
Abstract
Humans, like all organisms, are subject to fundamental biophysical laws. Van Valen predicted that, because of zero-sum dynamics, all populations of all species in a given environment flux the same amount of energy on average. Damuth’s ’energetic equivalence rule’ supported Van Valen´s conjecture by showing a tradeoff between few big animals per area with high individual metabolic rates compared to abundant small species with low energy requirements. We use metabolic scaling theory to compare variation in densities and individual energy use in human societies to other land mammals. We show that hunter-gatherers occurred at densities lower than the average for a mammal of our size. Most modern humans, in contrast, concentrate in large cities at densities up to four orders of magnitude greater than hunter-gatherers, yet consume up to two orders of magnitude more energy per capita. Today, cities across the globe flux greater energy than net primary productivity on a per area basis. This is possible by importing enormous amounts of energy and materials required to sustain hyper-dense, modern humans. The metabolic rift with nature created by modern cities fueled largely by fossil energy poses formidable challenges for establishing a sustainable relationship on a rapidly urbanizing, yet finite planet.
Similar content being viewed by others

Introduction
All populations, including humans, are sustained by fluxes of energy and materials from a finite environment. Physical constraints on biological design result in ubiquitous and predictable allometric scaling laws1. These take power law form where some trait of interest (R), scales with body size (M),

where β is the exponent and c is the intercept. Metabolic scaling theory predicts quarter-power exponents for rates and quantities across many levels of biological organization2,3 including whole organism and mass-specific metabolic rates, which scale as β ≈ ¾ and β ≈ −¼, respectively. Allometric parameters (i.e., intercept and slope) can be predicted theoretically and evaluated empirically to form a quantitative framework to carry out meaningful comparisons across scales from cells and organisms3 to human societies4,5. Using this framework as a reference we aimed to understand unique aspects of human ecology and to quantify the extent to which the human species has departed from the energetic constraints that keep all other species in check.
An important ecological implication of metabolic scaling is the inverse relationship between body size and density. Because individual metabolic rate (Ei) scales predictably with size2

the maximum number of individuals per unit area (Dmax), scales as the inverse of individual energy demands6,7

The result is a tradeoff between size and abundance. The scatter around the best-fit line reflects environmental fluctuations linked to biotic interactions and temporal and spatial heterogeneity in resource availability, which appear as positive and negative deviations from maximum densities. Abundance per se is not limited by body mass but instead by the energy required to support an individual, Ei, of a given body size. Rearranging the allometric relationships in Eqn 2 and 3, theory predicts that

and population energy flux, Ep, calculated as the product of Ei · Dmax, is invariant across species, with β = 0

This ‘energetic-equivalence rule’ (EER6,7) links individual metabolic requirements to population energy use in space and time. This is consistent with the existence of a zero sum game for energy use as predicted by Van Valen8,9 and quantified in local mammal communities10. Unique to industrial humans, however, is the capacity to harness extra-metabolic energy in the form of renewables and fossil fuels to power modern agricultural-technological-industrial lifestyles. Among human societies, individual energy consumption varies from ~120 watts of biological metabolism — the equivalent of ~2500 kilocalories per day — in hunter-gatherers to more than 10,000 watts in the most developed nations5,11,12. So clearly, humans have deviated from other species in their energy use.
Results
Figure 1 shows that herbivorous land mammals support theoretical predictions where density decreases proportionally with individual energy requirements (slope = −1.08; 95% CI: −0.88, −1.27). Hunter-gatherers, in contrast, occur at densities lower than expected based on other land mammals (ANOVA, F-interaction = 6.37, p < 0.001, Tukey post-hoc test). The trophic position, where an organism feeds in the food web, explains additional variation in the densities of land mammals and hunter-gatherers where densities decrease with higher trophic levels (Supplemental materials; Table S1).

Red circles represent vegetarian hunter-gatherers (n = 31), blue circles are pre-industrial societies (n = 4), purple circles are modern cities (n = 163), and green triangles are other herbivorous land mammal species (n = 74). Note that the slope for cities is shallower than herbivorous land mammals, which support theoretical predictions of −1. See supplemental materials for additional details and data sources.
Across cities, density scales negatively with increasing per capita energy requirements consistent with theoretical predictions and similar to the scaling of land mammals. However, modern city dwellers occur at densities that are four orders of magnitude greater than hunter-gatherers and other land mammals (Fig. 1; Table S1) even though they consume one to two-orders of magnitude greater energy per capita. The highest density city in our data (Dhaka, Bangladesh with 44,000 ind/km2) now surpasses the highest density wild rodent (Townsend’s vole with 34,349 ind/km2). The slope for urban humans (−0.44; 95% CI: −0.32, −0.55) is shallower than the theoretical expectation (slope of −1) and estimated empirical slope from land mammals (Table S1).
Consistent with Van Valen’s zero-sum prediction and the EER, the energy flux by herbivorous land mammal populations (10−1 to 104 watts/km2) is invariant with individual energy use (Fig. 2) with a slope indistinguishable from 0 (−0.08, 95% CIs: −0.28, 0.12). Energy flux by hunter-gatherers range from 102 to 103 watts/km2 and is lower than other land mammals (ANOVA, F-value interaction = 10.01, p < 0.001, Tukey post-hoc test). Members of pre-industrial societies fluxed greater energy (104 to 105 watts/km2) than hunter-gatherers and other land mammals on average but less than cities. Energy flux in modern cities ranges from 105 to 108 watts/km2 and surpasses global primary productivity on land (105 watts/km2 global avg). Unique to urban humans is the positive relationship between population energy flux and per capita energy requirements (slope = 0.56 [95% CI: 0.44, 0.68]), whereas other land mammals show a slope indistinguishable from 0 as theory predicts (Fig. 2). See supplemental for additional analyses by trophic levels.

Red circles represent vegetarian hunter-gatherers (n = 31), blue circles are pre-industrial societies (n = 4), purple circles are modern cities (n = 163), and green triangles are other herbivorous land mammal species (n = 74). Ep is estimated as the product of density (individuals/km2) and Ei (watts). Note that population energy use for herbivorous land mammals does not vary with individual energy use supporting theoretical expectations (slope = 0), whereas urban cities increase (positive slope). The dashed line represents the terrestrial average net primary productivity for the planet from23.
Discussion
Just like all mammals, hunter-gatherers are faced with the challenge of meeting metabolic demands from the local environment to power their lifestyles and sustain their populations. Pre-industrial societies lived at greater densities than hunter-gatherers through the use of agriculture. Humans in modern cities live at densities much greater than those in hunter-gatherers and pre-industrial societies and consume up to two orders of magnitude greater energy per capita than caloric needs alone. This shows that the rapid rise in human densities, which has occurred in less than 10,000 years, is coincident with innovations in food production and extra-metabolic energy use from renewable and fossil fuels13.
One salient characteristic of complex human social systems is our ability to copy the behavior of others to propel a cumulative cultural evolution (CCE14). It is easy to see that this process likely speeds up with greater population density and information flow as a result of greater energy throughput. The positive slope for population energy flux with per capita energy use in Fig. 2 is a consequence of a shallower slope in Fig. 1. This suggests that increased per capita energy use – which is tightly coupled to economic growth5,12 – results in economies of scale by packing more individuals in a given area. It is hypothesized that population density—through its effect on CCE—is a major driver of innovation4,15 and increased social complexity16. It is possible that these processes generate positive feedbacks and a runaway process of cultural niche construction17 contributing to the rapid divergence of humans from other species and the rise in human densities from hunter-gatherers to agriculturalists to modern cities. We propose that this unique aspect of human ecology is a result of the Malthusian-Darwinian dynamic that drives species to maximize power when innovations allow18.
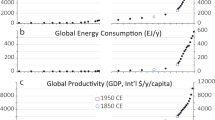
Throughout this process human societies have become increasingly decoupled from local environmental constraints and uncertainties. Technologies that increase resource production and global trade networks that offset imbalances in resource supply and demand provide an enormous buffer from local environmental constraints and perturbations such as drought and other human and natural disasters19. However, hyper-dense cities are only possible by the fluxes of vast quantities of energy, materials, and information across city boundaries in order to offset resource sinks and maintain dense, urban lifestyles20. The energy flux required to sustain hyper-dense cities now surpasses baseline levels of net primary productivity on a per area basis (Fig. 2) and is largely (~85% globally) in the form of carbon-based fossil fuels5. Increasing scarcity of essential resources including fossil fuels5, water21, and nutrients such as phosphorous20 pose formidable challenges for continued urbanization and high-density cities. The cumulative impact of hyper-dense cities may surpass planetary tipping points22 having rippling effects at multiple scales.
Our human macroecological approach offers illuminating insights into how humans have rapidly diverged from other species in the course of our unique CCE. Through the use of extra-metabolic energy, modern humans have escaped the energetic constraints of primary productivity that are imposed on all other species. Our approach also highlights the extraordinary densities that humans have obtained through the continued capacity to harness energy reserves from the planet in the form of sunlight23 to energy stored on geological time scales24. Whether density-dependent innovations will continue to outpace resource constraints on human population is uncertain. What is certain is that the steep metabolic rift with nature25,26 created by the vast extra-metabolic energy subsidies required to support growing, hyper-dense cities poses formidable challenges to achieving sustainability in a post-fossil fuel world.
Methods
Data
We use global data from ecological, archeological, demographic, and economic sources (Supplemental data). Data for metabolic rates and densities for 249 land mammals are from the PANTHERIA species-level database27. Densities for 339 hunter-gatherers are available in Binford’s comparative ethnographic and environmental database28. Hunter-gatherer lifestyles are powered by biological metabolism estimated to be 120 watts (~2500 kilocalories/day) based on established allometries for primates (following refs 5 and 11). Densities for pre-industrial societies (n = 4) are from29 and energy use estimated at 600 watts11.
The EER applies to primary consumers6,7, so we plot herbivorous land mammals (n = 74) and hunter-gatherers that obtain greater than two-thirds of their diet from plants (i.e., gatherers, N = 31) in Figs 1 and 2. It is well known that species at higher trophic levels occur at lower densities (e.g., refs 30 and 31) and hunter-gatherers are no exception with gatherers occurring at higher densities than omnivores, which occur at higher densities than carnivores (See supplemental materials for additional trophic analyses).
It has been suggested that the EER predicts maximum animal densities32,33. So, we use the densest city per country for which data are available from Demographia World Urban Areas (demographia.com; website has updated lists). The data consists of censuses conducted between 2000 and 2014. In contrast to hunter-gatherers, urban human lifestyles are powered by both biological metabolism and extra-metabolic energy in the form of renewables and fossil fuels11,34. However, a global database on metabolic (i.e., caloric) and extra-metabolic energy use for cities is not available. So we estimated per capita energy use by combining country-level data on food consumption per country (kcal per capita per day) from the Food and Agriculture Organization from of the United Nations (FAO) (http://faostat3.fao.org/) with extra-metabolic energy from The World Bank Indicators (http://data.worldbank.org/indicator; website has updated data). We use only one city (the densest) per country in our analyses although more cities are listed in the demographia dataset. This assumes greater variation among countries than within, which is supported by studies showing that resource use and waste production scale linearly (e.g., constant per capita) with city size within countries4,35.
Statistical Analyses
We determined the relationship between population density and per capita energy requirements separately for land mammals and urban humans using ordinary least squares regression of log transformed variables. We compared 95% confidence intervals of allometric parameters from linear models of the log-log relationships for land mammals and modern cities in order to evaluate theoretical expectations (slope = −1).
We conducted Analysis of Covariance (ANCOVA), with per capita energy use as the covariate, density as the dependent variable, and trophic level or human versus land mammal as fixed factors. Comparisons were made between: (i) land-mammals by trophic levels (ANCOVA), (ii) land-mammals and hunter-gatherers considering trophic level as a second factor (ANOVA), and (iii) land mammals (with trophic level as a cofactor) versus urban humans (ANCOVA).
We calculated population energy requirements (Ep) as the product of population density and per capita energy use to test the EER that all populations flux the same amount of energy per unit area as a consequence of the zero-sum. We conducted ANCOVAs and ANOVAS similar to above with Ep as the dependent variable.
Additional Information
How to cite this article: Burger, J. R. et al. Extra-metabolic energy use and the rise in human hyper-density. Sci. Rep. 7, 43869; doi: 10.1038/srep43869 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
West, G. B., Brown, J. H. & Enquist, B. J. A general model for the origin of allometric scaling laws in biology. Science 276, 122–126 (1997).
Brown, J. H., Gillooly, J. F., Allen, A. P., Savage, V. M. & West, G. B. Toward a metabolic theory of ecology. Ecology 85, 1771–1789 (2004).
Sibly, R. M., Brown, J. H. & Kodric-Brown, A. Metabolic ecology: a scaling approach. (John Wiley & Sons, 2012).
Bettencourt, L. M., Lobo, J., Helbing, D., Kühnert, C. & West, G. B. Growth, innovation, scaling, and the pace of life in cities. Proceedings of the National Academy of Sciences. 104, 7301–7306 (2007).
Brown, J. H. et al. Energetic limits to economic growth. BioScience. 61, 19–26 (2011).
Damuth, J. Population density and body size in mammals. Nature. 290, 699–700 (1981).
Damuth, J. Interspecific allometry of population density in mammals and other animals: the independence of body mass and population energy‐use. Biological Journal of the Linnean Society. 31, 193–246 (1987).
Van Valen, L. Body size and numbers of plants and animals. Evolution. 27–35 (1973).
Van Valen, L. Evolution as a zero-sum game for energy. Evolutionary Theory. 4, 289–300 (1980).
Ernest, S. M., Brown, J. H., Thibault, K. M., White, E. P. & Goheen, J. R. Zero Sum, the Niche, and Metacommunities: Long‐Term Dynamics of Community Assembly. The American Naturalist. 172, E257–E269 (2008).
Moses, M. E. & Brown, J. H. Allometry of human fertility and energy use. Ecology Letters. 6, 295–300 (2003).
Brown, J. H. et al. Macroecology meets macroeconomics: Resource scarcity and global sustainability. Ecological Engineering. 65, 24–32 (2014).
Krausmann, F., Fischer‐Kowalski, M., Schandl, H. & Eisenmenger, N. The global sociometabolic transition. Journal of Industrial Ecology. 12, 637–656 (2008).
Boyd, R., Richerson, P. J. & Henrich, J. The cultural niche: Why social learning is essential for human adaptation. Proceedings of the National Academy of Sciences. 108, 10918–10925 (2011).
Henrich, J. Demography and cultural evolution: how adaptive cultural processes can produce maladaptive losses: the Tasmanian case. American Antiquity. 197–214 (2004).
Marquet, P. A. et al. Emergence of social complexity among coastal hunter-gatherers in the Atacama Desert of northern Chile. Proceedings of the National Academy of Sciences. 109, 14754–14760 (2012).
Rendell, L., Fogarty, L. & Laland, K. N. Runaway cultural niche construction. Philosophical Transactions of the Royal Society of London B: Biological Sciences. 366, 823–835 (2011).
Nekola, J. C. et al. The Malthusian–Darwinian dynamic and the trajectory of civilization. Trends in Ecology & Evolution. 28, 127–130 (2013).
Hammond, S. T. et al. Food Spoilage, Storage, and Transport: Implications for a Sustainable Future. BioScience. 65, 758–768 (2015).
Burger, J. R. et al. The macroecology of sustainability. PLoS Biology. 10, e1001345 (2012).
Gleick, P. H. & Palaniappan, M. Peak water limits to freshwater withdrawal and use. Proceedings of the National Academy of Sciences. 107, 11155–11162 (2010).
Barnosky, A. D. et al. Approaching a state shift in Earth’s biosphere. Nature. 486, 52–58 (2012).
Haberl, H. et al. Quantifying and mapping the human appropriation of net primary production in Earth’s terrestrial ecosystems. Proceedings of the National Academy of Sciences. 104, 12942–12947 (2007).
Schramski, J. R., Gattie, D. K. & Brown, J. H. Human domination of the biosphere: Rapid discharge of the earth-space battery foretells the future of humankind. Proceedings of the National Academy of Sciences. 112, 9511–9517 (2015).
Marx, K. Capital: a critique of political economy, 3 vols (1967).
Foster, J. B. Marx’s Theory of Metabolic Rift: Classical Foundations for Environmental Sociology 1. American Journal of Sociology. 105, 366–405 (1999).
Jones, K. E. et al. PanTHERIA: a species‐level database of life history, ecology, and geography of extant and recently extinct mammals. Ecology. 90, 2648–2648 (2009).
Binford, L. R. Constructing frames of reference: an analytical method for archaeological theory building using ethnographic and environmental data sets. (Univ of California Press, 2001).
Zimmermann, A., Hilpert, J. & Wendt, K. P. Estimations of population density for selected periods between the Neolithic and AD 1800. Human Biology 81, 357–380 (2009).
Carbone, C. & Gittleman, J. L. A common rule for the scaling of carnivore density. Science 295, 2273–2276 (2002).
Marquet, P. A. Of predators, prey, and power laws. Science. 295, 2229–2230 (2002).
Belgrano, A., A. P. Allen, B. J. Enquist & J. R. Gillooly . Allometric scaling of maximum population density: a common rule for marine and phytoplankton and terrestrial plants. Ecology Letters. 5, 6-11-613 (2002).
Marquet, P. A., Navarrete, S. A. & Castilla, J. C. Body size, population density, and the energetic equivalence rule. Journal of Animal Ecology. 325–332 (1995).
Burger, O., DeLong, J. P. & Hamilton, M. J. Industrial energy use and the human life history. Scientific reports. 1 (2011).
Fragkias, M., Lobo, J., Strumsky, D. & Seto, K. C. Does size matter? Scaling of CO2 emissions and US urban areas. PLoS One. 8, e64727 (2013).
Acknowledgements
We thank Fernando Alfaro, Bill Burnside, Jim Brown, Melanie Moses, Allen Hurlbert and members of the Human Macroecology Group at UNM for helpful suggestions and discussion. Munik Shrestha brought to our attention a typo in equation 2 in an earlier version of the unpublished manuscript posted to arXiv.org. An earlier version of Figure 1 was prompted by talks and discussion at the Santa Fe Institute’s and Universidad de Desarrollo’s Complex Systems Summer School in Zapallar, Chile. JRB is supported by a Carolina Postdoctoral Fellowship for Faculty Diversity. VPW is supported by Programa Capital Humano Avanzado from CONICYT. PAM acknowledges support from Proyectos IEB ICM-MINECON, P05-002 and PFB-23.
Author information
Authors and Affiliations
Contributions
All authors designed the study. J.R.B. and V.P.W. compiled and analyzed the data. All authors contributed to writing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Burger, J., Weinberger, V. & Marquet, P. Extra-metabolic energy use and the rise in human hyper-density. Sci Rep 7, 43869 (2017). https://doi.org/10.1038/srep43869
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep43869
This article is cited by
-
City-scale energetics: window on adaptive thermal insulation in North American cities
Journal of Comparative Physiology B (2022)
-
Diversity begets diversity in mammal species and human cultures
Scientific Reports (2020)
-
The Central Role of Energy in the Urban Transition: Global Challenges for Sustainability
BioPhysical Economics and Resource Quality (2019)
-
Declining Country-Level Food Self-Sufficiency Suggests Future Food Insecurities
BioPhysical Economics and Resource Quality (2019)
-
An Updated Theoretical Framework for Human Sexual Selection: from Ecology, Genetics, and Life History to Extended Phenotypes
Adaptive Human Behavior and Physiology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
