
Abstract
The Mediterranean Sea has been defined “under siege” because of intense pressures from multiple human activities; yet there is still insufficient information on the cumulative impact of these stressors on the ecosystem and its resources. We evaluate how the historical (1950–2011) trends of various ecosystems groups/species have been impacted by changes in primary productivity (PP) combined with fishing pressure. We investigate the whole Mediterranean Sea using a food web modelling approach. Results indicate that both changes in PP and fishing pressure played an important role in driving species dynamics. Yet, PP was the strongest driver upon the Mediterranean Sea ecosystem. This highlights the importance of bottom-up processes in controlling the biological characteristics of the region. We observe a reduction in abundance of important fish species (~34%, including commercial and non-commercial) and top predators (~41%), and increases of the organisms at the bottom of the food web (~23%). Ecological indicators, such as community biomass, trophic levels, catch and diversity indicators, reflect such changes and show overall ecosystem degradation over time. Since climate change and fishing pressure are expected to intensify in the Mediterranean Sea, this study constitutes a baseline reference for stepping forward in assessing the future management of the basin.
Similar content being viewed by others


Introduction
Marine ecosystems around the world are increasingly pressured by a diversity of anthropogenic stressors, which include intensive fisheries and aquaculture, pollution, habitat loss and degradation, and species invasions1,2. These are acting synergistically with global climate change3. Since human stressors change over time4, the assessment of their temporal cumulative effects has been poorly studied and remains a challenging task2. Because these stressors are rapidly increasing, understanding how human interactions, the environment, and marine species interact and influence each other, and how such dynamics affect the sustainability of goods and services they provide, is of urgent importance. Currently this is a priority of many national and international regulations/initiatives (e.g., European Marine Strategy Framework Directive [MSFD; 2008/56/EC]; Convention of Biological Diversity, [CBD], Intergovernmental Platform on Biodiversity and Ecosystem Services [IPBES]) which promote the preservation of natural ecosystems and a sustainable use of biodiversity resources.
In support of these regulations, new comprehensive scientific tools have been developed with the goal of integrating the effects of the above-mentioned stressors into common frameworks in order to assist policy decisions1,5,6. Particularly in the context of ecosystem-based management approach (EBM), which assesses ecosystem dynamics rather than evaluating single resources and single threats, there has been a growing use of ecosystem models. These tools are improving their ability to predict complex system dynamics considering the impact of multiple pressures7 and assessing different policy objectives sought by management authorities6,8,9. Through hind-cast and forecast scenarios, ecosystem models allow for the quantitative assessment of the role of different stressors on ecosystem dynamics and for the calculation of model-based indicators able to evaluate whether an ecosystem and its services are maintained and used sustainably. Model-based indicators can complement data-based indicators10 and have been developed and widely used to capture the impact of specific pressures on marine ecosystems11,12, such as fishing, eutrophication. More recently, they have been utilized to assess socio-economic and governance issues13,14, as well as the cumulative impacts of multiple human activities12,15, informing management processes9,16.
This study applies the Ecopath with Ecosim (EwE) food web model approach to the Mediterranean Sea ecosystem as a whole, with the aim to evaluate temporal responses of species abundances and ecosystem dynamics to changes in primary productivity and fisheries (and their combination). The Mediterranean Sea is a highly diverse marine ecosystem that hosts 7–10% of the world’s marine biodiversity17,18. It is defined as “under siege” due to historical and current impacts of multiple stressors19. Among them, fishing practises, habitat loss and degradation, eutrophication, and more recently, the introduction of alien species and climate change effects17,19,20. Since the intensity of these stressors is increasing throughout most of the Mediterranean basin, temporal analyses are increasingly needed to inform effective current and future marine policies and management actions. In this study, we first quantify temporal dynamics of marine species in the Mediterranean Sea ecosystem as a whole, evaluating their historical dynamics. We then calculate a series of ecological indicators to analyse past ecosystem dynamics. Using the temporal-dynamic module of EwE, our specific goals are to investigate: 1) the temporal evolution of the Mediterranean marine ecosystem from 1950 to 2011 by developing a hind-cast scenario analysis that includes primary productivity, fisheries activities and food web dynamics; 2) differences and similarities in historical ecosystem dynamics using the primary productivity calculated by the EwE food web model and the primary productivity obtained from an hydrodynamic-biogeochemical model (GETM-MedERGOM); and 3) the structural and functional historical changes of the Mediterranean Sea ecosystem using specific model-based indicators.
In particular, the reason why we assessed temporal ecosystem dynamics using first the primary production from the high trophic level model, EwE, and then the primary production from the low trophic level model GETM-MedERGOM (n. 2 objective) was to evaluate if results from the modelling framework together (GETM-MedERGOM-EwE) were able to better capture the temporal evolution of the Mediterranean food web while accounting for dynamic forcing effects of climate and anthropogenic impacts (e.g., fishing)11. In recent years, under the growing need to provide guidance for biodiversity conservation and ecosystem based management, these so called end-to-end (E2E) models5,21 have been increasingly used to predict marine ecosystem changes and for scenario testing of climate change and anthropogenic impacts simultaneously22,23.
Thus, studies like the present are informative to support and implement European policies such as the Marine Strategy Framework Directive (MSFD; 2008/56/EC) that requires the development of strategies to achieve a “Good Environmental Status (GES)”. They can also support regional policies like the UNEP’s Mediterranean Action Plan (MAP) that aims at moving towards an ecosystem based management approach (EBM) for both EU and non-EU Mediterranean countries. The present study sets a baseline to further develop ecosystem analyses in order to facilitate the implementation of management policies and explore future plausible scenarios.
Materials and Methods
The baseline food web model of the Mediterranean Sea
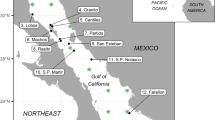
We used a previously developed food web model24 constructed with the Ecopath with Ecosim approach (EwE) using the Ecopath mass-balance module25 representing the whole Mediterranean ecosystem in the 1950 decade. We used the Ecopath model as a baseline to run temporal hind-cast (1950–2011) analyses, assessing the response of the Mediterranean marine ecosystem to changes in primary productivity and fishing effort. The baseline Ecopath model consisted of 103 functional groups, ranging from phytoplankton and invertebrates to top predator species and it was divided in four sub-models representing the four main areas of the Mediterranean basin: 1) Western Mediterranean Sea (W); 2) Adriatic Sea (A); 3) Ionian and Central Mediterranean Sea (I); and 4) Aegean Sea and Levantine Sea (E). These areas account for sub-regional differences in environmental and ecological characteristics of the ecosystem and are utilised also by the Marine Strategy Framework Directive (MSFD; Fig. 1).

Map was originated using the GIS (Geographical Information System; ArcMap version 10.3; www.esri.com) software.
The food web model had the following key input variables: biomass (B), production/biomass ratio (P/B), consumption/biomass ratio (Q/B), diet composition, and fisheries catches and discards. The main trophic structure of the Mediterranean Sea EwE model is shown in Fig. 2 and species and/or functional groups included in the model are listed in Table S1. A full description and sources of information of the input and output parameters of the baseline Ecopath model are available in Piroddi et al.24 and are presented in S2-S3 Tables in the Supporting Information.

Each functional group is shown as a circle and its size is proportional to the log of its biomass. The functional groups are represented by their trophic levels (y-axis) and linked by predator-prey relationships showed as light grey lines. Numbers refer to functional group codes, which are reported in the legend, while those in red are graphically represented with a drawing. Numbers in the figure: 1. Piscivorous cetaceans; 2. Other cetaceans; 3. Pinnipeds; 4. Seabirds; 5. Sea turtles; 6. Large pelagics; 7. Medium pelagics; 8. European pilchard; 9. European anchovy; 10. Other small pelagics; 11. Large demersals; 12. European hake; 13. Medium demersals; 14. Small demersals; 15. Deep sea fish; 16. Sharks; 17. Rays and skates; 18. Benthopelagic cephalopods; 19. Benthic cephalopods; 20. Bivalves and gastropods; 21. Crustaceans; 22. Jellyfish; 23. Benthos; 24. Zooplankton; 25. Phytoplankton; 26. Seagrass; 27. Detritus; 28. Discards. Drawings for species/functional groups 1–3, 5–6 and 8 are by Massimo Demma - http://www.massimodemma.it/-.
A set of pre-balancing (PREBAL26) analyses are presented in Figure S4 with the purpose of showing the coherency of the basic input parameters with respect to general rules/principles of ecosystem ecology. In particular, these rules include: 1. biomass estimates by functional group in the model, which span 5–7 orders of magnitude when arranged against their trophic levels; 2. slope of biomass (on a log scale) by functional group, which declines by 5–10% across all the taxa when arranged against trophic levels; 3. vital rates (P/B; Q/B) across taxa/trophic levels which decline with increasing trophic level26,27.
The Ecopath model constructed in Piroddi et al.24 included seven types of fishing fleets: trawlers, dredges, mid water trawlers, purse seiners, longliners, artisanal and recreational activities. In this study, these fleets were adapted due to a lack of time series of data regarding the number of vessels and gross tonnage (GT) for some of the fleets, which are important for estimating historical fishing effort. In particular, while recreational fishery was retained from the previous model, main commercial fisheries were divided in: 1. trawlers (which included trawlers and dredges); 2. purse seiners; 3. longliners and 4. artisanal fisheries. This new fishing fleets configuration was created to follow the same structure as in Sacchi et al.28, the main source of information for temporal time series data of number of vessels and gross tonnage (GT) for the above-mentioned fleets for each Mediterranean country for the period 1990–2010. For Italy and Greece, we were able to get longer time series data using detailed reconstructions respectively from Piroddi et al.24 for the 1950–2010 period, from Stergiou et al.29 for 1964–1989, and Moutopoulos et al.30 for 1990–2010. To estimate an overall trend of number of fishing vessels for the 1950–2010 period, for those countries with missing years, we assumed same trends as observed by Greer31 who reported the number of fishing vessels for each country of the world for 1950–2010. GT was extrapolated, for the missing years, as the average ratio of GT in the observed time periods, while number of days spent fishing were kept as the ratio of days at sea observed respectively in Sacchi et al.28 for the majority of the countries, Piroddi et al.24 for Italy, and Moutopoulos et al.30 for Greece. For Spanish and Italian trawlers, we complemented our trends with data from EVOMED32, a European project that assessed the evolution and technological improvement of fishing capacity for the major countries of the Mediterranean Sea for the early 1900–2010 period.
Fishing effort (kW*days−1) was calculated as the product of the number of fishing vessels, kW per vessel (inferred from their GT), and the number of days spent fishing. To account for improvements in technology (e.g., mobile phone, GPS, sonar, radio) that were not captured by kW as a measure of effort (Figure S5), a conservative technological “creep factor” of 1% as observed by Damalas et al.33 and EVOMED32 was applied from 1980 to 1995 while for the remaining periods (1950–1979 and 1996–2010) a 0.5% and ~ 1.9% (this value varied with gear type; see S6 Table) were respectively used following the work of Pauly and Palomares34.
Temporal dynamic modelling and model derived indicators
The dynamic module of the EwE software, Ecosim25,35, uses a set of differential equations to estimate biomass fluxes for each species and/or functional group of the ecosystem as follows:

where dBi/dt is the biomass growth rate of group (i) during the interval dt, gi is the net growth efficiency (production/consumption ratio), Ii is the immigration rate, Mi and Fi are natural and fishing mortality rates of group (i), ei is emigration rate and Bi the biomass25. Calculations of consumption rates (Qij) are based on the “foraging arena” theory36 where the biomass of prey i is divided between a vulnerable and a non-vulnerable fraction. This is represented as:

where vij and v’ij is the vulnerability and expresses the rate with which prey move between being vulnerable and not vulnerable, respectively, aij is the effective search rate for i (prey) by j (predator), Ti and Tj are the relative feeding time for prey and predator, Sij are the seasonal or long term forcing effects, Mij are the mediation forcing effects and Dj are the effects of handling time as a limit to consumption rate. One important feature in Ecosim is the use of a vulnerability term for each interaction between a predator and a prey. Low values of vulnerability (close to 1) indicate that prey production determines the predation mortality (phenomenon also known as ‘bottom-up’ control) and that the predator is close to carrying capacity, while high values of vulnerability (e.g., 100) indicate that predator biomass determines how much prey is consumed (top-down control) and that predators are far away from carrying capacity25. Mixed effect (vulnerability = 2) is set as the default value in Ecosim.
The Ecosim approach was used here to fit the model to observed time-series of data using the sum of squares (SS) ratio between predicted and observed data as a metric for assessing model performance37. We used survey biomasses and catches for those functional groups with available information to compare predicted and observed data (S1). In particular, biomass time series for sea turtles, pinnipeds, benthic invertebrates and deep sea fish were taken from scientific literature, whereas for demersal species (functional groups n° 12–14; 16–19 and 21 in Fig. 2), European anchovy, European pilchard and large pelagic fishes, we used scientific surveys (e.g., MEDITS trawl survey and MEDIAS acoustic survey) and stock assessments data (S2). Catch data was taken from the United Nation’s Food and Agriculture Organization (FAO) database (FishStat: http://data.fao.org/database) available from 1950 to 2010. These time series were complemented with data (available per country) from the Sea Around Us Project (www.searoundus.org) to assign species to fishing fleet.
When applying the fitting procedure, we noticed that the baseline fishing mortality (Ecopath baseline in 1950 s) for the most commercially important target species (European pilchard, anchovy and hake) was relatively low (between 0.02 and 0.05) compared to the reference levels reported in the literature38,39,40. This initially caused a very low reaction of these species to changes in historical fishing effort and primary productivity. To correct these estimates and reflect a more appropriate fishing mortality for these three species, we used the reconstruction of the catches of the Sea Around Us Project and, in particular, for each country of the Mediterranean Sea, we considered the proportion of catch of these species relative to the total catch and applied it respectively in each of our sub-areas.
To fit the temporal dynamic model accounting for data quality/reliability in available time series, we weighted the time series using a factor either of 0.5 or 1 (0 indicating that time series are not considered in the calculation of SS and 1 indicating that they are fully considered37). For all catch time series and for European pilchards and anchovies in the Ionian and Eastern Mediterranean Seas, we used a weight of 0.5, while the rest of the time series were assigned a weight of 1. This was done to consider questionable catch statistics reporting (as identified in previous research studies41,42,43,44), and to consider poor data availability for forage fish in the Ionian and Eastern Mediterranean Sea (i.e., long time series of European anchovies and pilchards were available only for the Aegean45 and the Strait of Sicily46,47). The choice of using these weights (0.5 and 1, respectively) puts less/more emphasis on selected species/functional groups of the ecosystem; still, since there are different methods to determine weighting factors27, further work should be developed to assess the outcome of the fit procedure using alternative weights.
Fishing effort (Figure S5-1) and primary production (PP) anomaly (Figure S5-2a) over time were used as main forcing time series to drive the model. The PP anomaly results from an Ecosim automated procedure that searches for time-series relative values of annual production (expressed as P/B ratio) of producer groups (in our case phytoplankton and seagrass). This routine considers that if primary production changes over time then the total amount of energy that enters in the ecosystem changes, causing a cascading-up effect that increases or decreases food availability through the ecosystem48. Once estimated by Ecosim, the predicted relative PP anomaly was tested against the relative PP (from phytoplankton) time-series data (Figure S5-2b) obtained from a biogeochemical model GETM-MedERGOM49 for the same time period using the Spearman’s rank-order correlation test (suitable for non-parametric data). Seagrass productivity, since it was not assessed in the biogeochemical model49, was excluded from the correlation test.
In addition, we re-run the Ecosim model using the relative PP time-series data from the biogeochemical model to compare and assess the model fit and results using the two different PP time series data (relative PP anomaly from Ecosim, and relative PP data from the biogeochemical model). As for fishing effort, since our reconstruction was done up to 2010, but the majority of our biomass time series were available until 2011, we decided to keep fishing efforts observed in 2010 constant until 2011.
The fitting procedure consisted of seven general steps (Table 1) following the same approach as described and applied by Mackinson50. This method uses the Akaike Information criterion (AIC)51,52:

where n is number of observations, minSS is the minimum sum of squares resulting from the comparison of predicted with observed datasets, and k is the number of parameters, to test statistical hypotheses related to changes in predator-prey dynamics (also called vulnerabilities: Vs); changes in primary production (PP anomaly, considering the number of PP spline points (sPP) for smoothing the time series); impact of fishing and possible combinations of the above-mentioned factors (Table 1). The AIC is a tool used for model selection that penalizes for fitting too many parameters, and which is used to choose the “best” model (the one yielding the lowest AIC) considering a good fit and the least number of estimated parameters to do so. In this study, we used the second-order Akaike Information Criterion (AICc) calculated as follow:

to account for small sample sizes (n of observations) in the dataset.
In our case, the fitting procedure was conducted five times: individually for the four sub-models (Western, Adriatic, Ionian and Eastern Mediterranean) as the majority of the functional groups are restricted to one sub-area only, and one extra time for the model representing the whole basin to fit highly migratory species (‘large pelagics’ and ‘sea turtles’ groups) that are allowed to move and feed in all four areas.
Once the temporal dynamic fitting procedure was completed, we used the “best” fitted models to calculate model-based indicators in each sub-area and for the whole Mediterranean Sea. To be able to compare these indicators with available ones from other regional seas, model-based indicators were selected from a list of indicators previously tested and assessed by international initiatives, mainly IndiSeas (“Indicators for the Seas”; www.indiseas.org; see e.g. refs 10,12). The list of selected indicators is presented in Table 2. Once estimated, we used the Spearman’s rank correlation to assess the significance and correlation between our suite of ecological indicators and time.
Addressing uncertainty
The Monte Carlo routine built into EwE25 was applied in Ecosim to assess sensitivity of Ecosim’s output to the basic Ecopath input parameters (B, P/B, Q/B, EE), drawing input parameters from a normal distribution centered on the base Ecopath value and using a defined coefficient of variation, in this case set to 0.137,53. Here, we run 1000 iterations, and the range of outputs (the 5th and 95th percentile) were plotted for both the fitted results (in our case time series of biomasses) and the model-based ecological indicators.
Results
Time series from the model fitting
The most statistically significant results in our model fitting exercise were obtained when trophic interactions, fishing and the primary productivity changes were included together in the model run (Step 7 in Table 3). Differences were found among the five areas with the “best” fitted models (lowest AICc) explaining from 50% to 69% of the variance of the data (Table 3). By looking at each area separately, the Ionian Sea sub-model was the one that showed the smallest improvement of prediction capabilities (thus the AICc estimates declined the least), while the Eastern followed by the Western Mediterranean were the areas with the highest improvement from the baseline AICc estimates. Both fishing and primary productivity drivers, when considered individually, were able to enhance the fit of all areas by ~16–~50% (when using the predicted PP anomaly) and by ~10–~37% when using fishing effort (steps 3 and 5 in Table 1).
The addition of trophic interactions to changes in PP anomaly alone (step 4 in Table 1) provided the second largest improvement for the Western, Ionian, and the whole Mediterranean Seas (AICc reduced further by ~10%). For the Adriatic Sea, this was obtained with the addition of trophic interactions to fishing effort (step 6 in Table 1). Also, different vulnerabilities were tested, and the largest enhancement was obtained using high vulnerabilities (step 7 in Table 3) for both the sub-models (maximum predator prey-interactions or Vs: #24) and the additional Mediterranean model as a whole (maximum predator prey-interactions or Vs: #2). Despite differences observed both at species/functional group and at sub-regional level, Ecosim results revealed that the Mediterranean Sea was mainly driven by bottom-up control interactions and low vulnerabilities were dominant (Western: 46%; Adriatic: 75%; Ionian: 50% and Eastern: 54%), followed by top-down controls interactions indicated by high vulnerabilities (Western: 42%; Adriatic: 21%; Ionian: 25% and Eastern: 33%). Mixed effect interactions represented a smaller proportion (vulnerabilities close to 2; Western: 13%; Adriatic: 4%; Ionian: 25% and Eastern: 13%) (S 14).
When we tested for correlation between the PP anomaly, resulting from the Ecosim fitting procedure, and the PP from the biogeochemical model, in all the areas these were positively and significantly correlated. Only the Adriatic Sea showed a significant negative correlation (Table 4 and Figure S7).
Using the “best” fitted models, Ecosim reproduced satisfactorily the biomasses trends for some of the functional groups with available survey data in all sub-areas (Figs 3 and 4). Overall, forage fishes (functional groups n. 8–9), demersal fishes (n. 12–14) and invertebrates (n. 18–19 and 21) showed a good fit in the different sub-models, while deep sea fish (n. 15) and benthos (n. 23) were the least well fitted (Figs 3, 4 and S7). These latter groups are the ones with the fewest data points. A satisfactory fit was also shown for sharks and rays/skates groups (n. 16–17), and, despite only few observed records, also for pinnipeds (n. 3) (Figs 3, 4 and S8).

Predicted biomass (t·km−2) is shown as solid black lines, while observed data is represented as black dots. Functional groups codes correspond to those given in Fig. 2. The predicted model (dashed red line) using modelled biogeochemical PP is also shown. Blue shadow represents the 95% percentile and 5% percentile obtained through the Monte Carlo routine. Drawings for species/functional groups 1–3, 5–6 and 8 are by Massimo Demma - http://www.massimodemma.it/-.

Predicted biomass (t·km−2) is shown as solid black lines, while observed data is represented as black dots. Functional groups codes correspond to those given in Fig. 2. The predicted model (dashed red line) using modelled biogeochemical PP is also shown. Blue shadow represents the 95% percentile and 5% percentile obtained through the Monte Carlo routine. Drawings for species/functional groups 1–3, 5–6 and 8 are by Massimo Demma - http://www.massimodemma.it/-.
In the Western Mediterranean, based on the biomass trends by area, the predicted time series suggested a decreasing pattern for the biomasses of several functional groups (Figs 3 and S8). European pilchard (n.8) showed a decline from the beginning of our study period (1950), which became more pronounced in the last years of the observed time series. A similar result was also observed for medium (n. 13) and small (n. 14) demersal fishes, and pinnipeds (n. 3), although the model was not able to capture the sharp decline of these marine mammals in the 1970 s. As for sharks (n. 16), rays/skates (n. 17) the model confirmed a decrease in trends until the end of the 1990 s and a slight increase in the 2000 s decade. For European anchovy (n. 9) and hake (n. 12), Ecosim had difficulties reflecting observed variations in their biomass, although suggesting a decreasing trend for both species. A poor fit was observed for benthos (n. 23) and deep sea fish (n. 15), where only few data points were available. A good reproduction of biomass time series was found for crustaceans (n. 21) and benthopelagic cephalopods (n. 18) where the model was able to follow the majority of the fluctuations in time (Fig. 3 and S8). When the model was run using the PP from the biogeochemical model as an alternative primary productivity driver, we observed similar pattern (red dashed line in Figs 3 and 4) as the ones obtained using the PP Ecosim anomaly, and for certain species/functional groups (n. 8 and n. 21 in Fig. 3) the fit improved.
As for the Western Mediterranean, also in the Adriatic Sea, Ecosim suggested a decline for some top predators, demersal and pelagic fish and for some invertebrates (Figs 3 and S8). In particular, the model was able to capture the steep decline of pinnipeds (n. 3) observed in the area since mid-1970s54 and a less marked decrease of medium (n. 13) and small (n. 14) demersal fish observed in mid 1990s55,56. Ecosim captured some of the pattern observed for European hake (n. 12), sharks (n. 16), rays/skates (n. 17) suggesting a decline of the groups until the end of the 1990 s, followed by a slight increase or by fluctuations (in the case of European hake) in the last years of the studied period. An overall satisfactory match between predicted and available data was found for benthopelagic cephalopods (n. 18) where a decrease was captured since the beginning of the survey period, and for benthic cephalopods (n. 19) and crustacean (n. 21) where the model followed some of the fluctuation of the groups and a slight increase at the end of 2000 s. Again, the model did not represent the trends well for deep sea fish (n. 15; Figure S8). Regarding forage fish (n. 8–9), when we run the model using PP anomaly as driver, Ecosim was not able to reflect the decreasing biomass trend observed in European anchovies, while it was able to pick up a general decline for European pilchards. However, it was when we applied the PP from the biogeochemical model in the model run that Ecosim was able to follow the steep decline observed in European anchovies in mid 1970 s and improve also slightly the decline of European pilchard. For the other species/functional groups, different trends were observed using the two different PPs particularly in the decades before the beginning of our time series of observations (Figs 3 and S8).
The Ionian Sea resulted to be the area with less biomass changes during the years with available survey data (Figs 4 and S8). Except for pinnipeds (n. 3), where the model was able to pick up the decline since the late 1970, despite the presence of only few data points, all the other groups didn’t show any directional variation in time resulting mainly in a series of fluctuations. However, by looking at the overall time period (1950–2011), the model suggested a small increase in biomass since the beginning of 1990 s for small demersal fish and crustaceans. The model partly underestimated and was not able to capture the biomass trends for European pilchards and medium demersals (respectively n. 8 and n. 13; Figs 4 and S8), and it did not represent well the trend for benthos (n. 23; Figure S8). The use of PP from the biogeochemical model improved slightly the fit for crustaceans (n. 21), sharks (n. 16) and benthopelagic cephalopods (n. 18) while maintaining the same pattern observed with the PP anomaly.
In the Eastern Mediterranean, different trends among species/functional groups were detected (Figs 4 and S8). Ecosim represented relatively well the biomass declines of European pilchards (n. 8) and anchovies (n. 9) since the 1990 s, despite underestimating the high peaks observed at the beginning of this decade. The model was able to capture the biomass trends for European hake (n. 12), small demersals (n. 14), sharks (n. 16), rays/skates (n. 17), benthic cephalopods (n. 19) and crustaceans (n. 21). All these groups showed similar patterns with signs of decrease in the 1990 s and fluctuations afterwards. An underestimation of biomass by the model was predicted for medium demersal fish (n. 13), deep sea fish (n. 15), benthopelagic cephalopods (n. 18), and benthos (n. 23) where the model was not able to reproduce observed trends and fluctuations (Figure S8). A good fit, even though for only few data points, was found for pinnipeds (n. 3) where the model was able to represent the fluctuation of these marine mammals over time (Fig. 4). The predicted trends obtained using PP from the biogeochemical model were similar to the ones found using the PP anomaly and for European hake (n. 12), small demersals (n. 14), sharks (n. 16), benthic cephalopods (n. 19) and crustaceans (n. 21) the fit slightly improved. Ecosim was able to represent the decrease in biomass of large pelagic fish (n. 6) particularly since the 80 s, while it failed to capture the fluctuation observed at the end of the 2000 s in the whole Mediterranean model for the two highly migratory species for which we had survey data: large pelagics and sea turtles. In the case of the sea turtles (n. 5), the model approximated the general increasing biomass trend, but it failed to reproduce its fluctuations over time (Fig. 5). We observed similar results with the PP from the biogeochemical model as a driver.

Predicted biomass (t·km−2) is shown as solid black lines, while observed data is represented as black dots. Functional groups codes correspond to those given in Fig. 2. The predicted model (dashed red line) using modelled biogeochemical PP is also shown. Blue shadow represents the 95% percentile and 5% percentile obtained through the Monte Carlo routine. Drawings for species/functional groups 1–3, 5–6 and 8 are by Massimo Demma - http://www.massimodemma.it/-.
The time series of catch trends estimated for the five areas, when compared with independent data, showed a general satisfactory match (Figs 6 and S9): the sub-models overestimated or underestimated some fractions of the time series trends, but overall, they were able to capture long-term trends similar to those observed (Fig. 6). In the Western Mediterranean, an increase (up to the end of the 1990 s) and posterior decrease in catches were predicted for the majority of the groups with the exception of other small pelagic fish (n. 10), large demersal (n. 11), and benthic cephalopods (n. 19) that continued to increase even afterwards. Non-significant trend was simulated for rays/skates (n. 17), while the model was not able to reflect the trend observed for benthopelagic cephalopods (n. 18). Regarding large pelagic fishes (n. 6), catches predicted for the whole Mediterranean were similar to those observed until the 1980 s, but the predicted catches did not reflect the increase observed in the last two decades (Fig. 6).

Predicted (solid lines) versus observed (dots) catches (t·km−2·year−1) for main commercially important functional groups of the Western Mediterranean (a) and Adriatic (b) ecosystems (1950–2011). Predictions obtained with the Mediterranean Sea model as whole for large pelagic catches are included in the Western Mediterranean plot (a. #6). Results for the Ionian and Aegean catches are shown in Figure S9. Drawings for species/functional groups 1–3, 5–6 and 8 are by Massimo Demma - http://www.massimodemma.it/-.
In the Adriatic, as for the Western Mediterranean, the model simulated the decrease in catches observed in the beginning of the 1990 s for the majority of the functional groups while it did not succeed to pick up the sharp decline of European anchovies (n. 9) in mid 1970 s, and of European hake (n. 12) and sharks (n. 16) in the 1990 s (Fig. 6).
In the Ionian Sea, the model predicted the increase in catches until the end of 2000 s for the majority of the functional groups. For European hake (n. 12), medium demersals (n. 13), sharks (n. 16), rays/skates (n. 17) and benthic cephalopods (n. 19), though, such increase turned into a decrease approximately around the 1990 s (Figure S9). In the Eastern Mediterranean Sea, predicted results indicated the increase in catches for the majority of the functional groups until the 1990 s and the decline afterwards and they also captured the continuous increase for other small pelagic fishes (n. 10) and benthopelagic cephalopods (n. 18). On the other hand, simulated results did not match the sharp decline of sharks (n. 16) observed since the 1980 s in the region (Figure S9).
Temporal model-based ecological indicators
Trends in ecological indicators calculated from Ecosim temporal outputs showed different patterns if we looked at each sub-regional sea individually or at the Mediterranean ecosystem as a whole. For example, considering the entire Mediterranean Sea, a clear decreasing trend was observed in community biomass indicators like the forage fish biomass (on average −34%) and, to less extent, for demersal fish (−14%), the Kempton’s biodiversity index (−18%) and in all the trophic level indicators considered (TLco, TL ≥ 3.25 and TL Catch) (−5%) (Fig. 7). On the contrary, an increase was predicted for invertebrate biomass (+23%) while no clear trend was visible for sharks and rays/skates. Total catch was the only indicator that clearly increased in time (until 1990 s) (+189%) and that gradually decreased afterwards. These patterns were also reflected through the Spearman correlation test (Fig. 8).

Forage fish biomass (t·km−2); 2. Demersal fish biomass (t·km−2); 3. Invertebrates biomass (t·km−2); 4. Sharks/rays and skate biomass (t·km−2); 5. Kempton Q: Kempton’s index of biodiversity; 6. TLco: Mean trophic level of the community; 7. TLco ≥ 3.25: Mean trophic levels of groups having trophic level ≥ 3.25 (excluding marine mammals, sea turtles and seabirds); 8. Tot Catch: Total catch (t· km−2 ·year-1); 9 TL Catch: Mean trophic level of the catches) estimated from the Ecosim results for the period 1950–2011 for the Catch Mediterranean Sea. Blue shadow represents the 95% percentile and 5% percentile obtained through the Monte Carlo routine.

Positive correlations are displayed in blue and negative correlations in red color. Color intensity and the size of the eclipses are proportional to the correlation coefficients. At the right side of the graph, the legend color shows the correlation coefficients and the corresponding colors. The color follows a gradient according to the strength of the correlation. The width of the ellipses is related to correlation strengths with more diffused ellipses representing lower correlation strengths. When the indicators are non-significant (>0.05) are represented with an X symbol.
Considering sub-regional seas (Figures S10–S13), we observed a clear decline of forage fish (−41%), demersal fish (−49%) and sharks/rays-skates (−60%) biomasses in the Western and Adriatic Seas, a fluctuation of these groups in the Ionian Sea while in the Eastern Mediterranean they respectively decreased, increased and fluctuated. Invertebrate biomass slightly decreased in the Adriatic Sea (−13%); fluctuated in the Western and Ionian Seas; and increased in the Eastern Mediterranean (+53%). The Kempton biodiversity index decreased in the Western (−49%) and in the Ionian Sea (−41%), it showed a slight increase in the Adriatic (+9%) while no clear trend was visible in the Eastern Mediterranean. Total catch increased in all the areas (+189%) until the beginning of 1990 s but in the Western and Ionian Seas started to fluctuate afterwards while in the Eastern and Adriatic Sea it gradually declined. As for the different trophic level indicators assessed, the mean TL of the community slightly increased in the Western Mediterranean (+10%) and decreased in the other sub-regions (−5%), while the mean TL ≥3.25 and mean TL catches decreased in all the seas (−4%) except in the Eastern Mediterranean where they respectively fluctuated with no clear trend and slightly increased (Figures S10–S13).
When we tested the significance and correlation of our suite of temporal ecological indicators, we noticed that in the Western and the Adriatic Seas the majority of the time series were negatively correlated with high significance (respectively 6 and 7 out of 9 indicators; Fig. 8). On the contrary, in the Ionian Sea and Eastern Mediterranean Sea, the community indicators (except for forage fishes in the Ionian that showed a weak negative correlation) were highly significant and positively correlated (Fig. 8). Also, we observed no significant and weakly correlated trends for mean TL ≥3.25 and Kempton biodiversity index in the Eastern Mediterranean Sea.
Discussion
Historical ecosystem drivers of the Mediterranean Sea ecosystem
Modelling results explained from 50% to 69% of the variability of historical time series data utilised in the present study. Both fishing pressure and PP anomaly played an important role in improving the model fit. In addition, our results indicated that the PP anomaly, representing the temporal variation of the primary productivity of the system, was the strongest driver upon the Mediterranean Sea ecosystem. This supports the results obtained from other studies49,57 that have shown how the Mediterranean Sea is driven by bottom-up processes where nutrient availability controls the biological characteristics of the region. The use of relative PP trends from a regional biogeochemical model helped validating our predicted PP anomaly trend and improved the temporal dynamics of selected species in the ecosystem (particularly for forage fish: European pilchards and anchovies). This was clearly visible, for example, in the Adriatic Sea where PP anomaly unsuccessfully reproduced the trends of European anchovies while PP trends from the biogeochemical model was able to better capture the dynamics. Since there are no official long-trend (from the 1950 s) records of primary production in the region (Macias et al. 2014), using a modelling framework that combines hydro-dynamic biogeochemical models with ecosystem models (such as EwE), becomes critically important, particularly in complex areas like the Adriatic Sea that show strong physical and biological oceanographic characteristics [decreasing gradient of trophic conditions from north to south58 ] and is also subjected to strong anthropogenic pressures (e.g., fishing)59,60. Currently there is a growing interest in these ensemble modelling frameworks5,21 in order to improve scientific capability to predict future ecosystem changes, and provide guidance for the setting of targets and implementation of management measures8,61. Accordingly to Macias et al.62, which explored potential changes in primary productivity in the Mediterranean Sea under different emission scenarios of future climate change, the Mediterranean basin will become warmer and more saline with consequences on the productivity of the region. The Western basin is expected to become more oligotrophic, associated to a surface density decrease influenced by the Atlantic waters, while the Eastern basin, on the contrary, is predicted to become more eutrophic due to a surface water density increase caused by increasing evaporation rate. Since these results are expected to alter the structural and functional processes shaping the Mediterranean food web, modelling tools like the one developed here are necessary in predicting the effects of climate variability and change on the future food web configurations.
Our study also highlights that fishing was an important driver affecting the dynamics of fish populations and invertebrates of the Mediterranean Sea ecosystem. This is in line with previous studies that documented the increasing impact of fishing in the Mediterranean Sea and the overexploitation of its marine resources39,40,56,63. Simulations, in fact, were able to reflect the impact of increased fishing effort in the basin starting, in all the four sub-areas, since the ‘50 s. Nominal fishing effort showed decreasing trends only after 2000; the only exception was found in the Eastern Mediterranean Sea where fishing effort showed a fluctuating trend in the 2000 s decade.
Historical trends of biomass, catch and ecological indicators
We provide here new insights on temporal dynamics of major marine species/functional groups of the Mediterranean Sea ecosystem. In general, both biomass trends and ecological indicators revealed that the combined effect of excessive fishing pressure and changes in the primary productivity have altered the Mediterranean marine ecosystem over time, especially reducing the proportions of top predators and larger fish (e.g., pinnipeds, large pelagic fish) and increasing the abundance of groups at lower trophic levels (e.g., invertebrates). This was already observed from west to east in other studies, for example, in the Catalan64, Adriatic55,59 and Ionian65,66 Seas. Our results also show that forage fish species were observed to decrease, at a different time scale, in the majority of the studied Mediterranean sub-areas; with the only exception in the Ionian Sea where no clear trends were observed. These forage fish (mainly European pilchard and anchovy) constitute the bulk of fish catches in the Mediterranean Sea, accounting for almost 40% of total landings67 and they are of high commercial interest. Therefore, an increase of fishing mortality, together with changes in productivity, have affected these stocks throughout the Mediterranean Sea. As for the Ionian Sea, the results obtained here should be taken with caution. Our fitting analysis for the majority of the species/functional group in this area didn’t show any clear trend besides fluctuations over time. These results for the Ionian Sea disagree with several studies that have shown decreasing trend in the abundance of many commercial and uncommercial species in the area47,65,68. Poor model performance could be related to poor quality of the available data used in our study (e.g., for forage fish species, long time series were available only from Sicily), or to the fact that important additional factors were missing from our modelling analysis (e.g., changes in oceanographic and physical characteristics, quality of prey availability, etc.) that could be affecting Ionian Sea populations. This will need further research.
Trends in demersal fish stocks also show signs of decrease, both at regional and sub-regional scale (specifically in the Western and Adriatic Sea), while sharks (which in our model were mainly represented by demersal species, see S1) and rays/skates seemed to have declined in the Western and Adriatic regions, but not in the whole Mediterranean Sea. Part of these results are in line with historical69 and recent studies39,40,70, which pointed to increased fishing pressure and lack in gear selectivity as the reason why 85% of the assessed demersal stocks (including demersal sharks, rays and skates) are currently overexploited.
Overall, our estimates of species declines are conservative in comparison with results from fisheries stock assessments available for the Mediterranean Sea39,40,71. The reason behind is likely related to the lack of data particularly in the 1950s–1970s period, which probably contributed to lower the reduction captured by the model in term of abundance of certain functional groups. In addition, our model was built with functional groups, which in some cases aggregated a vast number of species including commercial and non-commercial ones, and/or considered the North and South Mediterranean Seas together. This is probably masking the largest declines of highly commercial species in the Northwestern and Central Mediterranean Sea areas that we observe when looking at these species individually.
A clear sign of change in the structure of the Mediterranean Sea ecosystem is visible from results of the mean trophic level of the community, mean TL ≥ 3.25 and Q diversity index (which includes those species or groups with TL ≥ 3) showing a decline since 1950 s and reflecting the decline of large predators/fish stocks and increase of lower trophic level organisms. These results are consistent with previous ecosystem assessments17,63, although it is important to bear in mind that these results were assessed considering both fitted and non-fitted groups. Caution should be taken when interpreting the results. Differences in ecological indicators (e.g., community and TL based indicators) were found among the different sub-regions analyzed and, because of lack of studies at sub-regional scale to validate these results, we would like to stress the need to further develop this work.
Regarding catches, the fitting procedure enabled us to detect issues related to landings data at the beginning of our survey period. Low fishing mortalities were observed in the ‘50 s, in each sub-area, for three very important commercial species (European pilchard, anchovy and hake). Mortality rates for these species were between 5 and 10 times lower than the average reference values reported for these fish stocks in the Mediterranean Sea38,39,56. This supports the hypothesis, already highlighted by several studies30,41,72,73, of poor quality of fisheries statistics, particularly in historical times (1950–1970). Part of this problem could be related to the different way fisheries data were collected and aggregated by the different countries and regional institutions74. Poor data quality and availability could also be explained by the presence of illegal, unreported and/or unregulated (IUU) fishing activities occurring in the region41,42,43,44 especially with regard to illegal nets and mesh sizes, the landing and marketing of undersized fish, and compliance with restrictions on fishing season and areas39. This highlights the need to utilize better catch data in modelling exercises in the Mediterranean Sea in order to account for more realistic fishing mortality estimates and trends, and guide proper management decisions. Recent catch reconstruction efforts, which aim at considering all types of fisheries removals (from reported and unreported landings to recreational landings and discards), have been constructed and are now available (www.seaaroundus.org) for the different countries of the Mediterranean Sea75. In this study, the catch reconstruction has been partly used to fill up gaps encountered in the FAO catch database (e.g., contribution of specific species/functional groups to fishing fleets and the proportion of the reconstructed catches of European pilchard/anchovy/hake relative to the total catch). Unfortunately, though, at the time this modelling exercise was completed, the reconstruction of total catches for Mediterranean countries was still being developed. Therefore, a necessary further step of this study should be the integration of such catch reconstruction in the input modelling parameters to compare results with the current implementation.
Despite limitations, our model was able to reflect the temporal trends of fisheries across the Mediterranean Sea, with a general increase in the total catch and a decline in the mean TL catch. Such patterns could reflect that catch composition, with a highly diversified targeted species, continues to change in time as a result of fisheries expansion to further and deeper fishing grounds41,42. A different picture emerges when looking at total catches per sub-regional area, where clear signs of decrease are noticeable mainly in the Adriatic and the Eastern Mediterranean Sea - and for the last simulation years also in the Western Mediterranean and Ionian Seas. These results are consistent with previous studies that have pointed out excessive fishing mortality and food web degradation caused by fishing in the Eastern and Adriatic fisheries55,56,76. On the other hand, the more stable catches observed in the Western Mediterranean and Ionian Sea could be the result of exploiting new species, as observed for the Mediterranean as a whole42.
The trophic level of the catches for the whole Mediterranean Sea and as well for the majority of the sub-areas (Western, Adriatic and Ionian Seas) presented a clear ‘fishing down’ effect77 that occurs when top predators and large sized fish are removed from the ecosystem and gradually replaced by lower trophic level organisms. Similar trends had been observed in the Mediterranean Sea, both at regional77, sub-regional56 and more local scale10,78. The only exception was found in the Eastern Mediterranean Sea where, contrary to the rest of areas, a situation of ‘fishing up’ was found. Accordingly to Stergiou and Tsikliras79, though, this might be a ‘false fishing up effect’ occurring when small pelagic fishes and invertebrates, with a low trophic level, and larger-size predators fish are both intensely fished and/or depleted.
Management and conservation implications
The Mediterranean has been exploited for centuries, suffering the impacts of continuous and multiple anthropogenic pressures19,80. Because of increasing signs of deteriorations and degradations at species-, community- and ecosystem levels19,39,80, evidenced as well by this study, the basin is now of particular concern, and is a clear candidate for management actions to halt further decline and increase the sustainable use of marine resources74. Hindcasting analyses, as performed in this study, allow assessing historical changes in the ecosystem and in its marine resources, and are necessary pieces of the tool kit needed to support management and conservation processes.
Yet, to move towards better information to support regional policy and conservation plans, several additional steps should be developed in the near future.
First, spatial-temporal analyses able to identify spatial patterns that can directly assist spatial management actions (e.g., by prioritizing specific areas of concern), and facilitate the communication between scientists and policy makers, environmental managers, conservationists and the general public80 are needed to contribute to the recent Maritime Spatial Planning Directive (MSPD) of the European Commission81. A first attempt has been made in a recent study by Liquete et al.82, which assessed the delivery of five marine ecosystem services for the whole Mediterranean basin using several modelling approaches, both in time and space. However, as pointed out by these authors, more work is needed to be able to support management decisions.
Second, the integration of additional human stressors (e.g., aquaculture, invasive species, water warming, acidification, pollution, habitat degradation) as driving forces of species dynamics is needed to increase the reliability of this modelling exercise since marine ecosystems are impacted by simultaneous cumulative threats19,80. Currently the recent MSPD, which include the EU’s Blue Growth Strategy83 that supports sustainable growth in emergent marine sectors (e.g., aquaculture, coastal tourism, marine energies), is expected to impose further pressures on the Mediterranean Sea19,84.
Third, the development of forecast scenarios, including different future management actions, is crucial for the implementation of management plans. Future scenarios should follow the Intergovernmental Panel on Climate Change (IPCC) projections on climate-induced changes in sea surface temperature. They should also consider the relevant elements of the Common Fisheries Policy (CFP) on commercially important stocks to exploit them at Maximum Sustainable Yield (MSY) levels and the reduction of fishing effort needed to develop effective and appropriate policy and conservation plans in the region39,56.
In addition, this work emphasizes the need to use end-to-end modelling approaches as strategic tools for an explicit support to the decision-making process. Several end-to-end (E2E) models have now been developed22,23,61 to capture the feedbacks and multi-scale interactions that drive ecosystems dynamics with the goal of guiding responsible resource management decisions. As pointed out by Fulton85, there is no “one best E2E model”, on the contrary, a suite of tools should be considered given that such approaches are normally used to address questions at a scale where there is still a lot of uncertainty about how systems function. Our study advanced in this direction using results of complementary models together.
Model assumptions and limitations
Modelling the Mediterranean Sea ecosystem is a challenging task, not only because of the complex dynamics that characterize this Large Marine Ecosystem (e.g., differences in environmental and biological features), but also because of the difficulties of gathering and integrating regional data24. Several gaps have been already identified and described in a previous work of Piroddi et al.24 which identified the lack of trophic information with a temporal dimension, the limited information on biomass of different species (especially of those non-commercially important species and deep-sea organisms), and the lack of reliable catch data and fishing effort. The absence of time series of seagrass productivity at regional scale is another constraint. Assessing the role and contribution of seagrass productivity on the overall marine primary productivity is critically important for understanding how changes in seagrass growth rates might impact the different compartments of an ecosystem86. In addition, the lack of historical data series (particularly between 1950 s and 1970 s), and the problems with data accessibility limit the potential resolution of modelling studies and the development of robust EBM approaches24,74,87. Therefore, more efforts should be dedicated to improve data collection and quality, and to make data better accessible for the region.
This study includes the best available regional data (see Supplementary Information) and highlights, when necessary, gaps and difficulties encountered in the modelling process (see below). To account for the uncertainty around the model parameters, we applied a Monte Carlo routine to evaluate model outputs sensitivity (in our case for biomasses and model derived indicators) to data uncertainty. Considering data uncertainty in model development is critical if the purpose of modelling is to inform policy/management processes6,27. Still, the majority of available modelling tools lack an approach to take uncertainty of modelled data (both input and output) into account8,88.
Although some time series were not always well replicated and uncertainty analyses can be improved as higher quality data becomes available, our modelling exercise reproduced several surveyed datasets in a satisfactory way and, as such, it is to date the best available representation of historical trends for the Mediterranean Sea as a whole, and a first step towards the integrated and historical understanding of this complex ecosystem.
Conclusions
With anthropogenic pressures rapidly expanding in the Mediterranean Sea, there is a serious risk that these may push the system beyond the “point of no-return”, with consequence for marine biodiversity and the economies that depend on it, seriously constraining the ecosystem service options available to future generations. Ecosystem modelling tools can support the analysis and identification of the potential suitable options for ensuring the coexistence of sustainable human activities and the protection of healthy marine ecosystems. Temporal hind-cast analysis has enabled us to assess changes in the historical dynamics of species/functional groups inhabiting this system, quantifying the role and impact of changes in primary productivity and fishing pressure. Since climate variability and change in combination with fishing pressure is expected to intensify in the region, modelling approaches like the one presented here are necessary in predicting the effect of changes of the above-mentioned pressures on the marine food web. This constitutes an important first step further to advance in the regional assessment of the Mediterranean Sea ecosystem to inform conservation plans and management actions.
Additional Information
How to cite this article: Piroddi, C. et al. Historical changes of the Mediterranean Sea ecosystem: modelling the role and impact of primary productivity and fisheries changes over time. Sci. Rep. 7, 44491; doi: 10.1038/srep44491 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Halpern, B. S. et al. A global map of human impact on marine ecosystems. Science 319, 948–952 (2008).
Côté, I. M., Darling, E. S. & Brown, C. J. Interactions among ecosystem stressors and their importance in conservation Proc. R. Soc. B. 20152592 (2016).
Mora, C. et al. Biotic and human vulnerability to projected changes in ocean biogeochemistry over the 21st century. PLoS Biol 11, e1001682 (2013).
Halpern, B. S. et al. Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nature communications 6 (2015).
Travers, M. et al. Two-way coupling versus one-way forcing of plankton and fish models to predict ecosystem changes in the Benguela. Ecological modelling 220, 3089–3099 (2009).
Collie, J. S. et al. Ecosystem models for fisheries management: finding the sweet spot. Fish and Fisheries(2014).
Christensen, V. & Walters, C. J. In Ecosystem Approaches to Fisheries: A Global Perspective(eds V. Christensen & J. L. Maclean ) 189–205 (Cambridge University Press, 2011).
Piroddi, C. et al. Using ecosystem models to assess biodiversity indicators in support of the EU Marine strategy framework directive. Ecological Indicators 58, 175–191 (2015).
Levin, P. S., Fogarty, M. J., Murawski, S. A. & Fluharty, D. Integrated ecosystem assessments: developing the scientific basis for ecosystem-based management of the ocean. PLoS Biology 7, 23–28 (2009).
Shannon, L. et al. Trophic level-based indicators to track fishing impacts across marine ecosystems. Marine Ecology Progress Series 512, 115–140 (2014).
Cury, P. M. et al. Ecosystem oceanography for global change in fisheries. Trends in Ecology & Evolution 23, 338–346 (2008).
Coll, M. et al. Ecological indicators to capture the effects of fishing on biodiversity and conservation status of marine ecosystems. Ecological Indicators 60, 947–962 (2016).
Ehler, C. N. Indicators to measure governance performance in integrated coastal management. Ocean & Coastal Management 46, 335–345 (2003).
Rice, J. C. & Rochet, M.-J. A framework for selecting a suite of indicators for fisheries management. ICES Journal of Marine Science: Journal du Conseil 62, 516–527 (2005).
Halpern, B. S. et al. An index to assess the health and benefits of the global ocean. Nature 488, 615–620 (2012).
Shin, Y.-J. et al. Can simple be useful and reliable? Using ecological indicators to represent and compare the states of marine ecosystems. ICES Journal of Marine Science: Journal du Conseil 67, 717–731 (2010).
Coll, M. et al. The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS one 5, e11842 (2010).
Bianchi, C. & Morri, C. Marine biodiversity of the Mediterranean Sea: situation, problems and prospects for future research. Marine Pollution Bulletin 40, 367–376 (2000).
Coll, M. et al. The Mediterranean Sea under siege: spatial overlap between marine biodiversity, cumulative threats and marine reserves. Global Ecology and Biogeography 21, 465–480 (2012).
Costello, M. J. et al. A census of marine biodiversity knowledge, resources, and future challenges. PloS one 5, e12110 (2010).
Rose, K. A. et al. End-to-end models for the analysis of marine ecosystems: challenges, issues, and next steps. Marine and Coastal Fisheries 2, 115–130 (2010).
Travers-Trolet, M., Shin, Y.-J., Shannon, L. J., Moloney, C. L. & Field, J. G. Combined Fishing and Climate Forcing in the Southern Benguela Upwelling Ecosystem: An End-to-End Modelling Approach Reveals Dampened Effects. PloS one 9, e94286 (2014).
Niiranen, S. et al. Combined effects of global climate change and regional ecosystem drivers on an exploited marine food web. Global change biology 19, 3327–3342 (2013).
Piroddi, C., Coll, M., Steenbeek, J., Moy, D. M. & Christensen, V. Modelling the Mediterranean marine ecosystem as a whole: addressing the challenge of complexity. Mar Ecol Prog Ser 533, 47–65 (2015).
Christensen, V. & Walters, C. J. Ecopath with Ecosim: methods, capabilities and limitations. Ecological modelling 172, 109–139 (2004).
Link, J. S. Adding rigor to ecological network models by evaluating a set of pre-balance diagnostics: a plea for PREBAL. Ecological Modelling 221, 1580–1591 (2010).
Heymans, J. J. et al. Best practice in Ecopath with Ecosim food-web models for ecosystem-based management. Ecological Modelling(2016).
Sacchi, J. Analysis of economic activities in the Mediterranean: Fishery and aquaculture sectors. 84 (2011).
Stergiou, K. I., Moutopoulos, D. K., Tsikliras, A. C. & Papaconstantinou, C. In State of Hellenic Fisheries(eds C. Papaconstantinou, A. Zenetos, V. Vassilopoulou & G. Tserpes ) 132–140 (HCMR, 2007).
Moutopoulos, D. K. & Koutsikopoulos, C. Fishing strange data in national fisheries statistics of Greece. Marine Policy 48, 114–122 (2014).
Greer, K. Considering the ‘effort factor’in fisheries: a methodology for reconstructing global fishing effort and carbon dioxide emissions. 1950–2010 (2014).
EVOMED. The 20th Century evolution of Mediterranean exploited demersal resources under increasing fishing disturbance and environmental change. (2011).
Damalas, D. et al. “Once upon a Time in the Mediterranean” long term trends of mediterranean fisheries resources based on fishers’ traditional ecological knowledge. PloS one 10, e0119330 (2015).
Pauly, D. & Palomares, M. An empirical equation to predict annual increases in fishing efficiency. Fisheries Centre University of British Columbia Working Paper Series 07 (2010).
Walters, C., Christensen, V. & Pauly, D. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Reviews in fish biology and fisheries 7, 139–172 (1997).
Ahrens, R. N., Walters, C. J. & Christensen, V. Foraging arena theory. Fish and Fisheries 13, 41–59 (2012).
Christensen, V., Walters, C., Pauly, D. & Forrest, R. Ecopath with Ecosim 6: A user’s guide. Fisheries Centre, University of British Columbia, Vancouver. (2008).
Patterson, K. Fisheries for small pelagic species: an empirical approach to management targets. Reviews in Fish Biology and Fisheries 2, 321–338 (1992).
Colloca, F. et al. Rebuilding Mediterranean fisheries: a new paradigm for ecological sustainability. Fish and fisheries 14, 89–109 (2013).
Vasilakopoulos, P., Maravelias, C. D. & Tserpes, G. The alarming decline of Mediterranean fish stocks. Current Biology 24, 1643–1648 (2014).
Pauly, D., Ulman, A., Piroddi, C., Bultel, E. & Coll, M. ‘Reported’ versus ‘likely’ fisheries catches of four Mediterranean countries. Scientia Marina 78, 11–17 (2014).
Coll, M. et al. Closer to reality: Reconstructing total removals in mixed fisheries from Southern Europe. Fisheries Research 154, 179–194 (2014).
Ulman, A. et al. From bonito to anchovy: a reconstruction of Turkey’s marine fisheries catches (1950–2010). Mediterranean Marine Science(2013).
Piroddi, C. et al. Reconstruction of Italy’s marine fisheries removals and fishing capacity, 1950–2010. Fisheries Research 172, 137–147 (2015).
Jardim, E. et al. Stock assessment of Hellenic small pelagic stocks. EUR 27484. Luxembourg (Luxembourg): Publications Office of the European Union; 2015. JRC97817. (2015).
Fiorentino, F. et al. A comparison between acoustic and bottom trawl estimates to reconstruct the biomass trends of sardine and anchovy in the Strait of Sicily (Central Mediterranean). Fisheries Research 147, 290–295 (2013).
Patti, B. et al. Interannual fluctuations in acoustic biomass estimates and in landings of small pelagic fish populations in relation to hydrology in the Strait of Sicily. Chemistry and Ecology 20, 365–375 (2004).
Preikshot, D. B. The influence of geographical scale, climate and trophic dynamics upon North Pacific Oceanic ecosystem models, University of British Columbia (2007).
Macias, D., Garcia‐Gorriz, E., Piroddi, C. & Stips, A. Biogeochemical control of marine productivity in the Mediterranean Sea during the last 50 years. Global Biogeochemical Cycles 28, 897–907 (2014).
Mackinson, S. Combined analyses reveal environmentally driven changes in the North Sea ecosystem and raise questions regarding what makes an ecosystem model’s performance credible? Canadian Journal of Fisheries and Aquatic Sciences 71, 31–46 (2014).
Akaike, H. A new look at the statistical model identification. Automatic Control, IEEE Transactions on 19, 716–723 (1974).
Burnham, K. P. & Anderson, D. R. Model selection and multimodel inference: a practical information-theoretic approach. (Springer Science & Business Media, 2003).
Coll, M. & Steenbeek, J. New software plug-in to calculate biodiversity and conservation-based indicators from EwE food web models. In Ecopath 30 Years Conference Proceedings: Extended Abstracts. (ed. J. Steenbeek, C. Piroddi, M. Coll, J.J. Heymans, S., Villasante, V. Christensen) 67–68 (Fisheries Centre, 2014).
Sergeant, D., Ronald, K., Boulva, J. & Berkes, F. The recent status of Monachus monachus, the Mediterranean monk seal. Biological Conservation 14, 259–287 (1978).
Lotze, H. K., Coll, M. & Dunne, J. A. Historical changes in marine resources, food-web structure and ecosystem functioning in the Adriatic Sea, Mediterranean. Ecosystems 14, 198–222 (2011).
Tsikliras, A. C., Dinouli, A., Tsiros, V.-Z. & Tsalkou, E. The Mediterranean and Black Sea fisheries at risk from overexploitation. PloS one 10, e0121188 (2015).
Coll, M., Piroddi, C., Christensen, V., Palomera, I. & Arneri, E. Main drivers of marine resources and food-web changes in the Mediterranean Sea. In Ecopath 25 Years Conference Proceedings: Extended Abstracts. (ed M.L.D. Palomares, L. Morissette, A. Cisneros-Montemayor, D. Varkey, M. Coll, C. Piroddi) 148–149 (Fisheries Centre, 2009).
Polimene, L., Pinardi, N., Zavatarelli, M. & Colella, S. The Adriatic Sea ecosystem seasonal cycle: Validation of a three‐dimensional numerical model. Journal of Geophysical Research: Oceans 111 (2006).
Coll, M., Santojanni, A., Palomera, I. & Arneri, E. Food-web changes in the Adriatic Sea over the last three decades. Marine Ecology Progress Series 381, 17–37 (2009).
Steenbeek, J. et al. Bridging the gap between ecosystem modeling tools and geographic information systems: Driving a food web model with external spatial–temporal data. Ecological Modelling 263, 139–151 (2013).
Kaplan, I. C., Horne, P. J. & Levin, P. S. Screening California Current fishery management scenarios using the Atlantis end-to-end ecosystem model. Progress in Oceanography 102, 5–18 (2012).
Macias, D. M., Garcia-Gorriz, E. & Stips, A. Productivity changes in the Mediterranean Sea for the twenty-first century in response to changes in the regional atmospheric forcing. Frontiers in Marine Science 2, 79 (2015).
Lotze, H. K. et al. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312, 1806–1809 (2006).
Coll, M., Palomera, I., Tudela, S. & Sardà, F. Trophic flows, ecosystem structure and fishing impacts in the South Catalan Sea, Northwestern Mediterranean. Journal of Marine Systems 59, 63–96 (2006).
Piroddi, C., Bearzi, G. & Christensen, V. Effects of local fisheries and ocean productivity on the northeastern Ionian Sea ecosystem. Ecological modelling 221, 1526–1544 (2010).
Piroddi, C., Bearzi, G., Gonzalvo, J. & Christensen, V. From common to rare: the case of the Mediterranean common dolphin. Biological Conservation 144, 2490–2498 (2011).
FAO. Capture Production 1950–2010. http://www.fao.org/fishery/statistics/software/fishstat/en (2012).
Machias, A., Stergiou, K., Somarakis, S., Karpouzi, V. & Kapantagakis, A. Trends in trawl and purse seine catch rates in the north-eastern Mediterranean. Mediterranean Marine Science 9, 49–66 (2008).
Aldebert, Y. Demersal resources of the Gulf of Lions (NW Mediterranean). Impact of exploitation on fish diversity. Vie et milieu 47, 275–284 (1997).
Ferretti, F., Myers, R. A., Serena, F. & Lotze, H. K. Loss of large predatory sharks from the Mediterranean Sea. Conservation Biology 22, 952–964 (2008).
STECF. Assessment of Mediterranean Sea stocks part II (STECF-14-08). (Luxembourg, 2013).
EC. Proposal for a Council regulation concerning measures for the sustainable exploitation of fishery resources in the Mediterranean Sea and amending regulations (EC) No 2847/93 and (EC) No 973/2001. Commission of the European Communities. Brussels, 9 October 2003, COM (2003) 589 final, 2003/0229 (CNS), 39 pp. (2003).
Garibaldi, L. The FAO global capture production database: a six-decade effort to catch the trend. Marine Policy 36, 760–768 (2012).
Katsanevakis, S. et al. Marine conservation challenges in an era of economic crisis and geopolitical instability: the case of the Mediterranean Sea. Marine Policy 51, 31–39 (2015).
Pauly, D. & Zeller, D. Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nature communications 7 (2016).
Libralato, S. et al. Food-web traits of protected and exploited areas of the Adriatic Sea. Biological Conservation 143, 2182–2194 (2010).
Pauly, D., Christensen, V., Dalsgaard, J., Froese, R. & Torres, F. Fishing down marine food webs. Science 279, 860–863 (1998).
Stergiou, K. Fisheries impact on trophic levels: long-term trends in Hellenic waters. 326−329 (Hellenic Centre for Marine Research (HCMR), Athens, 2005).
Stergiou, K. I. & Tsikliras, A. C. Fishing down, fishing through and fishing up: fundamental process versus technical details. Marine Ecology Progress Series 441, 295–301 (2011).
Micheli, F. et al. Cumulative Human Impacts on Mediterranean and Black Sea Marine Ecosystems: Assessing Current Pressures and Opportunities. PloS one 8, e79889 (2013).
EC. Directive 2014/89/EU of the European Parliament and of the Council of 23 July 2014 establishing a framework for maritime spatial planning. Official Journal of the European Union L 257/135. (2014).
Liquete, C., Piroddi, C., Macias, D., Druon, J.-N. & Zulian, G. Ecosystem services sustainability in the Mediterranean Sea: assessment of status and trends using multiple modelling approaches. Scientific Reports 6, 34162 (2016).
EC. Innovation in the Blue Economy: realising the potential of our seas and oceans for jobs and growth. Brussels, 13.5.2014 COM (2014) 254 final/2 (2014).
Piante, C. & Ody, D. Blue Growth in the Mediterranean Sea: the Challenge of Good Environmental Status. 192 (WWF, France, 2015).
Fulton, E. A. Approaches to end-to-end ecosystem models. Journal of Marine Systems 81, 171–183 (2010).
Duarte, C. M., Kennedy, H., Marbà, N. & Hendriks, I. Assessing the capacity of seagrass meadows for carbon burial: current limitations and future strategies. Ocean & coastal management 83, 32–38 (2013).
Coll, M. et al. The scientific strategy needed to promote a regional ecosystem-based approach to fisheries in the Mediterranean and Black Seas. Reviews in fish biology and fisheries 23, 415–434 (2013).
Allen, J., Holt, J. T., Blackford, J. & Proctor, R. Error quantification of a high-resolution coupled hydrodynamic-ecosystem coastal-ocean model: Part 2. Chlorophyll-a, nutrients and SPM. Journal of Marine Systems 68, 381–404 (2007).
Heymans, J. J., Coll, M., Libralato, S., Morissette, L. & Christensen, V. Global patterns in ecological indicators of marine food webs: a modelling approach. PloS one 9, e95845 (2014).
Kempton, R. & Taylor, L. Models and statistics for species diversity. Nature 262, 818–820 (1976).
Ainsworth, C. H. & Pitcher, T. J. Modifying Kempton’s species diversity index for use with ecosystem simulation models. Ecological Indicators 6, 623–630 (2006).
Shin, Y.-J. et al. Using indicators for evaluating, comparing, and communicating the ecological status of exploited marine ecosystems. 2. Setting the scene. ICES Journal of Marine Science: Journal du Conseil 67, 692–716 (2010).
Pauly, D. & Watson, R. Background and interpretation of the ‘Marine Trophic Index’as a measure of biodiversity. Philosophical Transactions of the Royal Society B: Biological Sciences 360, 415–423 (2005).
Bundy, A. et al. The good (ish), the bad, and the ugly: a tripartite classification of ecosystem trends. ICES Journal of Marine Science: Journal du Conseil 67, 745–768 (2010).
Acknowledgements
M.C. acknowledges support by the European Commission through the Marie Curie Career Integration Grant Fellowships – PCIG10-GA-2011-303534 - to the BIOWEB project. RD, MC and JS were partially funded by the European Union’s Horizon research and innovation programme grant agreement No 689518 for the MERCES project.VC acknowledges support through NSERC Discovery Grant RGPIN-2014-05782.
Author information
Authors and Affiliations
Contributions
C.P. conceived the study design, developed the models, acquired the data, analysed and interpreted the data. M.C., C.L. and V.C. helped in developing the models, data analysis and interpretation. D.M., R.D., K.G. provided data respectively on primary production (from the biogeochemical model), deep fish and benthos, and fishing effort. J.B. and J.S. helped with the programming code of the software. C.P. led the drafting of manuscript with the contribution and critical revision from all the authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Piroddi, C., Coll, M., Liquete, C. et al. Historical changes of the Mediterranean Sea ecosystem: modelling the role and impact of primary productivity and fisheries changes over time. Sci Rep 7, 44491 (2017). https://doi.org/10.1038/srep44491
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep44491
This article is cited by
-
Importance of Copepods in the Diet of Sardina pilchardus and Sardinella aurita: Preliminary Investigation in Bou Ismail Bay (Algerian Basin - SW Mediterranean Sea)
Thalassas: An International Journal of Marine Sciences (2024)
-
Temporal changes in the diet composition of Shorttail Yellownose skate, when exposed to overfishing conditions in northern and central Patagonia, Argentina
Hydrobiologia (2024)
-
Global-scale parameters for ecological models
Scientific Data (2023)
-
Assessment of interactive acoustic deterrent devices set on trammel nets to reduce dolphin–fishery interactions in the Northern Tyrrhenian Sea
Scientific Reports (2023)
-
Shifts in the pelagic fishery dynamics in response to regional sea warming and fishing in the Northeastern Mediterranean
Regional Environmental Change (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
