
Abstract
Obtaining reliable estimates of the structure of carnivore communities is of paramount importance because of their ecological roles, ecosystem services and impact on biodiversity conservation, but they are still scarce. This information is key for carnivore management: to build support for and acceptance of management decisions and policies it is crucial that those decisions are based on robust and high quality information. Here, we combined camera and live-trapping surveys, as well as telemetry data, with spatially-explicit Bayesian models to show the usefulness of an integrated multi-method and multi-model approach to monitor carnivore community structures. Our methods account for imperfect detection and effectively deal with species with non-recognizable individuals. In our Mediterranean study system, the terrestrial carnivore community was dominated by red foxes (0.410 individuals/km2); Egyptian mongooses, feral cats and stone martens were similarly abundant (0.252, 0.249 and 0.240 individuals/km2, respectively), whereas badgers and common genets were the least common (0.130 and 0.087 individuals/km2, respectively). The precision of density estimates improved by incorporating multiple covariates, device operation, and accounting for the removal of individuals. The approach presented here has substantial implications for decision-making since it allows, for instance, the evaluation, in a standard and comparable way, of community responses to interventions.
Similar content being viewed by others
Introduction
Management goals commonly rely on information about the abundance of species. But monitoring is still one of the most controversial issues when managing wildlife1,2. Monitoring has traditionally followed a species-specific approach; although in recent times, management interventions oriented to ecosystem functioning perspectives demands reliable estimates of community-level structures, including density estimates of entire guilds3. Whereas obtaining reliable density estimates for some group of species, for instance, farmland birds, large herbivores in open landscapes, or soil invertebrates, seems achievable, estimates for elusive and cryptic vertebrate guilds, such as mammalian carnivores, commonly showing heterogeneous capture probabilities, remains a challenge.
The importance of obtaining reliable estimates of community structures for mammalian carnivores, including domestic species (dogs Canis familiaris and cats Felis catus), is of paramount importance to inform decision-making processes. Carnivores can be important drivers of ecosystem function, structure or dynamics. For example, this guild participates in different top-down ecosystem processes, such as trophic cascades4,5, or providing different ecosystem services, such as seed dispersal services6,7. Moreover, in the case of feral carnivores, this guild can pose particular problems for biodiversity conservation8,9.
Multiple tools have been developed over the last few decades to monitor mammalian carnivores10,11. But estimates on carnivore community structures (here we refer to the number of species present and their abundance) are still very rare12. Although different proxies (indices) for population abundance/density have been widely used (e.g., from sign counts and observations to number of captures in camera trapping surveys)11,13,14, they have also been criticised because they often do not account for variability and biases in detection probabilities among individuals15,16. Moreover, the use of non-spatially explicit analytical procedures has also been questioned because of overestimation problems in density estimates17. This issue has been recently overcome with the development of spatially explicit approaches15. However, we lack an integration of different methodological and analytical protocols to deal with different species-specific particularities yet.
Over the last decade, camera surveys have become the dominant tool to survey communities of rare and cryptic terrestrial mammals3,11,18,19. If monitoring surveys are well designed, they can yield standard and comparable data on distribution, abundance, behaviour, and community composition20,21. However, little attention has been given to the use of camera surveys to characterise community structures3,19. Within carnivore communities, although some species can be easily identified like most felid species22,23, the use of camera surveys to estimate community structures has been constrained by the fact that some individuals and species can not be identified. Moreover, only recently have detection probabilities been accounted for (in the absence of individual marks, detection rates confound abundance and detectability)24,25.
Nevertheless, the identification requirement has also been overwhelmed in recent times. State of the art analytical inference-based procedures to estimate densities using spatially explicit frameworks - linking abundance with location by estimating a latent variable representing the individual’s activity centers26,27, such as spatially explicit capture-recapture (SCR), mark-resight (SMR) and spatial count (SC) models, can be implemented even with species without marks (SMR, SC)15,24,26, common in several carnivore species. Interestingly, they can be combined with different sampling methods to estimate community structures. In this regard, the combination of simple camera surveys–standardisable and repeatable, and with low requirements and costs–with an integration of different spatially explicit analytical approaches (SCR, SMR, SC)28 –emerges therefore as a promising procedure to estimate densities for entire communities or guilds. Precision in the spatially explicit estimates, can be improved by integrating additional information, such as telemetry data from some individuals in order to improve the estimates of the movement parameters15,25, or using informative priors for sigma24,29,30 (i.e., the Gaussian scale parameter that determines the rate of decrease in detection probability between activity centers and traps).
In this study, we combined camera and live-trapping surveys with spatially explicit Bayesian models (SCR, SMR and SC), and additional telemetry data, to propose an integrated approach to monitor carnivore community structures, including those species where individuals can not be identified. Depending on the characteristics of each species and the number of capture events, we used SCR if all the animals were recognizable, SMR if some individuals were recognisable (including artificial and/or natural marks), and SC for unmarked animals. The procedure proposed here would allow for reliable and comparable density estimates for entire carnivore communities.
Results
We found 9 different carnivore species in the study area (Table 1), captured 21 animals from 5 species with traps and radio-tagged (VHF) 6 individuals. Considering the encounter histories for each species, we were able to estimate the densities of red fox (Vulpes vulpes), stone marten (Martes foina), European badger (Meles meles), Egyptian mongoose (Herpestes ichneumon), common genet (Genetta genetta) and feral cat (Felis catus). From the list of species expected to be found in this area, we did not detect the presence of wildcat (Felis sylvestris). Details on encounter frequencies for naturally and artificially marked animals as well as non-recognisable individuals for each species are shown in Table 1.
The SCR approach was only used with feral cats (55 photographic events, six of which were discarded because of very poor quality images, and 11 captures with extraction), the SMR approach was used for red fox, stone martens, badgers and common genets (a total of 12 individuals trapped and marked on primary occasions or recognised, from which 8 were re-sighted on secondary occasions; and 143 capture events on secondary occasions: 48 recapture events from marked/recognisable individuals and 85 unmarked individuals), whereas the SC was used for Egyptian mongooses (77 events of unmarked individuals). We assumed that marked individuals were a random sample from the resulting state–space because marking (live-trapping) took place across the extent of the re-sighting array (cameras) (Fig. 1). For SMR models, we used different numbers of secondary occasions (Table 1) in order to maximise the number of animals marked and/or recognisable and the number of events on secondary occasions.

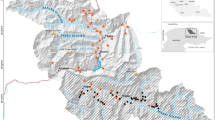
Mean camera spacing was 483 m. The figure was produced by José Jiménez using ArcGIS 10 (Esri Inc., Redlands, CA, USA).
The model including trap-specific covariate, behavior and sampling occasions was selected for red fox (Table 2). The model with a trap-specific covariate was selected for stone martens (Table 2). The null model -with no detection probability varying between traps- was the best candidate model for feral cats (Table 2). For stone martens and feral cats, for which known removals occurred during the study period (a monitored stone marten was found dead, whereas eleven feral cats were removed from the field; Table 1), we applied the extension to account for removal of individuals in the model. However, because of the extraction of individuals we did not use covariates from individuals (behaviour) or time to adjust the baseline encounter rate. Finally, for badgers and common genets the samples of populations were too small to adjust for covariates. For Egyptian mongoose (SC) we use a trap-specific covariate (Table 2).
The carnivore community in the study area was dominated by red fox (density >0.4 individuals/km2), followed by Egyptian mongooses, feral cats, and stone martens (densities between 0.2 and 0.3 individuals/km2), and finally, badgers and common genets (densities <0.2 individuals/km2) (Fig. 2). The most abundant species were red fox, with 0.410 individuals/km2 (95% BCI = 0.208–0.724; CV = 0.32; Table S1). The density estimate  for Egyptian mongooses was 0.252 individuals/km2 (95% BCI = 0.125–0.450; CV = 0.33 Table S2), for feral cats 0.249 individuals/km2 (95% BCI = 0.150–0.376; CV = 0.24; Table S3), and 0.240 individuals/km2 (95% BCI = 0.114–0.442; CV = 0.35; Table S4) for stone martens. Finally, the density estimates
for Egyptian mongooses was 0.252 individuals/km2 (95% BCI = 0.125–0.450; CV = 0.33 Table S2), for feral cats 0.249 individuals/km2 (95% BCI = 0.150–0.376; CV = 0.24; Table S3), and 0.240 individuals/km2 (95% BCI = 0.114–0.442; CV = 0.35; Table S4) for stone martens. Finally, the density estimates  for the less abundant species were 0.130 individuals/km2 (95% BCI = 0.036–0.339; CV = 0.59; Table S5) for badgers and 0.087 individuals/km2 (95% BCI = 0.024–0.229; CV = 0.62; Table S6) for common genets.
for the less abundant species were 0.130 individuals/km2 (95% BCI = 0.036–0.339; CV = 0.59; Table S5) for badgers and 0.087 individuals/km2 (95% BCI = 0.024–0.229; CV = 0.62; Table S6) for common genets.
 of the six carnivores analysed in this study.
of the six carnivores analysed in this study.
Polygons are shaped in proportion to the posterior probability density.
Discussion
Capture-recapture techniques are generally considered the “gold standard” for generating population estimates. But, in carnivore communities, the number of species and recognisable individuals are usually small. However, new analytical spatially explicit approaches allow for the estimation of densities of unmarked populations15,24,26, facilitating the study of carnivore community structures.
On the other hand, despite the fact that camera trapping has become popular in wildlife monitoring28, little attention has been paid to the use of camera surveys to characterise community structures3,12,19. However, this approach has been used in multispecies occupancy surveys, species richness inventories18,19 and estimates of relative abundance (indices) of species3,31, as the number of capture events has been correlated with species abundance11. Nonetheless, the use of indices wrongly assumes that detection probabilities are constant15. The combination of camera surveys, together with live-trapping (to artificially mark some animals in unrecognisable species, and a small number of individuals, e.g., 1 or 2 animals per species, to gather spatial information to be integrated into the models) and different spatially explicit analytical procedures (SCR, SMR and SC)24,26 facilitates the estimate of carnivore community structures. We recommend the use of SCR or SMR whenever possible because SC parameter estimates–when no individual information is available - are inherently less precise15.
We extended the use of trap operation in all models, because traps were not continuously operational during each sample occasion (Fig. 3). In addition, we accounted for removal of individuals (in our case, stone martens and feral cats). The use of covariates allowed us to identify an influence of the behavioural response in foxes to previous detection. For mongoose, red fox and stone marten there are differences between the probability of detection in traps and cameras, but not for feral cats. The numbers of captures also suggest that there are no differences for common genets. Finally, less logistical effort and lower costs are associated with the methods used here to estimate densities compared to other standard approaches. In the case of unmarked species, SMR only requires the physical capture of a few individuals, although the precision increased substantially with the proportion of marked individuals. In the case of SC, on the other hand, telemetry data could be used as data or as an informative prior for sigma15,24,26.

The 98 sampling days are grouped into 14 7-day blocks (sampling occasions). Within each occasion, for each survey device we show operation (the number of days within each occasion where the device was operative) in a colour scale (right, 0 = the device was not operative on any day, 1 = the device was operative on all days).
By combining spatially explicit models, we were able to estimate the structure of a close-to-complete terrestrial carnivore community in the southern Iberian Peninsula (Table 1). It is worth noting that SCR, SMR and SC density estimates are not totally comparable to non-spatially explicit procedures since the latter overestimate density estimates17.
With the previous caution stated, our carnivore community was dominated by red fox (Fig. 2). However, our density estimate for red fox (0.410 ± 0.133 individuals/km2) can be considered within the mean density values at the level of the Iberian Peninsula32,33, similar to the 0.54–0.69 red fox/km2 estimated using non-spatial procedures33. For mongooses, our density estimate (0.252 ± 0.082 individuals/km2) was below the estimate reported in the Doñana area, SW Spain (1.2 individuals/km2)34. The density estimates for feral cats (0.249 ± 0.059 individuals/km2) is the first spatially explicit density estimate for this non-native species in Spain. Additionally, to our knowledge, our density estimate for stone martens is one of the first estimates available in Spain (0.240 ± 0.083 individuals/km2). The density estimate for badgers was lower than those provided using telemetry and sign count data (0.23–0.67 individuals/km2 and 0.36–0.48 individuals/km2, respectively)35,36. Non-spatial capture-recapture and SCR methods have been used with common genets, showing density estimates ranging from 0.58 to 1.12 individuals/km233,37,38 and from 0.16 to 0.79 individuals/km2, respectively39. Our density estimate (0.087 ± 0.054 individuals/km2) is below the estimate provided by Sarmento & Cruz (2014)39.
The approach used in this study additionally allowed to obtain spatial characteristics of the carnivore community, such as a comparison of the use of space among species, as the output is a set of locations of the activity centers (Fig. 4). Therefore, this approach can contribute to substantially increasing our understanding of spatial inter-specific interactions in carnivore guilds40,41.

Locations of the activity centers for the six carnivores analysed in this study.
Coefficients of variation (CV) are dependent on the models and data used. Thus, high coefficients of variation in density estimates were found in badgers (CV = 0.59) and common genets (CV = 0.62), which were those species with fewer events (8 capture events on secondary occasions for both species, and 1 and 3 marked/recognisable individuals, respectively). However, it is worth noting that even in these situations the VHF collars allowed for estimates of scale (movement) parameters15 and thus, their use in the model. For SC model (Egyptian mongoose with 77 events) CV was 0.33. For those species where we used SMR approaches, there were 46 and 81 capture events on secondary occasions, and 4 marked/recognisable individuals (stone marten and red fox) resulting in CV of 0.35 to 0.32, respectively. For the feral cat model using a SCR model (55 events) we found the lowest CV = 0.24.
Reliable estimates of the abundance of species and community structures are essential to inform, support and accept decision-making management processes. The management and conservation of mammalian carnivores is controversial because of the multiple socio-economic and conservation interests involved. Information about their population status or the impact of management interventions is constantly demanded not only by managers, researchers and conservationists, but also by the general public. Consequently, the quality of the data and robustness of science behind the data are crucial not only to follow an adaptive management framework, but also to build support and acceptance for management decisions and policies. Otherwise, incorrect density estimates (e.g., inflated numbers) could lead to misinterpretations of the impact of management interventions, undesirable steps in the decision-making process or can even place species at risk42. The rise of spatially explicit modelling approaches (SCR, SMR, SC), however, facilitates the estimation of densities and the uncertainty around estimates, for populations and communities, including those species where individuals can not be identified. We believe that integrated approaches, as the one presented here, will therefore be very useful for gathering information on community structures, and evaluating, in a standard and comparable way, changes in carnivore communities when management and conservation interventions are implemented.
Methods
Study area
This study was carried out in the area of Valdecigüeñas, Badajoz, SE Spain (Fig. 1), covering ca. 10,000 ha. The landscape is dominated by Mediterranean woodland, pastures and dehesas of holm oak (Quercus ilex). The area occupied by Mediterranean scrubland is residual. The Viar River crosses the study area NW-SE (Fig. 1). The main human land use in the area is sheep farming. No apex predators were present during the study period, therefore we focused on estimating the carnivore community. The main prey for carnivores in this area are European rabbits (Oryctolagus cunniculus), with densities ranging between 0.5 and 1 rabbits/ha (rabbit density estimates based on pellet counts along transects)43. Previous to this study, but not during the sampling period, fox and Egyptian mongoose populations were controlled.
Preliminary considerations for survey design
To estimate the structure of the carnivore community, data collection was conditioned by the analytical procedures used: SCR, SMR and SC26. The three methods allow for the estimation of individual’s activity centers within the prescribed state space (S). To do this, on the one hand, SCR relies on identifying all individuals of the population captured in the survey devices (e.g., camera-traps). On the other hand, in SMR a sample of individuals must be naturally marked or captured and tagged (or otherwise artificially marked) on the primary occasion. This must be followed by re-sighting surveys (secondary occasions) combining information from both the marked and unmarked fractions of the population. For the unmarked population, we used traps/cameras and occasion data as reduced information of “latent” encounter histories of individuals. Higher percentages of recognisable individuals translates into more accurate and precise parameter estimates outcomes26. For SC (Spatial Counts), we used only the latent encounter histories. Although the analytical method that should be prioritized is SCR, a priori, it is difficult to predict which model will be the most appropriate given the available data, except for the fact that SCR cannot be applied to species where individuals are not recognisable. In the case of SMR and SC models, integrating data from telemetry can improve parameter estimates15.
Data collection
Between January 15, 2013 and April 23, 2013 (98 days) we collected data on the different carnivores occurring in the study area. A total of 66 cameras (models Ltl Acorn© 5210, ScoutGuard© SG560-8M and ScoutGuard© SG570-6M) were homogeneously deployed in an area of ca. 2,300 ha (Fig. 1). Cameras were placed at a height of 30–60 cm, operated 24 h/day, and were configured with a trigger delay of 1 s. Urine from Iberian lynx (Lynx pardinus) was used as attractant. In Mediterranean Spain, lynx urine has been proved as one of the most effective and generalist attractants of mesocarnivores44. At each camera station, we impregnated a piece of cork with lynx urine and placed it on the top of a metal rod at a height of 30–60 cm set at a distance between 2.5 and 4.5 m from the camera. Cameras were checked once per month. In addition, considering the grid of cameras, we additionally placed 69 homogeneously distributed live-traps in the study area (Fig. 1). Carnivores were live-trapped using several methods: self-made box-traps (n = 14), Tomahawk© box-traps (n = 33), and Collarum® (n = 22). Box-traps were baited with live-prey (pigeons provided with food and water) and the Collarum with COLLARUM® Canine Bait. Traps were visually checked daily early in the morning and through automatic alerts using GPS-GPRS transmitters. Sampling devices (69 traps and 66 cameras) were operative for a total (primary and secondary occasions pooled) of 1,391 and 5,395 days for traps and cameras, respectively (Fig. 3).
To set the distance between camera traps, for SCR, Sun, Fuller & Royle45 recommends a distance between cameras of less than 2σ, where σ is the scale or movement parameter for the target species. Chandler and Royle (2013)26 for SMR suggests that the distance between cameras should be enough to ensure that a given individual can potentially be captured in several traps, coercing the spatial correlation among captures. Here, as we were interested in the entire carnivore community, we used an estimate of the scale parameter  46 considering a species showing small spatial requirements. Thus, we considered acceptable a distance d between camera traps ranging between
46 considering a species showing small spatial requirements. Thus, we considered acceptable a distance d between camera traps ranging between  and
and  , where
, where  was calculated as follows:
was calculated as follows:

where q2,α was the value of a Chi-square with 2 degrees of freedom (α = 0.05, q2,α = 5.99) and S was the home range of the species (m2)26,46.
Based on available information on the spatial ecology of these species, we considered that a priori the species showing the smallest home range in our case was the Egyptian mongoose with ca. 300 ha47 and therefore  was set at 400 m. By default, we decided to use a distance between cameras of 500 m to establish the grid of camera traps. After fieldwork, the average distance between cameras was 483 m. We used a sampling area (2,300 ha) larger than the largest home range of the species expected to be present in this area and with the greatest spatial requirements: the home range for wildcats (Felis silvestris) (1,375 ha)48.
was set at 400 m. By default, we decided to use a distance between cameras of 500 m to establish the grid of camera traps. After fieldwork, the average distance between cameras was 483 m. We used a sampling area (2,300 ha) larger than the largest home range of the species expected to be present in this area and with the greatest spatial requirements: the home range for wildcats (Felis silvestris) (1,375 ha)48.
Additionally, we captured and marked a sample of individuals from each species within the study area to facilitate recognition during subsequent resighting events, a pre-requisite for SMR models (Table 1). Captured animals were marked with numbered plastic collars and photographed to facilitate their identification with the cameras. Some animals, 1–2 individuals per species, were also VHF radio-tagged (Ayama©). All captured animals were immobilised by intramuscular injection of medetomidine (Domitor®, Merial, Lyon, France) combined with ketamine (Imalgene®, Merial, Lyon, France). VHF collared individuals were located 2 to 3 times per week.
Data analyses
From the camera survey data, we considered a minimum independence time interval between successive pictures of 30 min and considered those as independent events for subsequent analyses. Although we eventually checked all the pictures to identify individuals, we selected independent events automatically using ExifTool49 from R50 using the library “dplyr”51 and a code (Appendix S1), which allowed us to discriminate those pictures with a temporal difference >30 min. In those cases where several animals were captured in a picture, a different event was considered for each individual.
In the spatially explicit models26 we used the following data: i) a set of marked individuals on the primary occasions (SMR); ii) the capture histories on the secondary occasions of the individuals previously artificially marked on the primary occasions (SMR); iii) the capture histories from all individuals identifiable using natural marks (SCR and SMR); iv) trap and occasion capture histories on secondary occasions from unmarked/unidentifiable individuals (SMR and SC). Precision in the scale or movement parameter sigma (σ) was improved by integrating telemetry data, considering at least 25 locations per individual, except for badgers for which we used home range estimates in similar environments (mean home range of 975 ha, and a standard deviation that covers the home range between 475 and 1,475 ha)52.
Sampling occasion was defined as a sequential 7-day period. We used Poisson encounter models and data augmentation in a Bayesian framework, modified from Royle et al.26, to implement our SCR, SMR and SC models. Details on the spatially explicit Bayesian modelling approaches used are provide in Appendix S2. We modelled the influence of three covariates on density estimates by including them in the baseline encounter rate26. First, we included as a covariate the type of trap used (cameras vs. live-trapping). We included a categorical variable tt[j], which assumed the values 0 or 1 for the traps (live-trapping) and cameras, respectively. Second, we considered the local behaviour of individuals in a binary matrix Lb[i, j, k]26, if we had enough information from individuals (Lbijk was equal to 1 if the individual i was captured at least once prior to session k, otherwise Lbijk was set to 0). This binary matrix was used to account for differential behavioral responses of individuals to survey devices related to different capture histories, for instance, whether past detection events could influence the probability of an individual of being captured again. Finally, we considered a covariate that varied with sampling occasion (t[k]).
We modelled λ0 (baseline detection probability) with a log function:

Therefore α0, α2, α3 and α4 were the parameters to be determined.
Moreover, we integrated in the models an extension to take into account known occurrences of individual removals (i.e., known mortality) with a matrix dead[i, k] indicating when (k-occasion) the individual (i) was extracted from the population. Finally, trap operation (Fig. 3) was included in all models.
The state space (S) is an area that includes the re-sighting grid and is sizable enough to include all individuals potentially exposed to sampling. To generate the state spaces in SMR, we used a buffer around the trap grid from the values of σ and λ0 obtained from a preliminary analysis in each case26.
For model selection, we used the Kuo & Mallick indicator (w)53 variable selection approach to select the best candidate model in relation to the use of different parameters in the models26, and we evaluated the sensitivity of posterior model probabilities to different prior specifications (normal norm(0, 0.1) and uniform unif(−100, 100)). We also used the spike and slab approach54. All models were run in NIMBLE55,56. We ran 3 chains of the MCMC sampler with at least 50,000 iterations in each case (see details for each species in Tables S1–S6). To check for chain convergence, we assessed MCMC convergence by visually inspecting trace plots for each monitored parameter, and we calculated the Gelman-Rubin statistic  57 using the R package “coda”58 where values below 1.1 indicated convergence. For all parameters in our models,
57 using the R package “coda”58 where values below 1.1 indicated convergence. For all parameters in our models,  was always <1.1. Details of the models are in Appendix S3. Different informative priors for sigma could influence our results. Therefore, for red fox, Egyptian mongoose and badger, we additionally explored the influence of different priors on estimates for
was always <1.1. Details of the models are in Appendix S3. Different informative priors for sigma could influence our results. Therefore, for red fox, Egyptian mongoose and badger, we additionally explored the influence of different priors on estimates for  and σ (Appendix S4). For red fox and Egyptian mongoose, 95% BCI were smaller when using an informative prior for sigma compared to use a non-informative prior for sigma (Appendix S4). On the other hand, for badger, 95% BCI in
and σ (Appendix S4). For red fox and Egyptian mongoose, 95% BCI were smaller when using an informative prior for sigma compared to use a non-informative prior for sigma (Appendix S4). On the other hand, for badger, 95% BCI in  was smaller when using the informative prior for sigma (described in Appendix S3) compared to an informative prior for sigma gamma distributed (Appendix S4). We therefore selected these informative priors for sigma in our models.
was smaller when using the informative prior for sigma (described in Appendix S3) compared to an informative prior for sigma gamma distributed (Appendix S4). We therefore selected these informative priors for sigma in our models.
Ethics statement
All field procedures, including animal trapping, telemetry, feral cat euthanasia and camera surveys, were carried out in accordance with animal welfare regulations. Experimental protocols were approved by the Regional Government of Extremadura, Spain, under permit CN0035/13/ACA.
Additional Information
How to cite this article: Jiménez, J. et al. Estimating carnivore community structures. Sci. Rep. 7, 41036; doi: 10.1038/srep41036 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Nichols, J. D. & Williams, B. K. Monitoring for conservation. TREE 21, 668–673 (2006).
Jones, J. P., Asner, G. P., Butchart, S. H. & Karanth, K. U. The ‘why’, ‘what’ and ‘how’ of monitoring for conservation In Key Topics in Conservation Biology 2 (eds Macdonald, D. W. & Willis, K. J. ) 327–343 (John Wiley & Sons, Oxford 2013).
Ahumada, J. A., Hurtado, J. & Lizcano, D. Monitoring the status and trends of tropical forest terrestrial vertebrate communities from camera trap data: A tool for conservation. PLOS ONE 8, e73707 (2013).
Roemer, G. W., Gompper, M. E. & Van Valkenburgh, B. The Ecological Role of the Mammalian Mesocarnivore. BioScience, 59, 165–173 (2009).
Ripple, W. J. et al. Status and ecological effects of the world’s largest carnivores. Science 343, 1241484 (2014).
López-Bao, J. V. & González-Varo, J. P. Frugivory and spatial patterns of seed deposition by carnivorous mammals in anthropogenic landscapes: A multi-scale approach. PLOS ONE 6, 19–21 (2011).
González-Varo, J. P., López-Bao, J. V. & Guitián, J. Functional diversity among seed dispersal kernels generated by carnivorous mammals. J. Anim. Ecol. 82, 562–571 (2013).
Hughes, J. & Macdonald, D. W. A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Cons. 157, 341–351 (2013).
Moseby, K. E., Peacock, D. E. & Read, J. L. Catastrophic cat predation: A call for predator profiling in wildlife protection programs. Biol. Cons. 191, 331–340 (2015).
Boitani, L. & Powell, R. A. Carnivore ecology and conservation: a handbook of techniques (Oxford University Press 2012).
O’Connell, A. F., Nichols, J. D. & Karanth, K. U. Camera traps in animal ecology: methods and analyses (Springer Science & Business Media 2010).
Rich, L. N., Kelly, M. J., Sollmann, R., Noss, A. J., Maffei, L. et al. Comparing capture–recapture, mark–resight, and spatial mark–resight models for estimating puma densities via camera traps. J. Mammal. 95, 382–391 (2014).
Schwarz, C. J. & Seber, G. A. F. Estimating animal abundance: review III. Stat. Sci. 14, 427–456 (1999).
Wilson, G. J. & Delahay, R. J. A review of methods to estimate the abundance of terrestrial carnivores using field signs and observation. Wildlife Res. 28, 151–164 (2001).
Sollmann, R., Gardner, B., Parsons, A. W., Stocking, J. J., McClintock, B. T. et al. A spatial mark-resight model augmented with telemetry data. Ecology 94, 553–559 (2013).
Hayward, M. W. & Marlow, N. Will dingoes really conserve wildlife and can our methods tell? J. Appl. Ecol. 51, 835–838 (2014).
Obbard, M. E., Howe, E. J. & Kyle, C. J. Empirical comparison of density estimators for large carnivores. J. Appl. Ecol. 47, 76–84 (2010).
Ahumada, J. A., et al. Community structure and diversity of tropical forest mammals: data from a global camera trap network. Philos. T. Roy. Soc. B 366, 2703–2711 (2011).
Tobler, M. W., Carrillo-Percastegui, S. E., Leite Pitman, R., Mares, R. & Powell, G. An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Anim. Conserv. 11, 169–178 (2008).
Meek, P. D., et al. Recommended guiding principles for reporting on camera trapping research. Biodivers. Conser. 23, 2321–2343 (2014).
Meyer, N. F. V., et al. An assessment of the terrestrial mammal communities in forests of Central Panama, using camera-trap surveys. J. Nat. Conserv. 26, 28–35 (2015).
Karanth, K. U. & Nichols, J. D. Estimation of tiger densities in India using photographic captures and recaptures. Ecology, 79, 2852–2862 (1998).
López-Bao, J. V., Rodríguez, A. & Palomares, F. Competitive asymmetries in the use of supplementary food by the endangered Iberian lynx (Lynx pardinus). PLOS ONE 4, 1–10 (2009).
Chandler, R. B. & Royle, J. A. Spatially-explicit models for inference about density in unmarked populations. Ann. Appl. Stat. 7, 936–954 (2013).
MacKenzie, D. I., Nichols, J. D., Royle, J. A., Pollock, K. H., Bailey, L. L. & Hines, J. E. Occupancy estimation and modeling. Inferring patterns and dynamics of species occurrence (Elsevier Inc. 2006).
Royle, J. A., Chandler, R. B., Sollman, R. & Gardner, B. Spatial Capture-Recapture (Elsevier/Academic Press 2014).
Efford M. G. & Fewster R. M. Estimating population size by spatially explicit capture– recapture. Oikos 122, 918–928 (2013).
Burton, A. C., Neilson, E., Moreira, D., Ladle, A., Steenweg, R. et al. Wildlife camera trapping: a review and recommendations for linking surveys to ecological processes. J. Appl. Ecol. 10, 1365–2664 (2015).
Chandler, R. Unmarked [Workshop–Athens GA 3/2015]. http://sites.google.com/site/spatialcapturerecapture/workshop-athens-ga-3-2015/day-4. (2015). Date of access: 23/07/2015.
Ramsey, D. S. L., Caley, P. A. & Robley, A. Estimating population density from presence-absence data using a spatially explicit model. J. Wildl. Manage. 79, 491–499.
Cusack, J. J., Dickman, A. J., Rowcliffe, J. M., Carbone, C., Macdonald, D. W. & Coulson, T. Random versus game trail-based camera trap placement strategy for monitoring terrestrial mammal communities. PLOS ONE 10, e0126373 (2015).
García-Peiró, I., Robledano-Aymerich, F., Ángel, M. & Selma, E. Abundancias y densidades relativas de zorro Vulpes vulpes (Linnaeus, 1758) en un humedal del sudeste ibérico: implicaciones para la conservación de sus poblaciones. Anales de Biología, 31, 43–48 (2009).
Sarmento, P., Cruz, J., Eira, C. & Fonseca, C. Evaluation of camera trapping for estimating red fox abundance. J Wildl. Manage., 73, 1207–1212 (2009).
Palomares, F. & Delibes, M. Some physical and population characteristics of Egyptian mongooses (Herpestes ichneumon L., 1758) in southwestern Spain. Z. Saugetierkunde 57, 94–99 (1992).
Revilla, E., Delibes, M., Travaini, A. & Palomares, F. Physical and population parameters of Eurasian badgers (Meles meles L.) from Mediterranean Spain. Z. Saugetierkunde 64, 269–276 (1999).
Rosalino, L., Macdonald, D. W. & Santos-Reis, M. Spatial structure and land-cover use in a low-density Mediterranean population of Eurasian badgers. Can. J. Zool. 1502, 1493–1502 (2004).
Munuera, D. C. & Llobet, F. L. Space use of common genets Genetta genetta in a Mediterranean habitat of northeastern Spain: Differences between sexes and seasons. Acta Theriol. 49, 491–502 (2004).
Peris, A., Tena, L. & Villena, A. Abundancia de ginetas (Genetta genetta) en un encinar mediterráneo. Estimación mediante trampeo fotográfico. Galemys 23, 73–79 (2011).
Sarmento, P. & Cruz, J. A spatially explicit approach for estimating space use and density of common genets. Anim. Biodivers. Conserv. 1, 23–33 (2014).
Viota, M., Rodríguez, A., López-Bao, J. V. & Palomares, F. Shift in microhabitat use as a mechanism allowing the coexistence of victim and killer carnivore predators. OJE 2, 115–120 (2012).
Soto, C. & Palomares, F. Coexistence of sympatric carnivores in relatively homogeneous Mediterranean landscapes: functional importance of habitat segregation at the fine-scale level. Oecologia 179, 223–235 (2015).
Gopalaswamy, A., Delampady, M., Karanth, K. U., Kumar, N. S. & Macdonald, D. W. An examination of index-calibration experiments: counting tigers at macroecological scales. Methods Ecol. Evol. 6, 1055–1066 (2015).
Ferreira, C. & Alves, P. Influence of habitat management on the abundance and diet of wild rabbit (Oryctolagus cuniculus algirus) populations in Mediterranean ecosystems. Eur. J. Wildl. Res. 55, 487–496 (2009).
Monterroso, P., Alves, P. Ć. & Ferreras, P. Evaluation of attractants for non-invasive studies of Iberian carnivore communities. Wildlife Res. 38, 446–454.
Sun, C. C., Fuller, A. K. & Royle, J. A. Trap configuration and spacing influences parameter estimates in spatial capture-recapture models. PLOS ONE 9, e88025 (2014).
Royle, J. A., Kéry, M. & Guélat, J. Spatial capture-recapture models for search-encounter data. Methods Ecol. Evol. 2, 602–611 (2011).
Palomares, F. & Delibes, M. Ecología comparada de la gineta Genetta genetta (L.) y el meloncillo Herpertes ichneumon (L.) en Doñana (SO Península Ibérica). Bol. R. Soc. Esp. Hist. Nat. Secc. Biol. 87, 257–266 (1991).
Monterroso, P., Brito, J. C., Ferreras, P. & Alves, P. C. Spatial ecology of the European wildcat in a Mediterranean ecosystem: dealing with small radio-tracking datasets in species conservation. J. Zool. 279, 27–35 (2009).
Harvey, P. EXIFtool. http://www.sno.phy.queensu.ca/~phil/exiftool/ (2015).
The R Development Core Team. R: A language and environment for statistical computing (R Foundation for Statistical Computing Vienna Austria 2014).
Wickham, H. & Francois, R. dplyr: A Grammar of Data Manipulation. http://cran.r-project.org/package=dplyr (2015).
Revilla, E. Organización social del tejón en Doñana. PhD Thesis (Universidad de León 1998).
Kuo, L. & Mallick, B. Variable selection for regression models. Sankhya Ser. B 60, 65–81 (1998).
Mitchell, T. J. & Beauchamp, J. J. Bayesian variable selection in linear regression. J. Amer. Statist. Assoc. 83, 1023–1032 (1988).
NIMBLE Development Team. NIMBLE: An R Package for Programming with BUGS models (2015).
Valpine, P. De, Turek, D., Paciorek, C. J., Lang, D. T. & Bodik, R. Programming with models: writing statistical algorithms for general model structures with NIMBLE. J. Comp. Graph. Stat., doi: 10.1080/10618600.2016.11724871–20 in press.
Gelman, A., Carlin, J. B., Stern, H. S. & Rubin, D. B. Bayesian data analysis 2nd edn (CRC/Chapman&Hall 2013).
Plummer, M., Best, N., Cowles, K. & Vines, K. CODA: Convergence diagnosis and output analysis for MCMC. R news 6, 7–11 (2006).
Acknowledgements
We are in debt to the staff of the Regional Government of Extremadura, and the Spanish Ministry of Agriculture, Food and Environment, for their collaboration and the logistical support they provided. We thank Albert Roura and Antonio López for their assistance during live-trapping sessions. We wish to thank two anonymous referees for their comments and suggestions. J.V.L.B. was supported by a ‘Juan de la Cierva’ research contract (JCI-2012-13066) from the Spanish Ministry of Economy and Competitiveness.
Author information
Authors and Affiliations
Contributions
J.J. and J.V.L.B. discussed and designed this study based on previous fieldwork carried out by J.J., L.M.G., F.G.D. and J.M.I.; J.C. and C.R. collected the data and contributed with reagents. J.J. analysed the data. J.V.L.B. and J.J. wrote the manuscript with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Jiménez, J., Nuñez-Arjona, J., Rueda, C. et al. Estimating carnivore community structures. Sci Rep 7, 41036 (2017). https://doi.org/10.1038/srep41036
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep41036
This article is cited by
-
Top-down and bottom-up effects modulate species co-existence in a context of top predator restoration
Scientific Reports (2023)
-
Comparing minimum number of individuals and abundance from non-invasive DNA sampling and camera trapping in the red fox (Vulpes vulpes)
Biodiversity and Conservation (2023)
-
Does the size of a protected area matter? An assessment of leopard population and habitat usage in a protected area of Shiwalik foothills, Himalaya
Mammal Research (2023)
-
Shedding light on long-term trends in Mediterranean carnivore populations: five species, one scenario, different responses
European Journal of Wildlife Research (2023)
-
Efficacy of camera traps in detecting primates in Hue Saola Nature Reserve
Primates (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
