
Abstract
Ammonia is both a building block and a breakdown product of amino acids and is found widely in the environment. The odor of ammonia is attractive to many insects, including insect vectors of disease. The olfactory response of Drosophila to ammonia has been studied in some detail, but the taste response has received remarkably little attention. Here, we show that ammonia is a taste cue for Drosophila. Nearly all sensilla of the major taste organ of the Drosophila head house a neuron that responds to neutral solutions of ammonia. Ammonia is toxic at high levels to many organisms, and we find that it has a negative valence in two paradigms of taste behavior, one operating over hours and the other over seconds. Physiological and behavioral responses to ammonia depend at least in part on Gr66a+ bitter-sensing taste neurons, which activate a circuit that deters feeding. The Amt transporter, a critical component of olfactory responses to ammonia, is widely expressed in taste neurons but is not required for taste responses. This work establishes ammonia as an ecologically important taste cue in Drosophila, and shows that it can activate circuits that promote opposite behavioral outcomes via different sensory systems.
Similar content being viewed by others
Introduction
Ammonia plays a critical role in biological systems. In the early Earth, ammonia likely combined with water, methane, and hydrogen in the presence of lightning bolts to produce simple organic compounds essential to the first forms of life1. Ammonia is an essential building block of amino acids, and it is a breakdown product of amino acids. It is found widely in the environment: in air, in waters, in soils, and in all forms of life2.
Ammonia is an olfactory cue for many organisms, including insect vectors of human disease. Anopheles mosquitoes that transmit malaria are attracted to ammonia3,4, which is a component of human sweat5, and ammonia is an effective attractant in Anopheles traps6. Aedes mosquitoes, which transmit Zika, dengue fever and yellow fever, are also attracted to the odor of ammonia7, as are insects that transmit Chagas disease8. Ammonia also attracts the widely used genetic model organism Drosophila and several other insect species9,10,11,12,13, perhaps because ammonia odor indicates protein-rich food sources10. The molecular and cellular mechanisms underlying the olfactory response to ammonia have been investigated in the antenna of the fly and the mosquito14,15,16,17.
The taste response to ammonia, by contrast, is largely unexplored. In principle, the ammonia in human sweat might elicit an appetitive response via the taste organs of mosquitoes. On the other hand, high levels of ammonia are toxic to most organisms18,19,20, so ammonia might elicit an aversive taste response. There is evidence that mice avoid ingestion of high concentrations of ammonium chloride21, and ammonium chloride has a bitter and salty taste to humans22,23. However, there has been remarkably little study of the taste response to ammonia in insects.
The major taste organ of the Drosophila head is the labellum, which contains 31 taste sensilla24. These sensilla divide into three morphological classes: long (L), intermediate (I), and short (S)25,26. Each individual sensillum is identified by its position on the labellum, e.g. L1. When a sensillum makes contact with a food source, tastants diffuse through a pore in the tip of the sensillum and activate neurons within24. For example, some taste neurons are activated by sugars and promote an appetitive response, while other neurons are activated by bitter compounds and elicit an aversive response. L sensilla exhibit strong electrophysiological responses to sugars27, but not to bitter compounds; I and S sensilla show strong responses to bitter compounds26. The legs, pharynx, and wings also contain taste neurons24,28,29,30.
Here we ask whether ammonia elicits a taste response from Drosophila. We find that ammonia elicits a strong physiological response from one class of labellar taste sensilla, the S sensilla, and weaker responses from others. Whereas ammonia elicits an attractive response via the olfactory system, we find that it elicits an aversive behavioral response in two taste paradigms. We then explore the cellular and molecular basis of the taste response. We find that physiological responses of the S sensilla and behavioral responses both depend on bitter-sensing neurons that activate an aversive circuit. We then demonstrate that Amt, an essential molecular component of ammonia response in the olfactory system, is also expressed in the taste system, although in a different cell type. Surprisingly, while Amt is required for the olfactory response to ammonia, it is not required for the taste response, indicating differences in the underpinnings of ammonia detection in olfaction and taste.
Results
Taste sensilla respond to ammonia
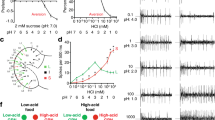
We systematically tested all 31 of the taste sensilla on the labellum for response to ammonia. We used electrophysiological tip recordings to determine whether a solution of ammonium chloride (NH4Cl) at neutral pH elicited action potentials from neurons in any of the sensilla. We initially tested NH4Cl at a concentration of 100 mM, which is well within the range found in ammonia-rich sources in natural environments31,32. We note that ammonia exists primarily in gaseous form at room temperature, but dissolves readily in water. At pH 7 most ammonia molecules are protonated, i.e. NH4+, but for simplicity we will refer to both NH3 and NH4+ as “ammonia”.
This NH4Cl stimulus elicited strong responses from most S sensilla, and weaker responses from some L and I sensilla (Fig. 1A). We tested two additional stimuli on the same sensilla to determine if the NH4Cl responses are in fact due to ammonia, as opposed to the chloride ion. We found that applying an equivalent concentration of ammonia using NH4 sulfate, i.e. (NH4)2SO4, at pH 7 also elicited strong responses from most S sensilla and weaker responses from L and I sensilla. In contrast, 100 mM choline chloride (ChCl), which contains chloride but not ammonia, elicited weaker responses than NH4Cl from all sensilla, indicating that the neurons do not respond to chloride ions. The diluent alone, a solution of tricholine citrate (TCC)33, produced little if any response from most of these sensilla (Fig. 1A).

(A) Mean responses in spikes/s per individual sensillum type. Responses to the diluent, TCC, along with responses to NH4Cl, NH4 sulfate, and ChCl are displayed. Values for the diluent were not subtracted from values for the other compounds. NH4Cl and NH4 sulfate solutions were neutralized to pH 7 with NH4OH. n values are provided in Table S1. (B) Mean responses of each morphological class of sensilla. S4 and S8 sensilla were excluded. n = 44–58 (C) Taste sensilla respond to ammonia in a dose-dependent manner. “0 mM NH4Cl” indicates the response to TCC alone. n = 8–52.(D) Representative electrophysiological traces. Scale bar indicates 1 mV, 100 ms.
When the responses were grouped by morphological type (S, L, I), each type gave stronger responses to NH4Cl or NH4 sulfate than to the control stimuli (Fig. 1B; p < 0.02). Moreover, in each sensillum type the responses to NH4Cl and NH4 sulfate were indistinguishable (p > 0.05). The simplest interpretation of these results is that labellar taste sensilla respond to ammonia.
We note that S4 and S8, which differ from other S sensilla in their lack of response to bitter compounds26, also differ in their lack of strong responses to NH4Cl or NH4 sulfate (Fig. 1A). Accordingly, S4 and S8 were excluded from the analysis shown in Fig. 1B and from further analysis of S sensilla.
The responses to ammonia were confirmed and extended by testing NH4Cl across a range of concentrations spanning four orders of magnitude (Fig. 1C). Responses were dose-dependent, with response thresholds between 10 mM and 100 mM for each sensillum type. Response magnitudes of S sensilla were higher than those of L or I sensilla at all concentrations above the threshold. Representative traces of responses to increasing NH4Cl concentrations are shown in Fig. 1D.
Ammonia is an aversive taste cue
What is the valence of ammonia taste? Ammonia odor is attractive to many insects, perhaps serving as a cue for protein sources10,34. However, high concentrations of ammonia are toxic, suggesting the need for a mechanism to avoid their ingestion.
To examine the valence of ammonia taste we used two distinct behavioral paradigms. First we used a CAFÉ assay in which small groups of starved flies were given 4 hours to feed from either of two capillary tubes, one containing sucrose alone and the other containing sucrose mixed with NH4Cl35. We found that flies preferred solutions containing sucrose alone to those containing sucrose and NH4Cl at levels greater than 30 mM (Fig. 2A).

(A) Top: CAFÉ assay. Starved flies are housed in a conical tube for 4 h with access to two capillary tubes, one containing 100 mM sucrose alone and the other containing 100 mM sucrose and concentrations of NH4Cl ranging from 1 mM to 300 mM. The amount of solution removed from each tube is measured and a preference index (PI) is calculated; a negative PI indicates a preference for sucrose alone. Bottom: flies prefer solutions of sucrose alone to sucrose solutions containing high concentrations of NH4Cl (n = 12–28). (B) Top: pharyngeal pumping assay. A fly restrained in a pipette tip is presented with a drop of a solution. The time spent feeding from the drop is measured. Bottom: addition of increasing concentrations of NH4Cl reduced the time spent feeding (n = 20).
Next we examined feeding over a much shorter time span, using a pharyngeal pumping assay36. Starved flies were presented with a water droplet, and the duration of ingestion was measured by visual inspection. Adding 10 mM NH4Cl or more to the droplet decreased feeding time, with greater doses producing shorter feeding periods (Fig. 2B).
In summary, NH4Cl had a negative valence in two taste paradigms, one extending over hours and the other over seconds. To confirm our interpretation of these results and to extend our analysis of ammonia taste response, we next investigated its cellular basis.
Taste response to ammonia requires Gr66a+ neurons
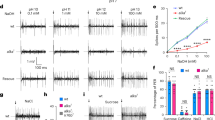
Which neurons are required in gustatory sensilla for the ammonia response? Previous work has supported a model in which S and I sensilla, but not L sensilla, contain a neuron that expresses the bitter receptor Gr66a, responds to bitter compounds, and activates an aversion circuit25,26,37,38. Since ammonia appeared aversive in two behavioral paradigms, we hypothesized that Gr66a-expressing “bitter” neurons functioned in ammonia response. To test this hypothesis we ablated bitter neurons by using a Gr66a-GAL4 construct to drive expression of diptheria toxin (UAS-DTA) and then measured neuronal spiking responses to ammonia. To confirm the ablation we also measured responses to berberine, a well-characterized bitter compound.
Physiological analysis of S sensilla from control flies containing either Gr66a-GAL4 or UAS-DTA alone produced strong responses to both berberine and NH4Cl (Fig. 3A,C). However, S sensilla of flies containing both constructs showed virtually no response to either berberine or NH4Cl. The same conclusion was reached with each of two concentrations of NH4Cl. The simplest interpretation of these results is that the ammonia responses of S sensilla require the Gr66a+ neuron.

(A) Ablation of bitter-sensitive Gr66a+ neurons with a Gr66a-GAL4 driver and UAS-DTA (diphtheria toxin) severely reduces responses of S sensilla to the bitter compound berberine (1 mM) and to NH4Cl (n = 22–37, p < 0.0001). Parental control lines respond strongly to these stimuli. (B) Ablation of sugar-sensitive Gr64f+ cells severely reduces the response to 100 mM sucrose in L sensilla (p < 0.0001), but does not affect responses to NH4Cl (n = 22–27). (C,D) Representative electrophysiological traces. Scale bar indicates 1 mV, 100 ms.
We also ablated neurons that express the sugar receptor Gr64f, respond to sugars, and activate an appetitive response27,39. For this experiment we examined L sensilla, whose sucrose responses are much more robust and consistent in our hands than those of S sensilla. Physiological analysis of sensilla from control flies containing Gr64f-GAL4 alone, or UAS-DTA alone, produced strong responses to sucrose (Fig. 3B,D). However, sensilla of flies containing both constructs showed a dramatically reduced response to sucrose, confirming successful ablation of the sugar neuron. Responses to NH4Cl were the same in all three genotypes, at each of two different NH4Cl concentrations. The simplest interpretation of these results is that these ammonia responses do not depend on sugar neurons.
Behavioral analysis was carried out to determine whether aversion of the fly to ammonia depended on Gr66a+ taste neurons. We used the pharyngeal pumping paradigm, which measures the response to NH4Cl and does not entail the use of sucrose, as does the CAFÉ assay. We found that in the absence of NH4Cl, feeding time on a water droplet was equivalent in flies containing Gr66a-GAL4 alone, UAS-DTA alone, or Gr66a-GAL4; UAS-DTA (Fig. 4). However, in the presence of NH4Cl, flies in which Gr66a+ cells were ablated showed increased feeding time compared to the parental controls. These results indicate that bitter neurons are required for the aversive, anti-feedant response to NH4Cl.

Flies in which Gr66a+ bitter-sensitive neurons were ablated with Gr66a-GAL4 and UAS-DTA spent the same amount of time feeding on a water droplet as parental controls. Flies in which Gr66a+ cells were ablated spent longer feeding on 100 mM NH4Cl than parental controls (p < 0.001; one-way ANOVA, Bonferroni post-test, n = 26).
The molecular underpinnings of ammonia response differ between taste and olfaction
We recently identified in Drosophila an ammonia transporter, Amt, which is expressed in a class of olfactory sensilla that respond strongly to airborne ammonia15. Amt is expressed in auxiliary cells of these sensilla. Analysis of an insertion mutation revealed a requirement for Amt in the electrophysiological response of these olfactory sensilla to ammonia.
We found that an Amt-GAL4 construct drives expression of a UAS-GFP reporter in two taste organs: the labellum and legs (Fig. 5A,B), in addition to the antenna. Surprisingly, in these taste organs Amt expression was detected in neurons, identified by the presence of labeled dendrites and axons, rather than in auxiliary cells. To confirm the expression of Amt in taste neurons, we generated an antibody to the 148 amino acids of the C-terminal tail of Amt. This antibody labeled neurons in taste sensilla of the labellum, confirming the results obtained with the Amt-GAL4 driver (Fig. 5C). The specificity of the antibody was confirmed by testing it against the Amt1 insertion mutant: labeling was greatly reduced in the labellum (Fig. 5D). Labeling was not completely eliminated, however, presumably reflecting the presence of some residual Amt expression in the insertion mutant.

Amt-GAL4 drives UAS-mCD8::GFP in taste neurons in (A) the labellum and (B) legs, as visualized in whole mount preparations. (C) An αAMT antibody labels cells in labellar sections of wiso control flies, but (D) most staining is lost in Amt1. (E,F) Labeling is eliminated in the deletion allele Amt2 compared to the control wCS. (G) In Amt+ flies, all neurons that express GFP driven by Amt-GAL4 are also labeled by the αAMT antibody. The αAMT antibody also labels some cells that show weak or no labeling with Amt-GAL4.
Using CRISPR-Cas9 technology we generated Amt2 40, in which most of the protein-coding sequence of the gene, including 8 of 11 transmembrane domains, are deleted (Fig. S1). The mutation was outcrossed against a control wCS stock for five generations. The Amt2 labellum revealed no labeling with the anti-Amt antibody, providing further confirmation of the antibody’s specificity (Fig. 5E,F). In Amt+ animals, all neurons labeled by Amt-GAL4; UAS-GFP were also labeled by the antibody (Fig. 5G), as expected of a faithful GAL4 driver; a small fraction of neurons labeled with the antibody were labeled weakly if at all by the driver, as often found for drivers that are not expressed as strongly as the genes they represent.
Amt-GAL4 is expressed in a single neuron per sensillum on the labellum. All or nearly all sensilla contain a neuron that expresses the driver (Fig. 5A).
Is Amt required for gustatory response to ammonia, as it is for olfactory response? We were surprised to find that Amt2 showed electrophysiological responses similar to those of the control line across a broad concentration range, in S, L, and I sensilla (Fig. 6A). Amt2 also showed a normal behavioral response in a CAFÉ assay to 100 mM NH4Cl, a concentration that elicits an intermediate response in wild type flies (Fig. 6B). Finally, Amt2 had a normal response in the pharyngeal pumping paradigm when tested with either water or ammonia (Fig. 6C).

(A) Amt2 flies have electrophysiological responses to NH4Cl that are not significantly different from responses of wCS controls (+). (B) Amt2 flies show the same preference for sucrose alone (0 mM NH4Cl) versus sucrose with 100 mM NH4Cl as controls in the CAFÉ assay (n = 47–57). (C) In the pharyngeal pumping assay, the time spent feeding on 100 mM NH4Cl by Amt2 flies is not significantly different from wCS controls (+) (n = 25–31).
These results prompted us to map Amt expression in more detail. We carried out double-label experiments using Amt-GAL4 and markers of sugar (Gr5a-LexA) or bitter (Gr66a-RFP) neurons. These experiments revealed expression of Amt in sugar neurons, but not bitter neurons (Fig. S2). This mapping is consistent with our finding that neither loss of Amt nor ablation of sugar cells affects ammonia response (Fig. 3).
Discussion
Ammonia has long been known to act as an olfactory cue for many animals7,8,9,10,11,12,41,42,43. Here we show that it also serves as a taste cue for the fly.
Ecological function of ammonia taste response
Why have flies evolved a taste response to ammonia? The taste response to sugars identifies sources of nutrition. The taste response to bitter compounds signals the presence of a wide diversity of toxic molecules that are produced by plants44. Ammonia, an inorganic compound that is chemically much simpler, is also toxic to animals at high concentrations18,19,20. The concentration of ammonia in fly culture vials may approach 30 mM45, near the threshold of the physiological responses we have found for ammonia. It seems likely that the taste response to ammonia thus warns the fly of the toxicity of food sources with high ammonia levels and inhibits their consumption. In support of this interpretation, we found that high levels of ammonia inhibit consumption in each of two feeding paradigms, one carried out over the course of hours and the other over seconds. We note that the threshold for ammonia detection appears somewhat higher in the CAFÉ assay than in the pharyngeal pumping assay, perhaps because of the presence of sucrose in the CAFÉ assay.
Opposite valences in taste and olfaction
The negative valence of ammonia as a taste cue is in striking contrast to its positive valence as an olfactory cue. Ammonia is aversive in our taste paradigms, yet is attractive in olfactory paradigms for either Drosophila or insect vectors that seek human hosts on which to feed3,7,8,9. Ammonia can thus activate sensory circuits that promote opposite outcomes.
There is precedent for compounds that activate opposing responses via the same sensory system, when presented at different concentrations. For example, low concentrations of the odorant ethyl acetate are attractive in an olfactory paradigm, while high concentrations are repellent46. Likewise, low concentrations of the tastant NaCl are appetitive in taste paradigms, while high concentrations are aversive25.
There is also precedent for compounds that activate both olfactory and gustatory circuits. The insect repellent DEET elicits responses from both systems, but in each case the response is of the same valence47. In Drosophila, CO2 activates the olfactory and gustatory systems with different valences48,49. However, the case of CO2 is distinct from that of ammonia, in that CO2 is aversive to flies when presented as an olfactory stimulus, but is attractive when presented as a gustatory stimulus. In contrast, recent work suggests that like ammonia, the odor of polyamines is attractive, whereas their taste is aversive50.
The ability to sense ammonia via two sensory systems and to activate circuits promoting opposing outcomes may be highly beneficial to insects in their natural environments. Low concentrations of ammonia detected via olfaction may signal the availability of a food source at a distance, whereas high concentrations sensed via gustation warn of toxicity and may signify that local nutrients have been metabolized.
Our work invites a detailed analysis of the taste responses of insect vectors of human disease to ammonia. Human sweat contains on the order of 2 mM–5 mM ammonia5, which is below the thresholds of physiological or behavioral taste response that we have found for Drosophila. It will be interesting to determine whether these levels activate an appetitive taste circuit in insects that are attracted to humans, land on them, and make feeding decisions after landing. We note the possibility of a synergistic effect between ammonia and other sweat compounds in eliciting taste responses; such synergy is observed in eliciting olfactory responses4,7.
The cellular basis of ammonia taste
We found that ammonia is detected by taste sensilla on the labellum, with S sensilla responding strongly and L and I sensilla responding less strongly. Ablation experiments showed that the physiological responses of S sensilla depend on the Gr66a+ bitter-sensing neuron. Likewise, aversion to ammonia in a feeding assay depended on bitter-sensing neurons. These results are consistent with previous findings that these neurons activate an aversive response to bitter compounds. The results suggest that ammonia activates an aversive circuit that is also activated by plant-derived bitter compounds.
L sensilla do not contain a canonical bitter-sensing neuron25,26, so which neuron mediates their ammonia response? The response does not derive from the sugar-sensing neuron, since ablation of this neuron in L sensilla did not reduce the physiological response to ammonia. In S and I sensilla, the bitter-sensitive neuron also responds to high concentrations of salt, an aversive stimulus25. In L sensilla, one of the four neurons is also responsive to high salt25. It seems plausible that the high-salt sensing neuron in L sensilla also senses ammonia, as in S sensilla. However, testing this hypothesis will require a suitable means of manipulating this neuron specifically.
The molecular basis of ammonia detection differs between olfaction and taste
We were surprised to find that the ammonium transporter Amt, which is required for olfactory response to ammonia15, is not required for gustatory response to ammonia, despite its expression in taste sensilla. Also unexpected was the localization of Amt to sugar-sensing neurons of taste sensilla, as opposed to support cells, as in olfactory sensilla.
Previous findings are also consistent with differences in the molecular basis of ammonia detection in olfaction and taste. In the antenna, strong ammonia responses are mediated by the neuronal receptor IR92a, which was not detected in the labellum9,51. Given that other IR family members are expressed in the labellum, another IR family member could mediate ammonia detection in this organ. At least one IR is expressed in some bitter-sensing neurons of the labellum52, and the labellar expression of several other IRs has not yet been examined.
One interpretation of these differences is that the underlying mechanisms of ammonia detection differ between olfaction and taste. In olfaction, low concentrations of airborne ammonia enter the sensillum via openings in the sensillar walls, and are then detected by olfactory neurons expressing IR92a. The basal ammonia concentration of the sensillum lymph is unknown, but the concentration in the larval hemolymph is ~1 mM53, and it is possible that the transport uptake function of Amt is needed to keep the ammonia concentration in the lymph of an olfactory sensillum sufficiently low to allow detection of low levels of airborne ammonia. By contrast, the ammonia levels that activate taste neurons (10–100 mM) and those found in a fly culture vial (~30 mM)45 are much higher than 1 mM. Thus the need to keep the ammonia concentration very low in the lymph of a taste sensillum may be less acute. In any case this unexpected difference invites a detailed investigation of the molecular mechanisms of ammonia taste.
In summary, in this study we have found that ammonia elicits a taste response from Drosophila, and we have characterized this response at the physiological, cellular and behavioral levels. We have identified taste sensilla that detect ammonia and have characterized their electrophysiological responses. We have shown that ammonia is an aversive stimulus in feeding assays, and that both physiological and behavioral responses depend on a class of bitter-sensitive neurons. Our results suggest a means by which flies may avoid ingesting toxic levels of ammonia, but remain attracted to low levels of airborne ammonia. These results lay a foundation for a molecular investigation of the mechanism of ammonia taste.
Materials and Methods
Drosophila stocks
Flies were grown on standard cornmeal-agar medium at 25 °C in a humidity-controlled incubator. The control flies used in Fig. 1 contained a piggyBac transposon, pBacWHf04393, that is not expected to affect ammonia responses. wCS flies were used for behavioral assays, unless otherwise noted. Gr66a-GAL426, Gr64f-Gal427, and UAS-DTA (Bloomington Stock 25039) flies were outcrossed into wCS for 5 generations before electrophysiological and behavioral assays. Amt-GAL4; UAS-mCD8::GFP, Amt1, and the Amt1 control, an isogenized w1118 line, were described previously15. Amt2 was generated as described below and was compared to its genetic control line, wCS. Gr5a-LexA54 was obtained from Kristin Scott. LexAop-m-tdTomato52 and Gr66a-RFP27 were previously generated by the Carlson lab.
Generation of the Amt2 allele
CRISPR-Cas9 genome engineering with homology-directed repair was used to generate the Amt2 null allele. This mutation eliminates ~63% of the Amt coding sequence, leaving 409-nt of coding sequence at the 5′ end and 233-nt at the 3′ end. The remaining sequence contains only the first three of eleven transmembrane regions and is likely to be a null allele.
Guide chiRNA cloning
Gibson Assembly was used to clone pU6-BbsI-chiRNA55,56 plasmids containing each of the chiRNAs, following protocols for Gibson Assembly Master Mix (New England BioLabs, Inc., Ipswich, MA). The reverse primer is complementary to the plasmid itself (chiRNA R), whereas the forward primers also include the 20-nt guide chiRNA sequence with the PAM sequence omitted (Amt 5pchiRNA F and Amt 3pchiRNA F) (Table 1). 20-nt guide chiRNAs were selected with the aid of the CRISPR Optimal Target Finder resource on the flyCRISPR website40. CRISPR targets with 5′ G and NGG PAM sequences were selected for optimal U6-driven transcription and subsequent Cas9 cleavage stringency. Primer sequences complementary to the plasmid sequence are represented as lowercase.
Homology-directed repair template cloning
Homology arms extending 0.98 kb upstream and 1.03 kb downstream of the Cas9 cut site were incorporated into multiple cloning sites of the pHD-DsRed-attP vector40 using amplification primers (Table 2).
To facilitate screening of Amt deletion mutants, this replacement donor plasmid contains removable LoxP-flanked DsRed, which was inserted in the genome at the same locus. A simultaneously inserted attP ΦC31 site facilitates future targeting of this locus.
Embryo injection
y2 cho2 v1; attP40{nos-Cas9}/CyO (CAS-0001, ref. 57) embryos were injected with guide chiRNA and donor plasmids by Bestgene, Inc. (Chino Hills, CA). G0 adults were crossed to w1118 flies. G1 adults expressing DsRed were identified, and were backcrossed to wCS for five generations.
Electrophysiology
Single-sensillum recordings were performed as described in Delventhal et al.58. Only females, aged 6–8 days, were used for electrophysiological recording. To quantify responses, the number of action potentials (spikes) was counted over a 500 ms period, starting 200 ms after contact. A high salt stimulus (400 mM NaCl) was used as a positive control at the beginning and end of the recording session for each sensillum to ensure that at least one GRN was functional. All recordings from sensilla that displayed an average NaCl response of less than 10 spikes/s at the beginning or end of a recording session were discarded (representing less than 10% of overall recordings performed). No more than eight tastants were tested on an individual sensillum of a given fly, with 2–3 minutes between presentations. Details on mean responses, SEM and n for electrophysiology experiments are provided in Supplementary Table 1.
Ammonia solutions were prepared at the indicated concentrations with either NH4Cl or (NH4)2SO4. In Fig. 1A,B, 100 mM NH4Cl and 50 mM (NH4)2SO4 were used to compare equivalent levels of ammonia. All tastants were dissolved in 30 mM tricholine citrate (TCC), an electrolyte that inhibits the water neuron33. Solutions were then brought to a pH of ~7 with NH4OH. Solution aliquots were stored at −20 °C long-term and kept at 4 °C while in use, for no more than a week.
CAFÉ assay
We used a modified version of the CAFÉ assay, similar to that originally described35. The chamber was prepared by filling a 50 ml conical tube with 30 ml of 2% agarose. Two holes were punched into the cap, and shortened 1 ml pipette tips were inserted through the holes partially into the chamber. Calibrated glass capillary tubes (Drummond Scientific Company, Catalog #2–000–001) were filled with liquid food by capillary action and inserted into the chamber through the pipette tips. Two tubes with liquid food were present in each chamber: one with 100 mM sucrose alone and the other with 100 mM sucrose and varying concentrations of NH4Cl.
For the assay, 13 female and 2 male flies (7 days old) were introduced into the CAFÉ chamber, and starved overnight in a 25 °C incubator. Two capillary tubes were introduced the next morning, and flies were given four hours to ingest the liquid food. The amount of solution ingested from each tube was measured, and the preference index (PI) value was calculated according to the formula (X − Y)/(X + Y), where X is the amount ingested from the tube containing sucrose alone and Y is the amount ingested from the tube containing sucrose and ammonia. Average values ± SEM are given.
Pharyngeal pumping assay
We used a pumping assay similar to that described by ref. 36. In brief, we used 7 day old female flies, which were starved 12–14 hours. Each fly was anesthetized and placed in a 1000-μl pipette tip. A second 1000-μl pipette tip was inserted into the first tip, thereby containing the fly for the subsequent starvation period. Flies were then kept in a 100 mm Petri dish with 3 Kimwipes wetted with 5 ml water to prevent dehydration.
After starvation, gentle aspiration was used to immobilize a fly for food presentation, as follows: a pulse of air was applied to the broad opening of the inserted 1000-μl pipette tip, such that the fly was pushed into the narrow opening of the second 1000-μl pipette tip, thereby immobilizing the fly with its head and proboscis exposed. Occasionally, ends of the 1000-μl pipette tips had to be trimmed with a razor blade, to widen the tip so as to accommodate the fly’s head. The prepared fly was then mounted on a micromanipulator for analysis under a Nikon SMZ800 stereomicroscope. Flies were filmed with a Sony HD Camcorder that was attached to the microscope through an adapter.
A drop of liquid medium was delivered from a P20 PipetteMan mounted on a micromanipulator, which allowed for fine adjustments during the delivery process. The liquid drop was carefully presented to the labellum, such that liquid only contacted the sensory hairs and cuticle present on the labellum. The fly was presented liquid medium for 2 seconds, during which it had the chance to respond by extending its proboscis to drink. Flies that began pharyngeal pumping to ingest the solution were allowed to continue until they ended the bout by breaking contact with the liquid drop. After breaking contact with the solution, flies were given 3 seconds rest before being presented the liquid solution once again. They were then given 2 seconds to initiate a second bout. This process was repeated until the fly no longer responded to presented media, after which the experiment was terminated. Typically, 2 seconds was a sufficient presentation time, as longer presentation times did not appreciably increase the likelihood that a fly would initiate a subsequent feeding bout. On average flies engaged in 1–3 bouts.
For flies that did not respond during the initial 2 second presentation, the liquid solution was removed and the fly was given 3 seconds rest before being presented the liquid solution once again. This presentation process was repeated four times. If flies did not initiate a feeding bout after the fourth presentation, they were considered non-responders and discarded. Approximately half of all flies were non-responders in the case of each genotype.
Flies were offered solutions containing varying concentrations of NH4Cl. Each fly was used for only one experiment to prevent previous experience from influencing its response decisions. The video was analyzed at a later time, during which the total time spent ingesting the solution was measured using QuickTime Media Player. When different compounds were tested, the analysis was conducted blind to the stimulus. When different genotypes were tested, the analysis was carried out blind to genotype. Average values ± SEM are indicated for responders.
Whole-mount imaging
Labella and legs were dissected from male and female flies (7–21 days) on a Sylgard plate. Tissues were immersed in a solution of 0.5X PBS, 0.1% Tween-20, and 50% glycerol for at least five minutes. Tissue was mounted on glass slides and imaged within one day. Confocal stacks were acquired on a Zeiss LSM510 confocal microscope and processed using NIH ImageJ (version 1.44o).
Generation of αAmt antibody
The last 148 amino acids of the Amt cytosolic C-terminal tail were expressed with a 6xHis-tag at the N-terminus in E. coli and affinity-purified by Novogene. Polyclonal antibodies against this protein antigen were generated in a guinea pig through standard protocols (Cocalico Biologicals, Inc.). Serum was used for immunofluorescence experiments at a 1:200 dilution.
Immunofluorescence labeling of labellar sections
7 day old flies were anesthetized, placed in a collar, covered with OCT (Tissue-Tek), and frozen on dry ice. 40 μm labellar sections were collected on slides, and stored at −80 °C up to one week. Slides were briefly thawed before a 10 minute fixation in 4% formaldehyde in PBS. Sections were washed 3 × 5 minutes in PBS, permeablized for 30 min in PBST (PBS plus 0.1% Tween-20) with 0.2% Triton X-100, blocked for 30 minutes in 1% BSA-PBST, then incubated overnight at 4 °C with primary antibodies diluted in 1% BSA-PBST. Anti-Amt was used at 1:200 and mouse anti-GFP (Roche) was used at 1:500. The following day, sections were washed 3 × 10 minutes in PBST, then incubated for 2 hours with Alexa Fluor secondary antibodies diluted in 1% BSA-PBST. Sections were washed 3 × 5 minutes in PBST and mounted in Vectashield. All microscopy was performed using a Zeiss LSM 510 Laser Scanning Confocal Microscope, and images were processed with ImageJ software.
Additional Information
How to cite this article: Delventhal, R. et al. The taste response to ammonia in Drosophila. Sci. Rep. 7, 43754; doi: 10.1038/srep43754 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Miller, S. L. A production of amino acids under possible primitive earth conditions. Science 117, 528–529 (1953).
Public Health Statement – Ammonia, CAS#: 7664-41-7. Department of Health and Human Services, Public Health Service Agency for Toxic Substances and Disease Registry (ATSDR) (2004).
Braks, M. A. H., Meijerink, J. & Takken, W. The response of the malaria mosquito, Anopheles gambiae, to two components of human sweat, ammonia and L-lactic acid, in an olfactometer. Physiol Entomol 26, 142–148, doi: 10.1046/J.1365-3032.2001.00227.X (2001).
Renate, C. S., Yu Tong, Q., Joop, J. A. v. L. & Willem, T. Synergism between ammonia, lactic acid and carboxylic acids as kairomones in the host-seeking behaviour of the malaria mosquito Anopheles gambiae sensu stricto (Diptera: Culicidae). Chem Senses 30, 145–152, doi: 10.1093/chemse/bji010 (2005).
Brusilow, S. W. & Gordes, E. H. Ammonia secretion in sweat. Am J Physiol. 214, 513–517 (1968).
Wolfgang, R. M. et al. A novel synthetic odorant blend for trapping of malaria and other african mosquito species. Journal of Chemical Ecology 38, 235–244, doi: 10.1007/s10886-012-0088-8 (2012).
Geier, M., Bosch, O. J. & Boeckh, J. Ammonia as an attractive component of host odour for the yellow fever mosquito, Aedes aegypti. Chem Senses 24, 647–653, doi: 10.1093/Chemse/24.6.647 (1999).
Taneja, J. & Guerin, P. M. Ammonia attracts the haematophagous bug Triatoma infestans: behavioural and neurophysiological data on nymphs. Journal of Comparative Physiology A 181, 21–34, doi: 10.1007/S003590050089 (1997).
Min, S. H., Ai, M. R., Shin, S. A. & Suh, G. S. B. Dedicated olfactory neurons mediating attraction behavior to ammonia and amines in Drosophila . PNAS 110, E1321–E1329, doi: 10.1073/pnas.1215680110 (2013).
Bateman, M. & Morton, T. The importance of ammonia in proteinaceous attractants for fruit flies (Family: Tephritidae). Crop and Pasture Science 32, 883–903 (1981).
Robacker, D. & Flath, R. Attractants fromStaphylococcus aureus cultures for Mexican fruit fly, Anastrepha ludens. Journal of chemical ecology 21, 1861–1874 (1995).
Richardson, C. H. The Response of the House-fly (Musca Domestica L.) to Ammonia and Other Substances. (New Jersey Agricultural Experiment Stations, 1916).
Haggart, D. A. & Davis, E. E. Ammonia-sensitive neurones on the first tarsi of the tick, Rhipicephalus sanguineus. Journal of Insect Physiology 26, 517–523 (1980).
Meijerink, J., Braks, M. A. H. & Loon, J. J. A. V. Olfactory receptors on the antennae of the malaria mosquito Anopheles gambiae are sensitive to ammonia and other sweat-borne components. Journal of Insect Physiology 47, 455–464, doi: 10.1016/S0022-1910(00)00136-0 (2001).
Menuz, K., Larter, N. K., Park, J. & Carlson, J. R. An RNA-seq screen of the Drosophila antenna identifies a transporter necessary for ammonia detection. PLoS Genetics 10, e1004810, doi: 10.1371/journal.pgen.1004810 (2014).
Tauxe, G., MacWilliam, D., Boyle, S., Guda, T. & Ray, A. Targeting a dual detector of skin and CO2 to modify mosquito host seeking. Cell 155, doi: 10.1016/j.cell.2013.11.013 (2013).
Syed, Z. & Leal, W. S. Acute olfactory response of Culex mosquitoes to a human- and bird-derived attractant. PNAS 106, 18803–18808, doi: 10.1073/pnas.0906932106 (2009).
Weihrauch, D., Donini, A. & O’Donnell, M. J. Ammonia transport by terrestrial and aquatic insects. Journal of insect physiology 58, 473–487 (2012).
Ip, Y., Chew, S. & Randall, D. Ammonia toxicity, tolerance, and excretion. Fish physiology 20, 109–148 (2001).
Randall, D. & Tsui, T. Ammonia toxicity in fish. Marine pollution bulletin 45, 17–23 (2002).
Bachmanov, A. A., Beauchamp, G. K. & Tordoff, M. G. Voluntary consumption of NaCl, KCl, CaCl2, and NH4Cl solutions by 28 mouse strains. Behavior genetics 32, 445–457 (2002).
van der Klaauw, N. J. & Smith, D. V. Taste quality profiles for fifteen organic and inorganic salts. Physiology & behavior 58, 295–306 (1995).
Schiffman, S. S. & Erickson, R. P. A psychophysical model for gustatory quality. Physiology & behavior 7, 617IN11623–11622IN12633 (1971).
Stocker, R. The organization of the chemosensory system in Drosophila melanogaster: a review. Cell & Tissue Research 275, 3–26 (1994).
Hiroi, M., Meunier, N., Marion-Poll, F. & Tanimura, T. Two antagonistic gustatory receptor neurons responding to sweet-salty and bitter taste in Drosophila . Journal of neurobiology 61, 333–342, doi: 10.1002/neu.20063 (2004).
Weiss, L. A., Dahanukar, A., Kwon, J. Y., Banerjee, D. & Carlson, J. R. The molecular and cellular basis of bitter taste In Drosophila. Neuron 69, 258–272, doi: 10.1016/j.neuron.2011.01.001 (2011).
Dahanukar, A., Lei, Y. T., Kwon, J. Y. & Carlson, J. R. Two Gr genes underlie sugar reception In Drosophila. Neuron 56, 503–516, doi: 10.1016/j.neuron.2007.10.024 (2007).
Ling, F., Dahanukar, A., Weiss, L., Kwon, J. & Carlson, J. The molecular and cellular basis of taste coding in the legs of Drosophila. J Neurosci 34, 7148–7164, doi: 10.1523/JNEUROSCI.0649-14.2014 (2014).
Raad, H., Ferveur, J. F., Ledger, N., Capovilla, M. & Robichon, A. Functional Gustatory Role of Chemoreceptors in Drosophila Wings. Cell Rep 15, 1442–1454, doi: 10.1016/j.celrep.2016.04.040 (2016).
Yanagawa, A., Guigue, A. M. & Marion-Poll, F. Hygienic grooming is induced by contact chemicals in Drosophila melanogaster. Front Behav Neurosci 8, 254, doi: D - NLM: PMC4107972 OTO - NOTNLM (2014).
Whole Food Catalog. wholefoodcatalog.info/nutrient/ammonia/foods/high/(Accessed 19 October 2016).
Ammonia Management from Manure Storage & Spreading. Nova Scotia Agricultural College (2007).
Wieczorek, H. & Wolff, G. The labellar sugar receptor of Drosophila . J Comp Physiol A Neuroethol Sens Neural Behav Physiol 164, 825–834 (1989).
Mazor, M., Gothilf, S. & Galun, R. The role of ammonia in the attraction of females of the Mediterranean fruit fly to protein hydrolysate baits. Entomologia Experimentalis et Applicata 43, 25–29 (1987).
Ja, W. W. et al. Prandiology of Drosophila and the CAFE assay. Proc Natl Acad Sci USA 104, 8253–8256, doi: 10.1073/pnas.0702726104 (2007).
Manzo, A., Silies, M., Gohl, D. M. & Scott, K. Motor neurons controlling fluid ingestion in Drosophila . Proc Natl Acad Sci USA 109, 6307–6312, doi: 10.1073/pnas.1120305109 (2012).
Marella, S. et al. Imaging taste responses in the fly brain reveals a functional map of taste category and behavior. Neuron 49, 285–295, doi: 10.1016/j.neuron.2005.11.037 (2006).
Thorne, N., Chromey, C., Bray, S. & Amrein, H. Taste perception and coding in Drosophila . Curr Biol 14, 1065–1079, doi: 10.1016/j.cub.2004.05.019 (2004).
Jiao, Y., Moon, S. J., Wang, X., Ren, Q. & Montell, C. Gr64f is required in combination with other gustatory receptors for sugar detection in Drosophila . Curr Biol 18, 1797–1801, doi: 10.1016/j.cub.2008.10.009 (2008).
Gratz, S. J. et al. Highly specific and efficient CRISPR/Cas9-catalyzed homology-directed repair in Drosophila . Genetics 196, 961–971, doi: 10.1534/genetics.113.160713 (2014).
Nevitt, G. A., Bergstrom, D. M. & Bonadonna, F. The potential role of ammonia as a signal molecule for procellarifform seabirds. Marine Ecology-Progress Series 315 (2006).
Strand, S. & Hamner, W. Schooling behavior of Antarctic krill (Euphausia superba) in laboratory aquaria: reactions to chemical and visual stimuli. Marine Biology 106, 355–359 (1990).
Frøkjær-Jensen, C., Ailion, M. & Lockery, S. R. Ammonium-acetate is sensed by gustatory and olfactory neurons in Caenorhabditis elegans. PLoS One 3, e2467 (2008).
Smith, C. Biology of sensory systems. (John Wiley & Sons, 2008).
Borash, D. J., Gibbs, A. G., Joshi, A. & Mueller, L. D. A genetic polymorphism maintained by natural selection in a temporally varying environment. Am Nat 151, 148–156, doi: Doi 10.1086/286108 (1998).
Rodrigues, V. & Siddiqi, O. Genetic analysis of chemosensory pathway. Proc. Indian Acad. Sci. 87 B, 147–160 (1978).
DeGennaro, M. The mysterious multi-modal repellency of DEET. Fly 9, 45–51 (2015).
Fischler, W., Kong, P., Marella, S. & Scott, K. The detection of carbonation by the Drosophila gustatory system. Nature 448, 1054–1057, doi: 10.1038/nature06101 (2007).
Suh, G. S. et al. A single population of olfactory sensory neurons mediates an innate avoidance behaviour in Drosophila . Nature 431, 854–859, doi: 10.1038/nature02980 (2004).
Hussain, A. et al. Ionotropic chemosensory receptors mediate the taste and smell of polyamines. PLoS Biol 14, e1002454 (2016).
Benton, R., Vannice, K. S., Gomez-Diaz, C. & Vosshall, L. B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila . Cell 136, 149–162 (2009).
Koh, T. W. et al. The Drosophila IR20a clade of ionotropic receptors are candidate taste and pheromone receptors. Neuron 83, 850–865, doi: 10.1016/j.neuron.2014.07.012 (2014).
Browne, A. & O’Donnell, M. J. Ammonium secretion by Malpighian tubules of Drosophila melanogaster: application of a novel ammonium-selective microelectrode. J Exp Biol 216, 3818–3827, doi: 10.1242/jeb.091082 (2013).
Gordon, M. D. & Scott, K. Motor control in a Drosophila taste circuit. Neuron 61, 373–384, doi: 10.1016/j.neuron.2008.12.033 (2009).
Barnes, W. M. PCR amplification of up to 35-kb DNA with high fidelity and high yield from lambda bacteriophage templates. Proc Natl Acad Sci USA 91, 2216–2220 (1994).
Gratz, S. J., Cummings, A. M., Nguyen, J. N. & Hamm, D. C. Genome engineering of Drosophila with the CRISPR RNA-guided Cas9 nuclease. Genetics 194, 1029–1035 (2013).
Kondo, S. & Ueda, R. Highly improved gene targeting by germline-specific Cas9 expression in Drosophila . Genetics 195, 715–721, doi: 10.1534/genetics.113.156737 (2013).
Delventhal, R., Kiely, A. & Carlson, J. Electrophysiological recording from Drosophila labellar taste sensilla. J Vis Exp. e51355, doi: 10.3791/51355 (2014).
Acknowledgements
We thank Dr. Kristin Scott and the Bloomington Drosophila Stock Center for fly lines. pDsRed-attP was a gift from Melissa Harrison & Kate O’Connor-Giles & Jill Wildonger (Addgene plasmid # 51019, ref. 40 (Gratz et al. 2014)). pU6-BbsI-chiRNA was a gift from Melissa Harrison & Kate O’Connor-Giles & Jill Wildonger (Addgene plasmid # 4594658). K.M. was supported by an NIH NRSA (F32DC011242). R.D. was supported by an NIH NRSA (5F31DC012985). R.M.J. was supported by an NIH NRSA (5F32DC013507). JSS acknowledges financial support from the NSF Graduate Research Fellowship Program, NIH Institute of Health (T32 GM007499), and the Dwight N. and Noyes D. Clark Scholarship Fund. The project was supported by NIH grants to JRC.
Author information
Authors and Affiliations
Contributions
J.R.C., R.D. and K.M. wrote the manuscript text. R.D. and K.M. prepared figures. R.D., K.M., R.M.J. and J.P. performed experiments and data analysis. K.M. and J.S.S. created tools. All authors reviewed manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Delventhal, R., Menuz, K., Joseph, R. et al. The taste response to ammonia in Drosophila. Sci Rep 7, 43754 (2017). https://doi.org/10.1038/srep43754
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep43754
This article is cited by
-
Necrophagy by insects in Oculudentavis and other lizard body fossils preserved in Cretaceous amber
Scientific Reports (2023)
-
The proton channel OTOP1 is a sensor for the taste of ammonium chloride
Nature Communications (2023)
-
Ionotropic receptors mediate nitrogenous waste avoidance in Drosophila melanogaster
Communications Biology (2021)
-
Identification and characterization of CYPs induced in the Drosophila antenna by exposure to a plant odorant
Scientific Reports (2021)
-
Recent advances in the genetic basis of taste detection in Drosophila
Cellular and Molecular Life Sciences (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
