
Abstract
Previous animal models and structural imaging investigations have linked hippocampal neuroplasticity to electroconvulsive therapy (ECT) response, but the relationship between changes in hippocampal volume and temporal coherence in the context of ECT response is unknown. We hypothesized that ECT response would increase both hippocampal resting-state functional magnetic resonance imaging connectivity and hippocampal volumes. Patients with major depressive disorder (n=19) were scanned before and after the ECT series. Healthy, demographically matched comparisons (n=20) were scanned at one-time interval. Longitudinal changes in functional connectivity of hippocampal regions and volumes of hippocampal subfields were compared with reductions in ratings of depressive symptoms. Right hippocampal connectivity increased (normalized) after the ECT series and correlated with depressive symptom reduction. Similarly, the volumes of the right hippocampal cornu ammonis (CA2/3), dentate gyrus and subiculum regions increased, but the hippocampal subfields were unchanged relative to the comparison group. Connectivity changes were not evident in the left hippocampus, and volume changes were limited to the left CA2/3 subfields. The laterality of the right hippocampal functional connectivity and volume increases may be related to stimulus delivery method, which was predominately right unilateral in this investigation. The findings suggested that increased hippocampal functional connectivity and volumes may be biomarkers for ECT response.
Similar content being viewed by others
Introduction
Disruptions in hippocampal structure and function have been implicated in the pathophysiology of depressive episodes. On a cellular level, depression is associated with increased hippocampal apoptosis,1 diminished neurogenesis2 and decreased number of glial cells.3 On a macroscopic level, depressive episodes are associated with diminished hippocampal volumes4, 5 and aberrant hippocampal connectivity.6 Depressive episodes also affect the inhibitory control of the hippocampus in regulating the hypothalamus–pituitary–adrenal axis, which results in excess glucocorticoid secretion.7 The cellular, structural and functional dysfunction coupled with the many hippocampal afferents and efferents support the role of the hippocampus in depression-related circuitry.8
The hippocampus has multiple roles and functions that correspond to histologically distinct anatomic subfields such as the cornu ammonis (CA; 1–3), subiculum (SUB) and dentate gyrus (DG), all of which have been implicated in major depression.9, 10 Post-mortem investigations in depression have shown smaller sizes of hippocampal pyramidal cells in the CA and increased density of neuronal and glial cell densities in both the CA and DG.10 Recently, measurement of hippocampal subfields with high-resolution structural magnetic resonance imaging has become possible,11 thereby permitting longitudinal investigations in state-related volume changes in psychiatric disorders.12
Effective antidepressant treatments may normalize aberrant depression-related hippocampal structure and function via neuroplasticity, which is defined as the brain’s capability to restructure itself by forming new neural connections.13 Neuroplasticity appears to be a common mechanism shared by both electroconvulsive therapy (ECT) and chemical antidepressant treatments, but ECT appears to be a more potent stimulator of neuroplasticity.14 Evidence from animal studies have demonstrated that a single electroconvulsive stimulation increases neurotrophic, neuropeptide and transcription factors, as well as gene expression in the hippocampus.15 Furthermore, in longitudinal neuroimaging investigations with depressed patients, ECT response is associated with increased hippocampal volumes.16, 17 Despite these findings, the relationship between structural and functional neuroplasticity changes in the context of ECT response remains unknown.
Here, we assess changes in hippocampal resting-state functional MRI (fMRI) temporal coherence as well as hippocampal subfields volumes among older, unipolar depressed subjects before and after treatment with ECT. Based on the evidence of ECT neuroplasticity, we hypothesized that ECT response would increase hippocampal resting-state fMRI functional connectivity, and that these findings would in parallel increase the hippocampal volume. We also assessed the relationship between these structural and functional changes with changes in depressive symptoms and cognitive functioning during the ECT series.
Materials and methods
Participants
The Human Research Protections Office at the University of New Mexico (UNM) approved this investigation. Patients were recruited from the UNM Mental Health Center’s inpatient and outpatient services, and age- and gender-matched healthy comparison (HC) subjects were recruited from the same demographic area. Participants had decisional capacity or assented for this investigation with a surrogate decision maker providing informed consent. Depressed subjects met the following inclusion criteria: (1) Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition, Text Revision, diagnosis of major depressive disorder; (2) clinical indications for ECT; (3) 24-item Hamilton Depression Rating Scale (HDRS-24) >21;18, 19 and (4) age >50 years. The age range was selected to minimize age-related variability and to increase the probability of ECT response with the young-old age range from 60 to 74 years of age.20 Depressed subjects were scanned within 2 days of starting the ECT series. Subjects were then scanned again at >5 days after the ECT series to avoid the immediate effect of the seizure on imaging and cognitive assessment.21, 22 HCs were scanned at one-time interval to establish a reference group for connectivity and structural changes among the ECT participants.
Clinical assessments
The primary outcome for this investigation was the change in depressive symptoms with ECT response defined as 50% reduction in baseline HDRS-24.23 Melancholic symptoms were measured pre- and post-ECT with the Hamilton Endogenomorphy Scale.24 Cognitive function was assessed pre- and post-ECT with the Repeatable Assessment for Neuropsychological Status25 and the Trail Making Tests Parts A and B.26 Because of symptom severity before ECT, only 15 depressed subjects were able to complete the neuropsychological assessment.
Demographic, clinical and neuropsychological data are summarized in Table 1. The average age for the depressed subjects (n=19) was 65.3 years±8.0 (6 males). The average age for the age- and gender-matched HCs (n=20) was 64.9±9.4 years (8 males). Eleven of the depressed subjects had a depressive episode with psychotic features. All depressed subjects were treated with antidepressant medications throughout this investigation. Concurrently, 11 subjects were treated with antipsychotics. Medication changes between the two imaging assessments were minimal and consisted of antidepressant cross-titration (n=3), antidepressant discontinuation (n=1) and the addition of an antipsychotic (olanzapine, n=1).
ECT procedure
A Thymatron System IV (Somatics, Lake Bluff, IL, USA) delivered a right unilateral (n=17) or bitemporal (n=2) stimulus based on clinical indication at the start of the ECT series. Seizure threshold was obtained during the first session, while a dose titration method guided subsequent stimulus dosage at 6 × threshold for right unilateral and 2 × threshold for bitemporal. Further adjustments to energy occurred as needed for inadequate seizure duration, defined as <25 s of electroencephalogram seizure activity. Treatments occurred three times weekly until adequate clinical response or clinical decision to stop treatment for non-responders. Anesthetic management included methohexital followed by succinylcholine for muscle relaxation. Patients were oxygenated throughout the procedure with a disposable bag and mask. Blood pressure, pulse and oxygen saturations were monitored throughout the procedure.
Image acquisition
Structural magnetic resonance imaging data were collected on a 3-Tesla Siemens Trio scanner (Siemens Healthcare, Malvern, PA, USA). Anatomic T1-weighted multi-echo MPRAGE sequence were collected with the following parameters: Repetiton time (TR)=2.53 s, echo time (TE)=1.64, 3.5, 5.32, 7.22, 9.08 ms, inversion time (TI)=1.2 s, flip angle=7, number of excitations=1, slice thickness=1 mm, field of view=256 mm, matrix 256 × 256 and voxel size=(1 × 1 × 1 mm). A whole-brain gradient-echo echoplanar imaging sequence were collected with the following parameters: TR=2 s, TE=29 ms, flip angle 75, slice thickness=3.5 mm, slice gap=1.05 mm, field of view 240 mm, matrix 64 × 64 and voxel size=3.75 × 3.75 × 4.55 mm. Functional images consisted of 154 volumes. Subjects were instructed to keep their eyes open during the scan and stare passively at a fixation cross with continuous monitoring of eye movements throughout the scan interval.
Volume analysis
Structural magnetic resonance imaging preprocessing and the delineation of structural images were conducted using FreeSurfer 4.5.0 software (https://surfer.nmr.mgh.harvard.edu).27 In brief, after removal of non-brain tissue via a hybrid watershed algorithm/surface deformation, the image then underwent an automated Talairach transformation and whole-brain segmentation. Gray matter was further segmented with a probabilistic brain atlas. The hippocampal subfields were based upon additional Bayesian modeling and a probabilistic atlas of a region of interest surrounding the hippocampal formation (94 × 66 × 144 voxels).11 The larger hippocampal subfields, including CA2/3, DG and SUB, have been shown to correlate well with manual tracings in previous studies.11
Resting-state fMRI image analysis
fMRI preprocessing was conducted with Statistical Parametric Mapping 8 (http://www.fil.ion.ucl.ac.uk/spm/software/spm8). Resting-state fMRI data were realigned with INRIalign and slice-timing correction was applied with the middle slice as the reference frame. Data were then spatially normalized into the standard Montreal Neurological Institute space, resliced to 3 × 3 × 3 mm voxels and smoothed using a Gaussian kernel with a full-width at half-maximum of 10 mm to account for the greater anatomic variability in the older sample.28 Signal from motion, cerebral spinal fluid and white matter was removed with tCompCor, a component-based noise correction method that removes voxels with high s.d. (0>0.98).29 The data were despiked (analysis of functional neuroimages 3dDespike (http://afni.nimh.nih.gov)) and then were band-pass filtered (0.008–0.09 Hz).
Study-specific mask hippocampal mask and functional connectivity analysis
Previous research has shown that hippocampal atrophy confounds standardized template-based methods by over-sampling surrounding areas such as the parahippocampal gyrus.30 Therefore, we created a study-specific hippocampal mask from the Freesurfer volumes. Each subject’s T1 structural image and segmented Freesurfer volume was warped with the Montreal Neurological Institute template. The warped hippocampal masks were then summed and eroded by dropping voxels where <90% of the subjects contributed to the summed hippocampal mask. The mask was then resliced to 3 × 3 × 3 mm voxels. The final mask was the intersection of the subject-specific mask and the Wake Forest University Pick-Atlas hippocampal mask. The average time courses for the right and left hippocampal masks (39 voxels each) generated correlation maps that were then r- to z-transformed.
Statistical analysis
Based on previous research,31 structural and connectivity changes were limited to ECT responders only (n=15). The HC group was scanned at one-time interval and precluded an interaction analysis. For group-level structural and functional connectivity analyses, we performed the following analyses: (1) between-subjects comparisons between the pre-ECT depression group and HC to establish pathology (two-sample t-tests); (2) longitudinal assessments with the depression group before and after ECT (paired t-tests); and (3) between-subjects comparisons between the post-ECT depression group and HC to assess for normalization of aberrant connectivity or volumes (two-sample t-tests). For group-level whole-brain functional connectivity analysis, statistical images were assessed for cluster-wise significance using a cluster-defining threshold of P=0.005. The 0.05 family-wise error critical cluster size was 321 voxels. This liberal cluster-defining threshold increased the sensitivity for detecting functional connectivity differences associated with ECT response while still being fairly focal and not having the problems associated with spanning many different brain regions.32 For hippocampal structural results, we reported on absolute and normalized volumes with respect to intracranial volume. For hippocampal subfields, we reported on absolute volumes only, as the volume determination was based on a region of interest surrounding the hippocampal formation. We compared changes in volume and connectivity with changes in symptoms among the entire sample of depressed subjects (n=19). Correlations with the entire data set of ECT participants increased the range of response in depressive symptoms to empirically determine whether functional and structural changes were related to clinical change. We also compared changes in cognitive performance among the entire sample of depressed subjects with a completed neuropsychological assessment (n=15). Because of the focus on the hippocampus, we used change in percent recall as the primary assessment of hippocampal cognitive performance.33
Results
Clinical
ECT participants received 11±2.7 treatments within the ECT series. The post-ECT HDRS-24 confirmed clinical response from a pre-ECT assessment of 32.6±8.5 to a post-ECT assessment of 8.4±8.6. Fifteen of the 19 ECT subjects were ECT responders with an average age 65.5±6.7 years (5 males). The neuropsychological performance was notable for an improvement in immediate memory (t14=−2.15, P=0.04) and percent retention (t14=−2.2, P=0.04), which may be owing to the length of time (11±8.4 days) between the last ECT session and the post-ECT assessment.21
Pre-ECT and HC contrasts
Structural results
The right and left hippocampal volumes and subfields were not significantly different between the pre-ECT responders (n=15) and HC groups (P>0.10).
fMRI results
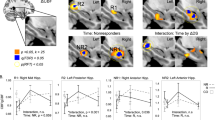
ECT responders (n=15) had lower right hippocampal connectivity in two clusters relative to HC: the left medial temporal lobe (Pfamilywise error (FWE)=0.008, KE=455 and local maxima: −51, −31 and −17) and the anterior cingulate cortex (PFWE=0.005, KE=502 and local maxima: −6, 26 and 13). ECT responders also had lower left hippocampal connectivity in two clusters in the left temporal lobe in Brodmann area 21 (PFWE=0.021, KE=377 and local maxima: −57, −1 and −8; PFWE=0.007, KE=479 and local maxima: −36, −67 and 16) and another cluster in the right middle temporal lobe (PFWE=0.014, KE=417 and local maxima: 48, −25 and −5). The differences in right and left hippocampal connectivity are shown in Figure 1. ECT responders did not have any areas of increased connectivity relative to HC (P>0.10).

The healthy comparison (HC) >pre-electroconvulsive therapy (ECT) contrast shows decreased left hippocampal connectivity in the left and right temporal lobe. Similarly, the HC >pre-ECT contrast shows decreased right hippocampal connectivity in the left medial temporal and anterior cingulate cortex. Differences in connectivity are mapped to color hue; t-statistic magnitude is mapped to transparency. White contour lines denote familywise error-corrected areas of significance with a cluster-defining threshold of P=0.005.
Pre- and post-ECT contrasts
Structural results
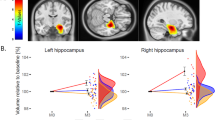
Right hippocampal normalized volumes (t14=−2.7, P=0.01) increased after the ECT series relative to pre-ECT scans among ECT responders. The left hippocampal volumes did not increase in size (P>0.10). Right hippocampal subfield volumes also increased in the right CA1 (t14=−2.6, P=0.02), CA2/3 (t14=−4.1, P=0.001), DG (t14=−2.4, P=0.03) and SUB (t14=−2.8, P=0.01). Increased volumes in the left hippocampus were limited to the CA2/3 (t14=−2.6, P=0.02) subfield as shown in Table 2.
fMRI results
Right hippocampal connectivity increased in the right temporal lobe, (PFWE =0.025, KE=307 and local maxima: 42, −22 and −11) post-ECT as shown in Figure 2 among ECT responders. The left hippocampus did not show a similar increase in connectivity (P>0.10). Right and left hippocampal connectivity did not decrease during the ECT series (P>0.10).

The post-electroconvulsive therapy (ECT) >pre-ECT contrast shows no significant differences with left hippocampal connectivity. The post-ECT >pre-ECT contrast shows increased right hippocampal connectivity among ECT responders in the left superior temporal lobe. The black contour lines denote the study-specific right hippocampal seed region.
Post-ECT and HC contrasts
Structural results
The right and left hippocampal volumes and subfields were not significantly different between the post-ECT responders and HC groups (P>0.10).
fMRI results
The right and left hippocampal connectivity maps were not significantly between post-ECT depressed subjects and HC groups (P>0.10), thereby providing confirmatory evidence of normalization.
Correlations with symptoms, cognitive performance and data integration
Structural results
The change in symptoms or cognitive performance did not correlate with change in hippocampal volumes (P>0.10) across all patients (n=15).
fMRI results
Using a mask of the area of increased right hippocampal connectivity, we assessed the relationship between changes in connectivity, symptoms and neuropsychological performance. A positive relationship (r=0.52, P=0.02) existed between differences in connectivity and change in depressive symptomatology among all ECT participants (n=19) as shown in Figure 3. The relationship between changes in connectivity and cognition, as measured by differences in percent retention, was not significant (P>0.10).

The scatter plot and linear fit show the relationship between differences in connectivity and symptom change among all electroconvulsive therapy (ECT) patients (n=19, r=0.52, P=0.024). The red line represents the linear fit and the shaded region represents 95% confidence intervals.
Data integration
Correlations between right hippocampal volume changes and right hippocampal functional connectivity changes (assessed with the mask of the area of increased right hippocampal connectivity) were not significant among all ECT participants (P>0.10).
Discussion
Consistent with our hypothesis, ECT response is associated with increased hippocampal functional connectivity and increased volume. Before the ECT series, the ECT responders had decreased right and left hippocampal connectivity in the anterior cingulate and temporal cortices relative to the HC reference group, which confirmed aberrant, bilateral hippocampal connectivity among depressed patients before the ECT series. ECT response was associated with increased right hippocampal connectivity, which also correlated with reduction in depressive symptoms. Among ECT responders, the aberrant connectivity normalized after the ECT series, and the aberrant pre-ECT connectivity differences were no longer evident relative to the HC reference group. With respect to the structural MRI findings, the hippocampal volumes and subfields between ECT responders and HCs before or after the ECT series were not statistically different, but the longitudinal analysis demonstrated increased right hippocampal and right hippocampal subfield volume increases associated with ECT response. The functional connectivity results did not correlate with the volumetric changes, but may have captured complementary neuroplastic changes relevant to the mechanisms of action of ECT. In the remaining section, we discuss the laterality of our findings and the possible mechanisms of ECT-induced structural and functional changes.
Changes in the laterality of the structural and functional changes were limited to the right hippocampus, which corresponded to the predominate side of stimulation. Previous hippocampal volumetric investigations have found changes that may be related to method of stimulus delivery. For example, left hippocampal volumetric increases have been found with ECT provided with bitemporal electrode placement.34 Right hippocampal volumetric increases have been found with right unilateral electrode placement.17 The increased hippocampal gray matter volume from the latter study closely mirrors the increased connectivity found with our investigation, which also used a right unilateral electrode placement. Computer modeling has shown that the method of electrode placement affects the location of current density, with right unilateral being more limited relative to bitemporal.35, 36 The unilateral correspondence between side of stimulation and functional/structural changes increases the likelihood that observed findings are a direct result of ECT treatment. A recent, longitudinal ECT investigation has provided further support for changes in connectivity in relation to electrode placement.37
Mechanisms of ECT-related neural plasticity include neurogenesis, synaptogenesis and gliogenesis (for recent review, see Bouckaert et al.13). The neurogenic hypothesis of depression posits that new neurons in the adult brain are needed for proper mood control and for antidepressant efficacy.38 The association between age-related decreases in neurogenesis and the increased prevalence of depression with age provides further support for this hypothesis.39, 40 In our results, neurogenesis may be supported by increased volume of the DG, a ‘neurogenic niche’ within the hippocampus.40 Speculations about structural changes related to neurogenesis must be balanced with the time frame (>6 months) and the likely amount of neurogenesis (between 1 and 30% of total granule cells) relative to substantial macroscopic volume changes observed in our investigation.41, 42
The structural and functional changes related to ECT are not limited to neurogenesis and could be explained by other mechanisms.43 Indeed, antidepressant volumetric changes related to vasculature, astrocytes and microglia are neurogenesis independent.44 The increased temporal coherence of functional connectivity results may also be suggestive of synaptogenesis of pre-existing neurons and is further supported by increased volume of the SUB, the primary location of hippocampal afferents with monosynaptic connections to subcortical (amygdala and nucleus accumbens) and cortical (frontal, cingulate and parietal) nuclei.9 The hippocampal volumes differences (pre-/post-ECT contrast) are relatively small (121.2 mm3 or 4.5 fMRI voxels) relative to the volume of increased fMRI connectivity (321 voxels). Furthermore, we did not find any significant correlations between structural and functional changes, which may suggest complementary but different processes of volume and connectivity increases (that is, neurogenesis and gliogenesis for volume increases and synaptogenesis for functional connectivity increases). Although we did not find a direct relationship between structural and functional changes, future investigations will elucidate more complex functional relationships mediated by indirect structural connections with advanced analytical methods such as data fusion.45, 46 Other controversial explanations for longitudinal changes in structural changes include confounding factors such as hydration, cardiac pulsation, B1 in homogeneities and subject motion.47, 48 These confounds caution against definitive, biological interpretations regarding neural plasticity.
Several limitations of this investigation should be acknowledged when interpreting our results, and most are owing to the clinical nature of the study. First, all of the ECT subjects were medicated. Concurrent antidepressant treatment will have synergy with ECT49 and may share a similar but less potent mechanism of action for clinical outcome,14 which may have impacted our results. Discontinuation of antidepressant therapy before the first imaging assessment was not feasible (vast majority of subjects were inpatient at the start of this investigation) and may have introduced additional confounds such as discontinuation syndrome.50 However, we limited cross-titrations to a small minority (~15%) of ECT participants. Second, our small sample of unipolar depressed patients had both psychotic and non-psychotic episodes, and the study was underpowered to compare these groups. In future studies with larger samples, we will assess the effect of psychosis and confirm the relationship between connectivity and symptom changes. Third, hippocampal subfields were assessed with standard T1 structural images (1 × 1 × 1 mm), which may have limited contrast and challenge template-based registration methods to reliably segment hippocampal subfields.51 However, Freesurfer segmentation and standard T1 structural images have previously demonstrated differences in hippocampal subfields in cross-sectional52 and longitudinal studies12 in patients with neuropsychiatric disorders.
Conclusion
The unilateral nature of our findings suggests that the concomitant structural and functional changes associated with ECT response are mechanistic in nature and extend previous animal and clinical investigations on the role of hippocampal plasticity with ECT response. Importantly, the current study is the first to demonstrate that ECT induces normalization of aberrant hippocampal connectivity and increased hippocampal volumes in the context of a clinical response (decreased depressive symptomatology). Current and previous findings highlight the role that the hippocampus has in the successful remediation of depressive symptomatology with ECT, and the role of neuroimaging in revealing potential mechanisms of therapeutic efficacy. These results will provide the framework for longitudinal investigations that will assess the permanence (or impermanence53) of hippocampal functional and structural changes in the context of sustained response and relapse.
References
Lucassen PJ, Muller MB, Holsboer F, Bauer J, Holtrop A, Wouda J et al. Hippocampal apoptosis in major depression is a minor event and absent from subareas at risk for glucocorticoid overexposure. Am J Pathol 2001; 158: 453–468.
Gould E, Tanapat P, McEwen BS, Flugge G, Fuchs E . Proliferation of granule cell precursors in the dentate gyrus of adult monkeys is diminished by stress. Proc Natl Acad Sci USA 1998; 95: 3168–3171.
Sahay A, Drew MR, Hen R . Dentate gyrus neurogenesis and depression. Prog Brain Res 2007; 163: 697–722.
Steffens DC, Byrum CE, McQuoid DR, Greenberg DL, Payne ME, Blitchington TF et al. Hippocampal volume in geriatric depression. Biol Psychiatry 2000; 48: 301–309.
Sheline YI, Wang PW, Gado MH, Csernansky JG, Vannier MW . Hippocampal atrophy in recurrent major depression. Proc Nati Acad Sci USA 1996; 93: 3908–3913.
Tahmasian M, Knight DC, Manoliu A, Schwerthoffer D, Scherr M, Meng C et al. Aberrant intrinsic connectivity of hippocampus and amygdala overlap in the fronto-insular and dorsomedial-prefrontal cortex in major depressive disorder. Front Hum Neurosci 2013; 7: 639.
Zhu LJ, Liu MY, Li H, Liu X, Chen C, Han Z et al. The Different Roles of Glucocorticoids in the Hippocampus and Hypothalamus in Chronic Stress-Induced HPA Axis Hyperactivity. PloS One 2014; 9: e97689.
Seminowicz DA, Mayberg HS, McIntosh AR, Goldapple K, Kennedy S, Segal Z et al. Limbic-frontal circuitry in major depression: a path modeling metanalysis. NeuroImage 2004; 22: 409–418.
Small SA, Schobel SA, Buxton RB, Witter MP, Barnes CA . A pathophysiological framework of hippocampal dysfunction in ageing and disease. Nat Rev 2011; 12: 585–601.
Stockmeier CA, Mahajan GJ, Konick LC, Overholser JC, Jurjus GJ, Meltzer HY et al. Cellular changes in the postmortem hippocampus in major depression. Biol Psychiatry 2004; 56: 640–650.
Van Leemput K, Bakkour A, Benner T, Wiggins G, Wald LL, Augustinack J et al. Automated segmentation of hippocampal subfields from ultra-high resolution in vivo MRI. Hippocampus 2009; 19: 549–557.
Kuhn S, Charlet K, Schubert F, Kiefer F, Zimmermann P, Heinz A et al. Plasticity of hippocampal subfield volume cornu ammonis 2+3 over the course of withdrawal in patients with alcohol dependence. JAMA Psychiatry 2014; 71: 806–811.
Bouckaert F, Sienaert P, Obbels J, Dols A, Vandenbulcke M, Stek M et al. ECT: its brain enabling effects: a review of electroconvulsive therapy-induced structural brain plasticity. J ECT 2014; 30: 143–151.
Malberg JE, Eisch AJ, Nestler EJ, Duman RS . Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J Neurosci 2000; 20: 9104–9110.
Segi-Nishida E . Exploration of new molecular mechanisms for antidepressant actions of electroconvulsive seizure. Biol Pharma Bull 2011; 34: 939–944.
Nordanskog P, Dahlstrand U, Larsson MR, Larsson EM, Knutsson L, Johanson A . Increase in hippocampal volume after electroconvulsive therapy in patients with depression: a volumetric magnetic resonance imaging study. J ECT 2010; 26: 62–67.
Dukart J, Regen F, Kherif F, Colla M, Bajbouj M, Heuser I et al. Electroconvulsive therapy-induced brain plasticity determines therapeutic outcome in mood disorders. Proc Natl Acad Sci USA 2014; 111: 1156–1161.
Kellner CH, Knapp RG, Petrides G, Rummans TA, Husain MM, Rasmussen K et al. Continuation electroconvulsive therapy vs pharmacotherapy for relapse prevention in major depression: a multisite study from the Consortium for Research in Electroconvulsive Therapy (CORE). Arch Gen Psychiatry 2006; 63: 1337–1344.
Hamilton M . Development of a rating scale for primary depressive illness. Br J Soc Clin Psychol 1967; 6: 278–296.
Tew JD Jr ., Mulsant BH, Haskett RF, Prudic J, Thase ME, Crowe RR et al. Acute efficacy of ECT in the treatment of major depression in the old-old. Am J Psychiatry 1999; 156: 1865–1870.
Semkovska M, McLoughlin DM . Objective cognitive performance associated with electroconvulsive therapy for depression: a systematic review and meta-analysis. Biol Psychiatry 2010; 68: 568–577.
Schmidt EZ, Reininghaus B, Enzinger C, Ebner C, Hofmann P, Kapfhammer HP . Changes in brain metabolism after ECT-positron emission tomography in the assessment of changes in glucose metabolism subsequent to electroconvulsive therapy—lessons, limitations and future applications. J Affect Disord 2008; 106: 203–208.
Husain MM, Rush AJ, Fink M, Knapp R, Petrides G, Rummans T et al. Speed of response and remission in major depressive disorder with acute electroconvulsive therapy (ECT): a Consortium for Research in ECT (CORE) report. J Clin Psychiatry 2004; 65: 485–491.
Thase ME, Hersen M, Bellack AS, Himmelhoch JM, Kupfer DJ . Validation of a Hamilton subscale for endogenomorphic depression. J Affect Disord 1983; 5: 267–278.
Randolph C, Tierney MC, Mohr E, Chase TN . The repeatable battery for the assessment of neuropsychological status (RBANS): preliminary clinical validity. J Clin Exp Neuropsychol 1998; 20: 310–319.
Reitan RM . Validity of the Trail Making test as an indicator of organic brain damage. Percept Mot Skills 1958; 8: 271–276.
Fischl B, Salat DH, Busa E, Albert M, Dieterich M, Haselgrove C et al. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron 2002; 33: 341–355.
Aizenstein HJ, Andreescu C, Edelman KL, Cochran JL, Price J, Butters MA et al. fMRI correlates of white matter hyperintensities in late-life depression. Am J Psychiatry 2011; 168: 1075–1082.
Behzadi Y, Restom K, Liau J, Liu TT . A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 2007; 37: 90–101.
Sandstrom CK, Krishnan S, Slavin MJ, Tran TT, Doraiswamy PM, Petrella JR . Hippocampal atrophy confounds template-based functional MR imaging measures of hippocampal activation in patients with mild cognitive impairment. Am J Neuroradiol 2006; 27: 1622–1627.
Abbott CC, Jones TR, Bustillo J, Calhoun VD, Turner J . Resting State fMRI Neural Correlates of ECT Response in Major Depressive Disorder. Translational Science: Washington DC, USA, 2013.
Woo CW, Krishnan A, Wager TD . Cluster-extent based thresholding in fMRI analyses: pitfalls and recommendations. NeuroImage 2014; 91: 412–419.
Hickie I, Naismith S, Ward PB, Turner K, Scott E, Mitchell P et al. Reduced hippocampal volumes and memory loss in patients with early- and late-onset depression. Br J Psychiatry 2005; 186: 197–202.
Tendolkar I, van Beek M, van Oostrom I, Mulder M, Janzing J, Voshaar RO et al. Electroconvulsive therapy increases hippocampal and amygdala volume in therapy refractory depression: a longitudinal pilot study. Psychiatry Res 2013; 214: 197–203.
Lee WH, Deng ZD, Kim TS, Laine AF, Lisanby SH, Peterchev AV . Regional electric field induced by electroconvulsive therapy in a realistic finite element head model: influence of white matter anisotropic conductivity. NeuroImage 2012; 59: 2110–2123.
Peterchev AV, Rosa MA, Deng ZD, Prudic J, Lisanby SH . Electroconvulsive therapy stimulus parameters: rethinking dosage. J ECT 2010; 26: 159–174.
van Waarde JA, Scholte HS, van Oudheusden LJ, Verwey B, Denys D, van Wingen GA . A functional MRI marker may predict the outcome of electroconvulsive therapy in severe and treatment-resistant depression. Mol Psychiatry advance online publication, 5 August 2014; doi: 10.1038/mp.2014.78 (e-pub ahead of print).
Petrik D, Lagace DC, Eisch AJ . The neurogenesis hypothesis of affective and anxiety disorders: are we mistaking the scaffolding for the building? Neuropharmacology 2012; 62: 21–34.
Lazarov O, Mattson MP, Peterson DA, Pimplikar SW, van Praag H . When neurogenesis encounters aging and disease. Trends Neurosci 2010; 33: 569–579.
Eisch AJ, Petrik D . Depression and hippocampal neurogenesis: a road to remission? Science 2012; 338: 72–75.
Kohler SJ, Williams NI, Stanton GB, Cameron JL, Greenough WT . Maturation time of new granule cells in the dentate gyrus of adult macaque monkeys exceeds six months. Proc Natl Acad Sci USA 2011; 108: 10326–10331.
Amrein I, Isler K, Lipp HP . Comparing adult hippocampal neurogenesis in mammalian species and orders: influence of chronological age and life history stage. Eur J Neurosci 2011; 34: 978–987.
Ho NF, Hooker JM, Sahay A, Holt DJ, Roffman JL . In vivo imaging of adult human hippocampal neurogenesis: progress, pitfalls and promise. Mol Psychiatry 2013; 18: 404–416.
David DJ, Samuels BA, Rainer Q, Wang JW, Marsteller D, Mendez I et al. Neurogenesis-dependent and -independent effects of fluoxetine in an animal model of anxiety/depression. Neuron 2009; 62: 479–493.
Sui J, He H, Yu Q, Chen J, Rogers J, Pearlson GD et al. Combination of Resting State fMRI, DTI, and sMRI Data to Discriminate Schizophrenia by N-way MCCA+jICA. Front Hum Neurosci 2013; 7: 235.
Damoiseaux JS, Greicius MD . Greater than the sum of its parts: a review of studies combining structural connectivity and resting-state functional connectivity. Brain Struct Funct 2009; 213: 525–533.
Thomas C, Baker CI . Teaching an adult brain new tricks: a critical review of evidence for training-dependent structural plasticity in humans. NeuroImage 2013; 73: 225–236.
Thomas C, Baker CI . On evidence, biases and confounding factors: response to commentaries. NeuroImage 2013; 73: 265–267.
Sackeim HA, Dillingham EM, Prudic J, Cooper T, McCall WV, Rosenquist P et al. Effect of concomitant pharmacotherapy on electroconvulsive therapy outcomes: short-term efficacy and adverse effects. Arch Gen Psychiatry 2009; 66: 729–737.
Abbott CC, Gallegos P, Rediske N, Lemke NT, Quinn DK . A review of longitudinal electroconvulsive therapy: neuroimaging investigations. J Geriatr Psychiatry Neurol 2014; 27: 33–46.
Yushkevich PA, Wang H, Pluta J, Das SR, Craige C, Avants BB et al. Nearly automatic segmentation of hippocampal subfields in in vivo focal T2-weighted MRI. NeuroImage 2010; 53: 1208–1224.
Ezzati A, Zimmerman ME, Katz MJ, Sundermann EE, Smith JL, Lipton ML et al. Hippocampal subfields differentially correlate with chronic pain in older adults. Brain Res 2014; 1573: 54–62.
Nordanskog P, Larsson MR, Larsson EM, Johanson A . Hippocampal volume in relation to clinical and cognitive outcome after electroconvulsive therapy in depression. Acta Psychiatr Scand 2014; 129: 303–311.
Acknowledgements
The Dana Foundation Brain and Immuno-imaging, and Centers of Biomedical Research Excellence (2P20GM103472-01) supported this investigation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Abbott, C., Jones, T., Lemke, N. et al. Hippocampal structural and functional changes associated with electroconvulsive therapy response. Transl Psychiatry 4, e483 (2014). https://doi.org/10.1038/tp.2014.124
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/tp.2014.124
This article is cited by
-
How electroconvulsive therapy works in the treatment of depression: is it the seizure, the electricity, or both?
Neuropsychopharmacology (2024)
-
Transcranial direct current stimulation (tDCS) in depression induces structural plasticity
Scientific Reports (2023)
-
Cerebro-cerebellar functional neuroplasticity mediates the effect of electric field on electroconvulsive therapy outcomes
Translational Psychiatry (2023)
-
Cortical excitation/inhibition ratios in patients with major depression treated with electroconvulsive therapy: an EEG analysis
European Archives of Psychiatry and Clinical Neuroscience (2023)
-
Links between electroconvulsive therapy responsive and cognitive impairment multimodal brain networks in late-life major depressive disorder
BMC Medicine (2022)
