as a marker for prostate cancer stem cells (Moreb, 2008; Burger et al, 2009; Li et al, 2010; Yu et al, 2011), has an important role in maintaining antioxidative capacity and CD44, known to characterise cancer stem cells and to correlate with the ability of prostate cancer cells to migrate, regulates redox status in cancer cells (Klarmann et al, 2009; Ishimoto et al, 2011). Thus, these studies indicate that modulation of the redox status could be a potential therapeutic avenue to impact on prostate cancer cells, including the cancer-initiating cells.
We have recently identified monensin as a novel oxidative stress inducer and antineoplastic compound in prostate cancer cells (Iljin et al, 2009; Ketola et al, 2010). Interestingly, salinomycin, a structurally similar ionophorous antibiotic as monensin, was recently identified as a breast cancer stem cell inhibitor in vitro and shown to inhibit breast cancer xenograft growth in vivo (Gupta et al, 2009). Salinomycin reduces cancer and cancer stem cell growth also in other tumour types, such as leukaemias and uterine sarcoma cells (Fuchs et al, 2009, 2010). Traditionally, salinomycin has been used as an antimicrobial agent in veterinary medicine (Miyazaki et al, 1974; Mahmoudi et al, 2006) and it has been shown to inhibit oxidative phosphorylation in mitochondria by increasing the cation transport across the mitochondrial membrane (Mitani et al, 1976). Results from recent mechanistic studies in breast cancer cells indicate that salinomycin can induce DNA damage (Kim et al, 2010). However, the effect of salinomycin exposure on prostate cancer growth has not been studied before. Therefore, in this study, we explored the growth inhibitory potential of salinomycin in cultured prostate cancer and non-malignant prostate epithelial cells and studied its mechanism of action focusing on the relationship between antioxidant and cancer-initiating cell properties.
Materials and Methods
Cells
VCaP prostate carcinoma cells (TMPRSS2–ERG positive, received from Drs Adrie van Bokhoven, University of Colorado Health Sciences Center, Denver, CO, USA and Kenneth Pienta, University of Michigan, Ann Arbor, MI, USA) were grown in Dulbecco's modified Eagle's medium (Korenchuk et al, 2001). LNCaP prostate carcinoma cells (received from Dr Marco Cecchini, University of Bern, Bern, Switzerland) were grown in T-Medium (Invitrogen Molecular Probes, Carlsbad, CA, USA). The non-malignant RWPE-1 prostate epithelial cells (Webber et al, 1997) and prostate carcinoma cells PC-3 and DU 145 were purchased from American Type Culture Collection (LGC Promochem AB, Borås, Sweden) and grown according to provider's instructions. The non-malignant EP156T prostate epithelial cells were received from Dr Varda Rotter (Weizmann Institute of Science, Rehovot, Israel) and grown in the media recommended by the distributor (Kogan et al, 2006). Primary prostate epithelial cells were ordered from Lonza Walkersville, Inc. (Walkersville, MD, USA).
Compounds
Salinomycin and vitamin C were purchased from Sigma-Aldrich (St Louis, MO, USA) and diluted in DMSO.
Cell viability and apoptosis assays
Cell viability and apoptosis assays were done on Falcon 384-well plates (BD Biosciences, San Jose, CA, USA) as previously described (Ketola et al, 2010). Cell viability was determined with CellTiter-Glo cell viability assay (Promega, Madison, WI, USA) and induction of caspase 3 and 7 activities was detected with homogenous Apo-ONE assay (Promega) according to the manufacturer's instructions. The signals were quantified using Envision Multilabel Plate Reader (Perkin-Elmer, Massachusetts, MA, USA).
Determination of ALDH activity
The activity of ALDH in response to 1 μ M salinomycin exposure in prostate cancer cells was determined with Aldefluor reagent (Stemcell Technologies, Vancouver, BC, Canada) as previously described (Ketola et al, 2010). ALDH inhibitor diethylaminobenzaldehyde (DEAB) was used as a negative control. The fluorometric signal was determined with Envision Multilabel Reader (Perkin-Elmer).
Fluorescence-activated cell-sorting analysis
VCaP, LNCaP, PC-3 and DU-145 cells were exposed to salinomycin (1 μ M) for 6 h, samples were fixed with 2% paraformaldehyde and stained with CD44 (FITC-conjugated mouse monoclonal anti-human, BD Pharmingen (San Diego, CA, USA) 555478, BD Pharmingen) antibody for 45 min at 4°C in the dark. Cells were washed and the fluorescence intensity was measured using Accuri C6 Flow Cytometer (BD Accuri Cytometers, Ann Arbor, MI, USA).
Gene expression analysis using bead arrays
VCaP cells were treated with 1 μ M salinomycin for 3, 6 and 24 h, total RNA was extracted and RNA integrity was monitored using an Experion electrophoresis station (Bio-Rad Laboratories, Hercules, CA, USA). Purified RNA (300 ng) was used for amplification with the Illumina RNA TotalPrep Amplification kit (Ambion, Austin, TX, USA) and the biotin-labeled cRNA was hybridised to Sentrix HumanRef-8 vs 3 Expression BeadChips (Illumina, San Diego, CA, USA). The arrays were scanned with the BeadArray Reader (Illumina).
Statistical analysis of gene expression data
The raw gene expression data were quantile-normalised (Gentleman et al, 2004) and analysed as previously described (Ketola et al, 2010). The gene names from Illumina experiments were rendered to Affymetrix gene IDs with Ensemble Genes 59 database. Ingenuity Pathway Analysis (IPA) software (Ingenuity Systems Inc., Redwood City, CA, USA) was used to analyse the functional gene ontology and pathway annotations. Differentially expressed genes (logFC >0.5 or <−0.5) were selected for the IPA analysis. Connectivity Map 02 was used to identify drugs with similar or opposite effects on gene expression (Lamb, 2007).
Statistical analyses
The error bars in the figures are shown as standard deviations. The asterisks indicate statistical significance. *P<0.05; **P<0.01; ***P<0.001.
RNA extraction and quantitative reverse transcriptase PCR
Total RNA was extracted and quantitative real-time PCR was done as previously described (Ketola et al, 2010). TaqMan gene expression probes and primers from the Universal Probe Library (Roche Diagnostics, Espoo, Finland) were used to study AR, prostate-specific antigen (PSA), ERG, MYC, Kruppel-like factor 6 (KLF6) and activating transcription factor 3 (ATF3), metallothioneins MT1G and MT1F, thioredoxin-binding protein (TXNIP), DNA damage-inducible transcripts 3 and 4 (DDIT3 and DDIT4) and β-actin mRNA expression (Supplementary Table 1). Three replicate samples were studied for quantitation of mRNA expression.
Western blot analysis
Western blot analysis was performed for compound-treated samples using specific antibodies against AR (1 : 1000 dilution, mouse monoclonal, Labvision, Fremont, CA, USA), PSA (1 : 1000 dilution, rabbit polyclonal, DakoCytomation, Glostrup, Denmark) and β-actin (1 : 4000 dilution, mouse-monoclonal, Becton Dickinson, Franklin Lakes, NJ, USA). Signal was detected with 1 : 4000 dilution of appropriate HRP-conjugated secondary antibodies (all from Invitrogen Molecular Probes, Carlsbad, CA, USA) followed by visualisation with the enhanced chemiluminescence reagent (Amersham Biosciences, Little Chalfont, UK).
Reactive oxygen species detection
The intracellular ROS was measured in response to salinomycin exposure for 48 h with carboxy-H2DCFDA as previously described (Ketola et al, 2010). As a positive ROS control, hydrogen peroxide exposure (400 μ M) for 4 h was used. The oxidation of the probe was measured in PBS by monitoring the increase in fluorescence with Envision Multilabel Plate Reader (Perkin Elmer).
Cancer luciferase reporter assay
The activity of nuclear factor-κB (NF-κB) signalling pathway was measured using luciferase reporter assay (SABiosciences, Frederick, MD, USA). In brief, inducible NF-κB transcription factor responsive firefly luciferase reporter with constitutively expressing Renilla construct transcription factor reporter were transfected in prostate cancer cells. A mixture of non-inducible firefly luciferase reporter and constitutively expressing Renilla construct was used as a negative control. After 24 h, salinomycin (100 nM) or control were added onto the cells for 18 h. The Dual-LuciferaseReporter (DLR) Assay System (Promega) was used to measure the luciferase activities and results were analysed according to the manufacturer's instructions.
Wound-healing assay
The effect of salinomycin (100 nM and 1 μ M) alone and in combination with vitamin C (10 μ M) on prostate cancer cell migration was studied using a wound-healing assay. PC-3 cells were plated on 96-well plates (Essen ImageLock, Essen Instruments, Birmingham, UK) and a wound was scratched with wound scratcher (Essen Instruments). Compounds and appropriate controls were added immediately after wound scratching and wound confluence was monitored with Incucyte Live-Cell Imaging System and software (Essen Instruments). Wound closure was observed every hour for 24 h by comparing the mean relative wound density of three biological replicates in each experiment.
Steroid quantification
VCaP cells were exposed to 1 μ M salinomycin for 6 h, harvested and counted. An internal standard (labelled C16 : 0) and chloroform/methanol (2 : 5) mixture were added, the samples were homogenised with Retsch system (5 min, 20 Hz), centrifuged and the supernatant was collected and evaporated. MOX (25 μl, TS-45950, Thermo Scientific, Helsinki, Finland) was added and the mixture was incubated at 45°C for 60 min. Next, 100 μl of MSTFA with 1% trimethylchlorosilane (Fluka, St Louis, MO, USA) was added and the mixture was incubated at 70°C for 60 min. Injection standard was added to the mixture before gas chromatography-mass spectrometry analysis (GC-MS, Agilent 6890 gas chromatograph (GC) combined with Agilent 5973 mass selective detector (MSD), Agilent Technologies, Espoo, Finland). The injector (injection volume 1 μl with pulsed splitless injection) and MSD temperatures were 230°C (MS Source) and 150°C (MS Quad). The analyses were performed on Supelco 38499-02C capillary column. Selective ion monitoring using specific masses for each target analyte was used in the detection. The following steroids were quantified: 7-ketocholesterol, aldosterone, progesterone, pregnenolone, estrone, 17B-estradiol, 4B-hydroxycholesterol, 25-hydroxycholesterol, 5a,6a-epoxycholesterol (Mono-TMS), dihydrotestosterone and testosterone (the standards were from Steraloids, Newport, RI, USA).
Results
Salinomycin inhibits prostate cancer cell growth but does not induce apoptosis
The effect of salinomycin on cell growth was studied in panel of malignant (VCaP, LNCaP, PC-3, DU 145) and non-malignant (RWPE-1, EP156T and PrEC) prostate cells. Interestingly, salinomycin was the most effective in inhibiting VCaP cells (EC50=380 nM), whereas non-malignant prostate epithelial cells RWPE-1, EP156T and PrEC were non-responsive (EC50>10 μ M) (Table 1). Salinomycin was also at least 10-fold more potent growth inhibitor in other prostate cancer cells studied compared with non-malignant prostate epithelial cells (Table 1). To determine whether salinomycin induces apoptosis in VCaP and LNCaP cells, caspase 3 and 7 activities were determined by a quantitative fluorometric assay. No significant increase in caspase activity was observed in response to salinomycin exposure (up to 10 μ M) for 48 h in VCaP and LNCaP cells (Supplementary Figure S1). Therefore, salinomycin reduces the growth of prostate cancer cells, but does not induce apoptosis.
Salinomycin inhibits ALDH activity and reduces CD44 cell fraction
Aldehyde dehydrogenase activity is considered as a marker for stem cell potential (Moreb, 2008) and tumourigenic prostate cancer cells (Burger et al, 2009; Li et al, 2010; Yu et al, 2011). As salinomycin has been previously identified as a cancer stem cell growth inhibitor (Gupta et al, 2009), we studied whether salinomycin reduces ALDH activity in VCaP and LNCaP prostate cancer cells. The results indicated that salinomycin exposure for 48 h resulted in a significant decrease in ALDH activity in VCaP (by 30%) and LNCaP (by 26%) cells (Figure 1). To confirm the impact on prostate cancer stem cells, the intensity of cancer stem cell marker CD44 in VCaP, LNCaP, PC-3 and DU-145 prostate cancer cells was studied in response salinomycin exposure (1 μ M) using fluorescence-activated cell-sorting analysis. The results indicated that salinomycin reduced the amount of CD44+ cells in all prostate cancer cells tested already after 6-h exposure (Figure 2). The division of CD44-positive and -negative cells (shown as scattered lines in Figure 2) in all cell lines are presented in Supplementary Figure S2. Taken together, these results indicate that salinomycin decreases ALDH activity and CD44+ cancer stem cell fraction in cultured prostate cancer cells.

Salinomycin inhibits aldehyde dehydrogenase (ALDH) activity in prostate cancer cells. ALDH activity was measured with Aldefluor assay in response to 1-mmol l−1 exposures of salinomycin or control for 48 h in VCaP and LNCaP cells. ALDH inhibitor diethylaminobenzaldehyde (DEAB) was used as negative control. Asterisks indicate statistical significance. *P<0.05; **P<0.01; ***P<0.001.

Salinomycin reduces the fraction of CD44+ cells in prostate cancer cells. VCaP, LNCaP, PC-3 and DU-145 cells were stained with CD44 antibody and the fluorescence intensities were identified for cells exposed to 6 h of salinomycin or DMSO control.
Salinomycin reduces AR signalling and alters genes involved in lipid metabolism, cell-cycle checkpoint regulation and oxidative stress response
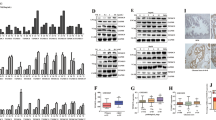
To get additional insights into the salinomycin-induced alterations in prostate cancer cells, quantitative RT-PCR and genome-wide gene expression profiles were analysed in prostate cancer cells. First, the effect of salinomycin on the expression of key genes involved in prostate cancer, AR, ERG and MYC was studied. The results indicated that salinomycin decreased AR mRNA levels in VCaP and in LNCaP cells (by 32% and 56% at 6-h time points; Figure 3A). Accordingly, a decrease in the levels of PSA mRNA, considered as a marker of the activity of androgen signalling, was also seen in response to salinomycin exposure for 24 h in VCaP and in LNCaP cells (by 50 and 90%; Figure 3B). The decrease in AR and PSA protein levels was also confirmed (Supplementary Figure S3). In addition, salinomycin reduced ERG mRNA expression in VCaP cells (by 30%; Figure 3C) and MYC mRNA levels in both VCaP and LNCaP cells (by 60%; Figure 3D).

Salinomycin reduces the expression levels of key prostate cancer oncogenes, androgen receptor (AR), ERG and MYC in prostate cancer cells. (A) AR, (B) PSA, (C) ERG and (D) MYC mRNA expression in response to 3-, 6-, and 24-h salinomycin exposure in VCaP and LNCaP cells. Asterisks indicate statistical significance. *P<0.05; **P<0.01; ***P<0.001.
Second, salinomycin-induced alterations in gene expression profiling were analysed in VCaP cells using IPA. The most prominent changes at all time points studied were seen in lipid metabolism and in cholesterol and steroid biosynthesis (Supplementary Tables S2 and S3). In addition, cell-cycle checkpoint regulation and aryl hydrocarbon receptor (AhR) signalling were deregulated at all time points, with the most significant changes seen after a 24-h exposure. However, the level of AhR mRNA were not changed (Supplementary Figure S4). Moreover, changes were seen in oxidative stress response, mitochondrial membrane characteristics and cellular movement (Supplementary Tables S2 and S3). Taken together, these results indicate that salinomycin reduces the expression of key prostate cancer oncogenes and AR signalling as well as causes alterations in steroid biosynthesis, cell-cycle checkpoint regulation, AhR signalling and oxidative stress response in prostate cancer cells.
Salinomycin induces oxidative stress
The alterations in oxidative stress response were seen in the gene expression analysis. Thus, the expression of oxidative stress markers TXNIP, MT1G, MT1F, ATF3, KLF6, DDIT3 and DDIT4 were further validated using quantitative real-time PCR. The results indicated that salinomycin induced the gene expression signature characteristic of oxidative stress induction in both VCaP and LNCaP cells (Figure 4A, Supplementary Figure S5 and S6). However, the induction was not as strong in LNCaP cells as in VCaP cells. To validate the oxidative stress induction, the increase in intracellular level of ROS was studied using carboxy-H2DCFDA marker. The results confirmed that salinomycin increases ROS in both VCaP and LNCaP cells (Figure 4B).

(A) Salinomycin induces the gene expression signature characteristic of oxidative stress induction in prostate cancer cells. The expressions of MT1F, MT1G, ATF3, KLF6, DDIT3, DDIT4 and TXNIP mRNAs in VCaP and LNCaP cells in response to salinomycin exposure for 3, 6 and 24 h. (B) Salinomycin induces the level of intracellular reactive oxygen species in prostate cancer cells. Reactive oxygen species generation in response to salinomycin exposure in VCaP and LNCaP cells detected with carboxy-H2DCFDA. Hydrogen peroxide (400 mmol l−1) exposure for 4 h was used as a positive control. Asterisks indicate statistical significance. *P<0.05; **P<0.01; ***P<0.001.
Salinomycin shows similar effects as niclosamide and terfenadine
Analysis of connectivity map data was used to identify compounds with similar or opposite effects as salinomycin. The differentially expressed genes in response to salinomycin exposure for 6 h in VCaP cells were compared with the >7000 expression profiles representing drug responses to >1309 compounds. Niclosamide and terfenadine were the most enriched compounds altering gene expression in the same direction as salinomycin (Supplementary Table S4). Interestingly, niclosamide is an antihelmintic drug that was recently shown to inactivate the NF-κB pathway and to generate ROS in leukaemic stem cells, whereas terfenadine is an antihistamine that induces apoptosis in melanoma cells and causes massive hydrogen peroxide production in cultured cerebellar neurons (Diaz-Trelles et al, 2000; Jangi et al, 2008; Jin et al, 2010). These results give further support to the fact that salinomycin induces oxidative stress in prostate cancer cells.
Salinomycin reduces NF-κB pathway activity in prostate cancer cells
Studies of the NF-κB inactivator niclosamide suggest that inhibition of NF-κB activity could also have a role in the induction of oxidative stress in salinomycin-exposed prostate cancer cells. Thus, we studied the effects of salinomycin on NF-κB pathway activity using a cancer reporter assay. Interestingly, NF-κB pathway was 10 times higher in VCaP than in LNCaP cells (Figure 5). The results indicated that salinomycin reduces NF-κB activity in VCaP (by 52%) and LNCaP (by 48%) cells (Figure 5). These results support the hypothesis that salinomycin-induced growth inhibition and induction of oxidative stress in prostate cancer cells is mediated by the inhibition of NF-κB activity.

Salinomycin reduces NF-κB activity in prostate cancer cells. The activity of NF-κB and negative control was measured using cancer reporter array in (A) VCaP and in (B) LNCaP cells in response to 100 nM salinomycin exposure for 24 h and the results were compared with control exposures. Asterisks indicate statistical significance. *P<0.05; **P<0.01.
Salinomycin reduces prostate cancer cell migration
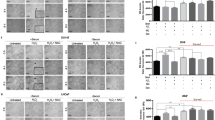
The analysis of gene expression profiling results indicated that cellular movement and migration were altered in response to 6-h exposure of salinomycin (Supplementary Table S2). Furthermore, NF-κB and ALDH activities as well as CD44+ cells have all been highlighted as markers of prostate cancer cell migration (Klarmann et al, 2009; Zhang et al, 2009b; van den Hoogen et al, 2010). Thus, we explored the effect of salinomycin on prostate cancer cell motility using a wound-healing assay. As VCaP and LNCaP cells do not migrate and therefore are not suitable for this experiment, PC-3 prostate cancer cells were used. Cells were exposed to salinomycin (100 nM) and wound confluence was monitored every hour for 24 h. The results indicated that salinomycin reduces the migration of PC-3 cells. As the EC50 value of salinomycin in PC-3 cells in response to 48-h exposure was higher than 1 μ M, the salinomycin-induced anti-migratory effect is not due to inhibition of cell proliferation. This was confirmed also by cell viability assay (data not shown). To study whether salinomycin-induced oxidative stress has a role in the migratory phenotype, PC-3 cells were exposed to antioxidant vitamin C (10 μ M) and salinomycin (100 nM) and the wound confluence was monitored for 24 h. The results show that vitamin C antagonises the anti-migratory effect of salinomycin in PC-3 cells, indicating that induction of oxidative stress inhibits prostate cancer cell migration (Figure 6).

Salinomycin reduces PC-3 prostate cancer cell migration and the effect is antagonised with antioxidant vitamin C. (A) Relative wound density in response to salinomycin alone (100 nM, P<0.001 for control vs salinomycin) and in combination with vitamin C (10 μM) as well as appropriate controls monitored for 24 h. (B) The morphological pictures from wound density after 24-h exposures of salinomycin alone and in combination vitamin C. The cells in the beginning of the experiment are marked in black.
Salinomycin induces 7-ketocholesterol and aldosterone levels as well as reduces progesterone and pregnenolone levels
As salinomycin altered lipid metabolism and steroid biosynthesis as well as reduced AR signalling, steroid profiling was performed in salinomycin-exposed VCaP cells using GC-MS. The results showed that the most prominent changes in response to salinomycin exposure were the increase in 7-ketocholesterol and aldosterone levels as well as the decrease in progesterone and pregnenolone (Figure 7, Supplementary Table S5). Interestingly, 7-ketocholesterol is a cholesterol oxidising product that induces ROS, apoptosis, mitochondrial DNA damage and dysfunction as well as ER stress (Leonarduzzi et al, 2006; Lee et al, 2009; Gramajo et al, 2010). Recently, aldosterone has also been linked to oxidative stress induction (Calo et al, 2010; Queisser et al, 2011). In contrast, progesterone and pregnenolone are precursors of androgens and progesterone has shown to have antioxidant properties (Ozacmak and Sayan, 2009). Moreover, progesterone has recently been identified as an inducer of adult mammary stem cell expansion (Joshi et al, 2010). Taken together, salinomycin induces steroids involved in oxidative stress induction and reduces steroids that sustain antioxidative capacity and induce stem cell expansion.

Salinomycin induces the levels of 7-ketocholesterol and aldosterone and reduces the levels of progesterone and pregnenolone. The cells were exposed to salinomycin (1 μ M) for 6 h and the steroid profile was measured with gas chromatography-mass spectrometry.
Discussion
The treatment options for advanced prostate cancer are limited. We have recently identified monensin, a widely used antibiotic in poultry, as an inducer of oxidative stress and a potent inhibitor of prostate cancer cell growth. Antibiotic compound salinomycin shares a similar structure as monensin and was recently identified as a novel cancer stem cell inhibitor in breast cancer (Miyazaki et al, 1974; Danforth et al, 1977; Mahmoudi et al, 2006; Gupta et al, 2009). The inhibition of cancer stem cells has been suggested as a potential new therapeutic option for advanced and metastatic cancer (Clayton and Mousa, 2011). In addition, cancer stem cells are known to have strong antioxidative defence mechanisms and the reduction of antioxidative genes has been suggested as a means to target cancer stem cells (Kobayashi and Suda, 2012). Interestingly, many key oncogenes in prostate cancer cells are also known to induce antioxidative properties. However, cancer stem cell inhibitors have not been identified for prostate cancer therapy. Thus, here we studied the growth inhibitory potential and the mechanism of action of salinomycin in cultured human prostate cancer cells.
In this study, we showed for the first time that prostate cancer cells, but not non-tumourigenic prostate epithelial cells, are sensitive to salinomycin. Although salinomycin reduced prostate cancer cell growth, it did not markedly induce caspase 3- and 7-mediated apoptosis. We also showed that salinomycin had an impact on prostate cancer stem cell population using ALDH activity and the amount of CD44+ cells as markers. Aldehyde dehydrogenase activity has been used as a marker of prostate cancer stem cells and proposed as a marker of poor outcome in prostate cancer (Davydov et al, 2004; Burger et al, 2009; Zhang et al, 2009a; Li et al, 2010; Yu et al, 2011). A decrease in cancer stem cell population in response to salinomycin exposure has been shown with breast leukaemia and lung cancer (Gupta et al, 2009; Fuchs et al, 2009, 2010; Wang, 2011). Aldehyde dehydrogenase enzymes catalyse the dehydrogenation of aldehydes, and thereby protect cells from oxidative stress (Davydov et al, 2004; Klarmann et al, 2009; Zhang et al, 2009a). In addition, CD44 is known to defend cancer cells against oxidative stress by increasing reduced glutathione synthesis (Ishimoto et al, 2011). We also identified that salinomycin reduced the expression of prostate cancer oncogenes, MYC, AR and ERG, which are known to have antioxidative properties (Tam et al, 2003; Benassi et al, 2006; Pinthus et al, 2007; Swanson et al, 2011). Thus, the results suggest that salinomycin may inhibit prostate cancer stem cells by impairing the redox control. The altered redox regulation in response to salinomycin exposure was confirmed by the induction of intracellular ROS production as well as the gene expression signature characteristic of oxidative stress. Taken together, the antineoplastic effects of salinomycin resulted from decreased cancer stem cell population, reduced expression of key oncogenes and induction of oxidative stress in cultured prostate cancer cells.
Connectivity map analysis indicated that salinomycin has similar effects as terfenadine, known to induce ROS, and niclosamide, an inhibitor of NF-κB. The decrease in NF-κB activity in salinomycin-exposed cells was validated in prostate cancer cells. Nuclear factor-κB is known to regulate cellular antioxidant defence capacity as well as prostate cancer cell viability, tumourigenesis and metastasis (Gloire et al, 2006; Sarkar et al, 2008; Gluschnaider et al, 2010). Nuclear factor-κB pathway is active in prostate stem-like tumour-initiating cells and its inhibition induces apoptosis in prostate cancer stem cells. Therefore, NF-κB is considered as a promising therapeutic target (Birnie et al, 2008; Jin et al, 2008; Rajasekhar et al, 2011). Nuclear factor-κB was recently shown to be activated by specific TMPRSS2–ERG fusion isoforms, which may explain the 10-fold increase in NF-κB activity seen in VCaP cells in comparison to the LNCaP cells (Wang et al, 2010). As NF-κB and ALDH activities as well as CD44+ cell population have all been previously shown to regulate prostate cancer cell migration (Klarmann et al, 2009; Zhang et al, 2009b; van den Hoogen et al, 2010), the migratory effect of salinomycin was studied in prostate cancer cells. Our results revealed that salinomycin exposure inhibited prostate cancer cell migration and was antagonised by antioxidant vitamin C, indicating that induction of oxidative stress has an important role in mediating the salinomycin-induced anti-migratorial phenotype. Salinomycin was recently also shown to reduce cell invasion in cultured colorectal cancer cells (Dong et al, 2011).
Results from steroid profiling showed that salinomycin reduced the levels of progesterone and pregnelonone, both precursors of androgens. Progesterone is known to induce antioxidative capacity and mammary stem cell expansion (Ozacmak and Sayan, 2009; Joshi et al, 2010). Moreover, we show that salinomycin induced the levels of 7-ketocholesterol and aldosterone, which are known to act as oxidative stress inducers (Leonarduzzi et al, 2006; Lee et al, 2009; Calo et al, 2010; Gramajo et al, 2010; Queisser et al, 2011). The cholesterol oxidation product 7-ketocholesterol is a ligand for AhR and may function as an AhR antagonist (Savouret et al, 2001). Aryl hydrocarbon receptor pathway has been suggested to have an essential role in detoxification of foreign chemicals and in the protection against oxidative stress by increasing the expression of ALDH proteins (Lindros et al, 1998; Nebert et al, 2000; Vrzal et al, 2004; Kohle and Bock, 2007). Moreover, AhR is overexpressed in prostate cancer and cancer stem cells and it can bind to NF-κB and promote activation of MYC (Kim et al, 2000; Blum et al, 2009; Gluschnaider et al, 2010). Our gene expression analysis results showed that AhR target genes were reduced by salinomycin, although the AhR receptor mRNA levels were not changed. However, many processes known to be modulated via AhR were altered in salinomycin-exposed prostate cancer cells.
In conclusion, our results reveal that the ability of salinomycin to inhibit prostate cancer cell growth and cancer stem cell population, without major effects on non-malignant prostate epithelial cells, is due to the induction of oxidative stress and the reduction of antioxidative properties. Thus, salinomycin and its derivatives may provide a novel selective approach for prostate cancer therapy.
Change history
29 March 2012
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
Acharya A, Das I, Chandhok D, Saha T (2010) Redox regulation in cancer: a double-edged sword with therapeutic potential. Oxid Med Cell Longev 3: 23–34
Auten RL, Davis JM (2009) Oxygen toxicity and reactive oxygen species: the devil is in the details. Pediatr Res 66: 121–127
Benassi B, Fanciulli M, Fiorentino F, Porrello A, Chiorino G, Loda M, Zupi G, Biroccio A (2006) c-Myc phosphorylation is required for cellular response to oxidative stress. Mol Cell 21: 509–519
Birnie R, Bryce SD, Roome C, Dussupt V, Droop A, Lang SH, Berry PA, Hyde CF, Lewis JL, Stower MJ, Maitland NJ, Collins AT (2008) Gene expression profiling of human prostate cancer stem cells reveals a pro-inflammatory phenotype and the importance of extracellular matrix interactions. Genome Biol 9: R83
Blum R, Gupta R, Burger PE, Ontiveros CS, Salm SN, Xiong X, Kamb A, Wesche H, Marshall L, Cutler G, Wang X, Zavadil J, Moscatelli D, Wilson EL (2009) Molecular signatures of prostate stem cells reveal novel signaling pathways and provide insights into prostate cancer. PLoS One 4: e5722
Burger PE, Gupta R, Xiong X, Ontiveros CS, Salm SN, Moscatelli D, Wilson EL (2009) High aldehyde dehydrogenase activity: a novel functional marker of murine prostate stem/progenitor cells. Stem Cells 27: 2220–2228
Cairns RA, Harris IS, Mak TW (2011) Regulation of cancer cell metabolism. Nat Rev Cancer 11: 85–95
Calo LA, Pagnin E, Davis PA, Armanini D, Mormino P, Rossi GP, Pessina AC (2010) Oxidative stress-related proteins in a Conn's adenoma tissue. Relevance for aldosterone′s prooxidative and proinflammatory activity. J Endocrinol Invest 33: 48–53
Clayton S, Mousa SA (2011) Therapeutics formulated to target cancer stem cells: Is it in our future? Cancer Cell Int 11: 7
Danforth HD, Ruff MD, Reid WM, Miller RL (1977) Anticoccidial activity of salinomycin in battery raised broiler chickens. Poult Sci 56: 926–932
Davydov VV, Dobaeva NM, Bozhkov AI (2004) Possible role of alteration of aldehyde's scavenger enzymes during aging. Exp Gerontol 39: 11–16
Diaz-Trelles R, Novelli A, Vega JA, Marini A, Fernandez-Sanchez MT (2000) Antihistamine terfenadine potentiates NMDA receptor-mediated calcium influx, oxygen radical formation, and neuronal death. Brain Res 880: 17–27
Dong TT, Zhou HM, Wang LL, Feng B, Lv B, Zheng MH (2011) Salinomycin selectively targets ‘CD133+’ cell subpopulations and decreases malignant traits in colorectal cancer lines. Ann Surg Oncol 18: 1797–1804
Fuchs D, Daniel V, Sadeghi M, Opelz G, Naujokat C (2010) Salinomycin overcomes ABC transporter-mediated multidrug and apoptosis resistance in human leukemia stem cell-like KG-1a cells. Biochem Biophys Res Commun 394: 1098–1104
Fuchs D, Heinold A, Opelz G, Daniel V, Naujokat C (2009) Salinomycin induces apoptosis and overcomes apoptosis resistance in human cancer cells. Biochem Biophys Res Commun 390: 743–749
Gentleman RC, Carey VJ, Bates DM, Bolstad B, Dettling M, Dudoit S, Ellis B, Gautier L, Ge Y, Gentry J, Hornik K, Hothorn T, Huber W, Iacus S, Irizarry R, Leisch F, Li C, Maechler M, Rossini AJ, Sawitzki G, Smith C, Smyth G, Tierney L, Yang JY, Zhang J (2004) Bioconductor: open software development for computational biology and bioinformatics. Genome Biol 5: R80
Gloire G, Legrand-Poels S, Piette J (2006) NF-kappaB activation by reactive oxygen species: fifteen years later. Biochem Pharmacol 72: 1493–1505
Gluschnaider U, Hidas G, Cojocaru G, Yutkin V, Ben-Neriah Y, Pikarsky E (2010) beta-TrCP inhibition reduces prostate cancer cell growth via upregulation of the aryl hydrocarbon receptor. PLoS One 5: e9060
Gramajo AL, Zacharias LC, Neekhra A, Luthra S, Atilano SR, Chwa M, Brown DJ, Kuppermann BD, Kenney MC (2010) Mitochondrial DNA damage induced by 7-ketocholesterol in human retinal pigment epithelial cells in vitro. Invest Ophthalmol Vis Sci 51: 1164–1170
Gupta PB, Onder TT, Jiang G, Tao K, Kuperwasser C, Weinberg RA, Lander ES (2009) Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 138: 645–659
Iljin K, Ketola K, Vainio P, Halonen P, Kohonen P, Fey V, Grafstrom RC, Perala M, Kallioniemi O (2009) High-throughput cell-based screening of 4910 known drugs and drug-like small molecules identifies disulfiram as an inhibitor of prostate cancer cell growth. Clin Cancer Res 15: 6070–6078
Ishimoto T, Nagano O, Yae T, Tamada M, Motohara T, Oshima H, Oshima M, Ikeda T, Asaba R, Yagi H, Masuko T, Shimizu T, Ishikawa T, Kai K, Takahashi E, Imamura Y, Baba Y, Ohmura M, Suematsu M, Baba H, Saya H (2011) CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc(-) and thereby promotes tumor growth. Cancer Cell 19: 387–400
Jangi SM, Ruiz-Larrea MB, Nicolau-Galmes F, Andollo N, Arroyo-Berdugo Y, Ortega-Martinez I, Diaz-Perez JL, Boyano MD (2008) Terfenadine-induced apoptosis in human melanoma cells is mediated through Ca2+ homeostasis modulation and tyrosine kinase activity, independently of H1 histamine receptors. Carcinogenesis 29: 500–509
Jin RJ, Lho Y, Connelly L, Wang Y, Yu X, Saint Jean L, Case TC, Ellwood-Yen K, Sawyers CL, Bhowmick NA, Blackwell TS, Yull FE, Matusik RJ (2008) The nuclear factor-kappaB pathway controls the progression of prostate cancer to androgen-independent growth. Cancer Res 68: 6762–6769
Jin Y, Lu Z, Ding K, Li J, Du X, Chen C, Sun X, Wu Y, Zhou J, Pan J (2010) Antineoplastic mechanisms of niclosamide in acute myelogenous leukemia stem cells: inactivation of the NF-kappaB pathway and generation of reactive oxygen species. Cancer Res 70: 2516–2527
Joshi PA, Jackson HW, Beristain AG, Di Grappa MA, Mote PA, Clarke CL, Stingl J, Waterhouse PD, Khokha R (2010) Progesterone induces adult mammary stem cell expansion. Nature 465: 803–807
Ketola K, Vainio P, Fey V, Kallioniemi O, Iljin K (2010) Monensin is a potent inducer of oxidative stress and inhibitor of androgen signaling leading to apoptosis in prostate cancer cells. Mol Cancer Ther 9: 3175–3185
Khandrika L, Kumar B, Koul S, Maroni P, Koul HK (2009) Oxidative stress in prostate cancer. Cancer Lett 282: 125–136
Kim DW, Gazourian L, Quadri SA, Romieu-Mourez R, Sherr DH, Sonenshein GE (2000) The RelA NF-kappaB subunit and the aryl hydrocarbon receptor (AhR) cooperate to transactivate the c-myc promoter in mammary cells. Oncogene 19: 5498–5506
Kim JH, Chae M, Kim WK, Kim YJ, Kang HS, Kim HS, Yoon S (2010) Salinomycin sensitizes cancer cells to the effects of doxorubicin and etoposide treatment by increasing DNA damage and reducing p21 protein. Br J Pharmacol 162: 773–784
Klarmann GJ, Hurt EM, Mathews LA, Zhang X, Duhagon MA, Mistree T, Thomas SB, Farrar WL (2009) Invasive prostate cancer cells are tumor initiating cells that have a stem cell-like genomic signature. Clin Exp Metastasis 26: 433–446
Kobayashi CI, Suda T (2012) Regulation of reactive oxygen species in stem cells and cancer stem cells. J Cell Physiol 227 (2): 421–430
Kogan I, Goldfinger N, Milyavsky M, Cohen M, Shats I, Dobler G, Klocker H, Wasylyk B, Voller M, Aalders T, Schalken JA, Oren M, Rotter V (2006) hTERT-immortalized prostate epithelial and stromal-derived cells: an authentic in vitro model for differentiation and carcinogenesis. Cancer Res 66: 3531–3540
Kohle C, Bock KW (2007) Coordinate regulation of phase I and II xenobiotic metabolisms by the Ah receptor and Nrf2. Biochem Pharmacol 73: 1853–1862
Korenchuk S, Lehr JE, MClean L, Lee YG, Whitney S, Vessella R, Lin DL, Pienta KJ (2001) VCaP, a cell-based model system of human prostate cancer. In Vivo 15: 163–168
Lamb J (2007) The Connectivity Map: a new tool for biomedical research. Nat Rev Cancer 7: 54–60
Lee WH, Lee CS, Kwon K, Kwon YS, Kim SW, Goo TW, Kwon OY (2009) 7-Ketocholesterol induces endoplasmic reticulum stress in HT-29 cells. Z Naturforsch C 64: 307–310
Leonarduzzi G, Vizio B, Sottero B, Verde V, Gamba P, Mascia C, Chiarpotto E, Poli G, Biasi F (2006) Early involvement of ROS overproduction in apoptosis induced by 7-ketocholesterol. Antioxid Redox Signal 8: 375–380
Li T, Su Y, Mei Y, Leng Q, Leng B, Liu Z, Stass SA, Jiang F (2010) ALDH1A1 is a marker for malignant prostate stem cells and predictor of prostate cancer patients’ outcome. Lab Invest 90: 234–244
Lindros KO, Oinonen T, Kettunen E, Sippel H, Muro-Lupori C, Koivusalo M (1998) Aryl hydrocarbon receptor-associated genes in rat liver: regional coinduction of aldehyde dehydrogenase 3 and glutathione transferase Ya. Biochem Pharmacol 55: 413–421
Mahmoudi N, de Julian-Ortiz JV, Ciceron L, Galvez J, Mazier D, Danis M, Derouin F, Garcia-Domenech R (2006) Identification of new antimalarial drugs by linear discriminant analysis and topological virtual screening. J Antimicrob Chemother 57: 489–497
Mitani M, Yamanishi T, Miyazaki Y, Otake N (1976) Salinomycin effects on mitochondrial ion translocation and respiration. Antimicrob Agents Chemother 9: 655–660
Miyazaki Y, Shibuya M, Sugawara H, Kawaguchi O, Hirsoe C (1974) Salinomycin, a new polyether antibiotic. J Antibiot (Tokyo) 27: 814–821
Moreb JS (2008) Aldehyde dehydrogenase as a marker for stem cells. Curr Stem Cell Res Ther 3: 237–246
Nebert DW, Roe AL, Dieter MZ, Solis WA, Yang Y, Dalton TP (2000) Role of the aromatic hydrocarbon receptor and [Ah] gene battery in the oxidative stress response, cell cycle control, and apoptosis. Biochem Pharmacol 59: 65–85
Ozacmak VH, Sayan H (2009) The effects of 17beta estradiol, 17alpha estradiol and progesterone on oxidative stress biomarkers in ovariectomized female rat brain subjected to global cerebral ischemia. Physiol Res 58: 909–912
Pinthus JH, Bryskin I, Trachtenberg J, Lu JP, Singh G, Fridman E, Wilson BC (2007) Androgen induces adaptation to oxidative stress in prostate cancer: implications for treatment with radiation therapy. Neoplasia 9: 68–80
Queisser N, Oteiza PI, Stopper H, Oli RG, Schupp N (2011) Aldosterone induces oxidative stress, oxidative DNA damage and NF-kappaB-activation in kidney tubule cells. Mol Carcinog 50: 123–135
Rajasekhar VK, Studer L, Gerald W, Socci ND, Scher HI (2011) Tumour-initiating stem-like cells in human prostate cancer exhibit increased NF-kappaB signalling. Nat Commun 2: 162
Sarkar FH, Li Y, Wang Z, Kong D (2008) NF-kappaB signaling pathway and its therapeutic implications in human diseases. Int Rev Immunol 27: 293–319
Savouret JF, Antenos M, Quesne M, Xu J, Milgrom E, Casper RF (2001) 7-Ketocholesterol is an Endogenous Modulator for the Arylhydrocarbon Receptor. J Biol Chem 276: 3054–3059
Swanson TA, Krueger SA, Galoforo S, Thibodeau BJ, Martinez AA, Wilson GD, Marples B (2011) TMPRSS2/ERG fusion gene expression alters chemo- and radio-responsiveness in cell culture models of androgen independent prostate cancer. Prostate 71 (14): 1548–1558
Tam NN, Gao Y, Leung YK, Ho SM (2003) Androgenic regulation of oxidative stress in the rat prostate: involvement of NAD(P)H oxidases and antioxidant defense machinery during prostatic involution and regrowth. Am J Pathol 163: 2513–2522
Trachootham D, Alexandre J, Huang P (2009) Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nat Rev Drug Discov 8: 579–591
Vainio P, Gupta S, Ketola K, Mirtti T, Mpindi JP, Kohonen P, Fey V, Perala M, Smit F, Verhaegh G, Schalken J, Alanen KA, Kallioniemi O, Iljin K (2011) Arachidonic acid pathway members PLA2G7, HPGD, EPHX2, and CYP4F8 identified as putative novel therapeutic targets in prostate cancer. Am J Pathol 178: 525–536
van den Hoogen C, van der Horst G, Cheung H, Buijs JT, Lippitt JM, Guzman-Ramirez N, Hamdy FC, Eaton CL, Thalmann GN, Cecchini MG, Pelger RC, van der Pluijm G (2010) High aldehyde dehydrogenase activity identifies tumor-initiating and metastasis-initiating cells in human prostate cancer. Cancer Res 70: 5163–5173
Vrzal R, Ulrichova J, Dvorak Z (2004) Aromatic hydrocarbon receptor status in the metabolism of xenobiotics under normal and pathophysiological conditions. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub 148: 3–10
Wang J, Cai Y, Shao LJ, Siddiqui J, Palanisamy N, Li R, Ren C, Ayala G, Ittmann MM (2010) Activation of NF-kB by TMPRSS2/ERG fusion isoforms through Toll-like receptor-4. Cancer Res 71: 1325–1333
Wang Y (2011) Effects of salinomycin on cancer stem cell in human lung adenocarcinoma A549 cells. Med Chem 7: 106–111
Webber MM, Bello D, Kleinman HK, Hoffman MP (1997) Acinar differentiation by non-malignant immortalized human prostatic epithelial cells and its loss by malignant cells. Carcinogenesis 18: 1225–1231
Yu C, Yao Z, Dai J, Zhang H, Escara-Wilke J, Zhang X, Keller ET (2011) ALDH activity indicates increased tumorigenic cells, but not cancer stem cells, in prostate cancer cell lines. In Vivo 25: 69–76
Zhang M, Shoeb M, Goswamy J, Liu P, Xiao TL, Hogan D, Campbell GA, Ansari NH (2009a) Overexpression of aldehyde dehydrogenase 1A1 reduces oxidation-induced toxicity in SH-SY5Y neuroblastoma cells. J Neurosci Res 88: 686–694
Zhang Q, Helfand BT, Jang TL, Zhu LJ, Chen L, Yang XJ, Kozlowski J, Smith N, Kundu SD, Yang G, Raji AA, Javonovic B, Pins M, Lindholm P, Guo Y, Catalona WJ, Lee C (2009b) Nuclear factor-kappaB-mediated transforming growth factor-beta-induced expression of vimentin is an independent predictor of biochemical recurrence after radical prostatectomy. Clin Cancer Res 15: 3557–3567
Acknowledgements
We thank Finnish DNA Microarray Centre for doing the Illumina experiments and Miro Viitala for excellent technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License.
Supplementary Information accompanies the paper on British Journal of Cancer website
Supplementary information
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Ketola, K., Hilvo, M., Hyötyläinen, T. et al. Salinomycin inhibits prostate cancer growth and migration via induction of oxidative stress. Br J Cancer 106, 99–106 (2012). https://doi.org/10.1038/bjc.2011.530
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bjc.2011.530
Keywords
This article is cited by
-
Pretreatment of prostate cancer cells with salinomycin and Wnt inhibitor increases the efficacy of cabazitaxel by inducing apoptosis and decreasing cancer stem cells
Medical Oncology (2023)
-
The synergistic anticancer effect of salinomycin combined with cabazitaxel in CD44+ prostate cancer cells by downregulating wnt, NF-κB and AKT signaling
Molecular Biology Reports (2022)
-
Drug Repurposing in Medulloblastoma: Challenges and Recommendations
Current Treatment Options in Oncology (2021)
-
Vasculogenic mimicry is associated with trastuzumab resistance of HER2-positive breast cancer
Breast Cancer Research (2019)
-
Salinomycin triggers endoplasmic reticulum stress through ATP2A3 upregulation in PC-3 cells
BMC Cancer (2019)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}