
Abstract
An urgent need exists to identify more brown planthopper (Nilaparvata lugens Stål, BPH) resistance genes, which will allow the development of rice varieties with resistance to BPH to counteract the increased incidence of this pest species. Here, using bioinformatics and DNA sequencing approaches, we identified a novel BPH resistance gene, LOC_Os06g03240 (MSU LOCUS ID), from the rice variety Ptb33 in the interval between the markers RM19291 and RM8072 on the short arm of chromosome 6, where a gene for resistance to BPH was mapped by Jirapong Jairin et al. and renamed as “Bph32”. This gene encodes a unique short consensus repeat (SCR) domain protein. Sequence comparison revealed that the Bph32 gene shares 100% sequence identity with its allele in Oryza latifolia. The transgenic introgression of Bph32 into a susceptible rice variety significantly improved resistance to BPH. Expression analysis revealed that Bph32 was highly expressed in the leaf sheaths, where BPH primarily settles and feeds, at 2 and 24 h after BPH infestation, suggesting that Bph32 may inhibit feeding in BPH. Western blotting revealed the presence of Pph (Ptb33) and Tph (TN1) proteins using a Penta-His antibody, and both proteins were insoluble. This study provides information regarding a valuable gene for rice defence against insect pests.
Similar content being viewed by others

Introduction
Rice (Oryza sativa L.) is the primary source of calories for more than one-third of the world’s population, particularly in Asia. Rice crops are often attacked by many diseases and insects, resulting in substantial yield losses. Since the 1960 s, among insect pests, the brown planthopper (Nilaparvata lugens Stål, BPH) has become one of the most important pests in the rice-growing areas of Asia1. BPH not only directly damages rice by sucking phloem sap and by ovipositing in plant tissues, but also transmits viral diseases such as grassy stunt virus (RGSV) and ragged stunt virus (RRSV)2. Although different trends of damage have been observed in Asian countries, problems involving rice planthopper are intensifying3,4. The excessive use of insecticides has led to the development of insecticide resistance in BPH and has disrupted the ecological balance in the rice ecosystem, representing a key factor in the increased incidence of the BPH3,5,6.
A sound balance between breeding for resistance and pest management for BPH management is important; the objective of such management is to reduce the ecological fitness of the BPH, thereby keeping its numbers below economic threshold levels7. However, frequent changes in the biotypes and populations of BPH are among the major challenges for rice breeders. For example, the BPH resistance genes Bph1 and bph2 rapidly became obsolete in just 3–5 years because of the development of new BPH biotypes7. Since the first reports of the BPH resistance genes Bph1 and bph2 by Athwal et al.8, at least 30 new BPH resistance loci have been identified from cultivated and wild species of Oryza7,9,10,11,12,13,14,15,16,17,18. To date, four of these resistance genes, Bph1419, BPH2611, Bph320 and BPH2912, have been cloned. Bph14 and BPH26 both encode a coiled-coil, nucleotide-binding-site, leucine-rich repeat (CC-NBS-LRR) protein19,21. BPH29 contains a B3 DNA-binding domain12. Three BPH resistance genes were inferred to activate the salicylic acid signaling pathway11,12,19. Bph3 is a cluster of three BPH resistance genes encoding plasma membrane-localized lectin receptor kinases (OsLecPK1- OsLecPK3)20. These studies provide insights into the molecular mechanisms of plant-insect interactions and serve as resources for molecular BPH-resistant rice breeding.
The gene Bph3, was first designated by Lakshminarayana and Khush and identified in the rice variety Rathu Heenati22. Subsequent studies have found that Bph3 displays resistance to four BPH biotypes23,24 and is currently still effective despite more than 30 years of deployment20,25,26,27. However, the map position of the Bph3 locus on the rice chromosome has been disputed. The Bph3 gene has primarily been reported to be either allelic or tightly linked to another BPH resistance gene, bph4, in the rice variety Babawee28. This allelic relation was further confirmed by Angeles et al.29, and these two allelic BPH resistance genes were first located on chromosome 7 according to trisomic analysis30. However, the Bph3 gene was subsequently found to be physically located on chromosome 4 based on florescence in situ hybridization31. Although a major BPH resistance gene, tentatively referred to as Bph17, was identified from the rice variety Rathu Heenati on chromosome 432, the gene that was thereafter named Bph3, was cloned by Yuqiang Liu et al.20. However, Jirapong Jairin et al. mapped the Bph3 gene to the short arm of chromosome 6 using two backcross populations, BC1F2 and BC3F2, from crosses of Ptb33 × RD6 and Rathu Heenati × KDML10533. Furthermore Bph3 was physically mapped to an approximately 190-kb interval flanked by the markers RM19291 and RM807234. The BPH resistance locus has widely been used in BPH resistance breeding via marker-assisted selection (MAS)35,36,37, revealing that the locus contains another valuable BPH resistance gene.
In the present study, a dominant gene against BPH, Bph32, was cloned from the rice variety Ptb33 in an approximately 190-kb interval flanked by the markers RM19291 and RM8072 on the short arm of chromosome 6 using bioinformatics analysis and a transgenic approach. The evaluation of BPH resistance in transgenic plants confirmed the crucial function of Bph32 in BPH resistance. Bph32 encodes a short consensus repeat (SCR) domain-containing protein that confers an antibiosis resistance to BPH and is localized in the plasma membrane of the cell. This gene is highly expressed in the leaf sheaths, where the BPH first settles and feeds. The results confirm that Bph32 is a stable BPH resistance gene and provides a valuable gene for rice defence against insect pests.
Results
Identification of Bph32
To identify the Bph32 gene, an approximately 190-kb nucleotide sequence from Nipponbare and an approximately 170-kb nucleotide sequence from 9311, flanked by the simple sequence repeat (SSR) markers RM19291 and RM8072, were analysed using bioinformatics methods. Totals of 39 and 31 putative genes were identified from the 190 and 170-kb fragments, respectively, using the Fgenesh gene-finder38 at http://linux1.softberry.com/berry.phtml?topic=fgenesh&group=programs&subgroup=gfind (Table S1). Eight previously reported putative genes34 were removed from the predicted putative genes. The protein domains of 31 remaining genes were detected using SMART39,40 at http://smart.embl-heidelberg.de/ (Table S1). The LOC_Os06g03240 (MSU LOCUS ID) encoding 194 amino acids was chosen because its protein contained a scop dig6xa_ domain, which was described as a Kunitz/Bovine pancreatic trypsin inhibitor domain (Table S2a and Fig. 1a).

Identification of Bph32 using bioinformatics and DNA sequencing.
(a) Comparison of the genomic fragments flanked by the SSR markers, RM19291 and RM8072 in the 9311 and Nipponbare genomes, and comparison of the cDNA sequences of the LOC_Os06g03240 gene in Ptb33, IR60, IR70, 195B, 121216, TN1, Nipponbare and 9311 revealed that all of these resistant varieties/lines have identical nucleotides and several nucleotide polymorphisms and multiple nonsynonymous nucleotide polymorphisms in the susceptible TN1, Nipponbare and 9311 lines. (b) Comparison of the LOC_Os06g03240 protein sequences in Ptb33 and TN1 revealed the presence of amino acid substitutions, deletions, insertions and different protein domains and binding sites between the two varieties. (c) The 3D protein structure of LOC_Os06g03240 in Ptb33 based on a template, 3DEE_A, with a −9.19 Z-Score using Alignment Mode. Six α-helixes and six β-sheets are present.
Sequence comparison of the LOC_Os06g03240 exons was performed for the resistant varieties Ptb33, IR60, IR70, 195B, and 121216 and for the susceptible varieties, Taichung Native (TN1), Nipponbare and 9311. The LOC_Os06g03240 gene had an identical sequence in all the resistant varieties/lines but displayed several nucleotide polymorphisms compared with the susceptible TN1, Nipponbare and 9311. These polymorphisms included, two deletions of 6 bp(119–124 bp) and 3 bp(544–546 bp); a 9-bp insertion between 389 and 397 bp; and multiple nonsynonymous nucleotide polymorphisms, such as G/C in the 41st bp, G/T in the 91st bp, and T/G in the 105th bp (Fig. 1a and Figure S1). Moreover, short insertions and deletions of several nucleotides (InDels) were detected in the promoter of LOC_Os06g03240 between Ptb33 and TN1 (Figure S1). Sequence comparison of the LOC_Os06g03240 protein between Ptb33 and TN1 revealed 25 amino acid substitutions, 2 deletions and one 3-amino-acid insertion (Fig. 1b and Figure S2). The LOC_Os06g03240 gene in Ptb33 encodes an unknown protein with a molecular weight (MW) of 21658.34 Da and an isoelectric point (pI) of 7.71. This protein contains a signal peptide and a SCOP (Structural Classification of Proteins) d1gkna2 domain belonging to the complement control module/SCR domain, which is considered to be a type of lectin or cell adhesion protein41. Plant lectin are among the most important direct defence proteins in plants against attack by insect pests42. The LOC_Os06g03240 protein of Ptb33 contains seven protein-binding (SO:0000410)43,44 and three polynucleotide-binding regions (SO:0001429)44,45 (Table S2b and Fig. 1b). The LOC_Os06g03240 gene in TN1 encodes a protein with an MW of 21,789.62 Da and a pI of 7.67, that contains twelve protein-binding regions (SO:0000410)43,44 (Table S2b and Fig. 1b). The two LOC_Os06g03240 proteins from Ptb33 and TN1 are predicted to be localized to the plasma membrane of the cell46.
Based on a template, 3DEE_A, the 3D structure of the LOC_Os06g03240 protein in Ptb33 was successfully modeled with a Z-Score of −9.1947 using the Alignment Mode of the SWISS-MODEL workspace. This 3D structure contains six α-helixes and six β-sheets (Fig. 1c). However, an attempt to model the 3D structure of the LOC_Os06g03240 protein in TN1 using the Alignment Mode was unsuccessful. These observations led us to speculate that the LOC_Os06g03240 gene, renamed as “Bph32” in this study, might be a candidate gene for conferring BPH resistance.
Bph32 confers an antibiosis resistance to BPH
To confirm that LOC_Os06g03240 confers BPH resistance, we transformed a susceptible indica variety, Kasalath (Ka), with the cDNA sequence of the Ptb33 Bph32 gene. Six independent transgenic events were detected using genomic southern blotting. Among them, N65-7-1-1-8 contains a single-copy of Bph32 and N65-2-5-2-6 contains a double-copy. Their T2:3 plants were used to measure the levels of resistance of Bph32. As demonstrated in Fig. 2, upon infestation with BPH at the seedling stage, all the wild-type control (Kasalath and TN1) and empty-vector transgenic plants died, whereas the transgenic rice plants expressing the Ptb33 Bph32 gene survived (Fig. 2a,b and c). When infected at the maturing stage, the wild-type plants exhibited leaf wilting, a decrease in seed and grain plumpness, and even death of the whole plant, whereas the Bph32 transgenic plants were all healthy (Fig. 2d).

Complementation test of the Bph32 gene and characterization of BPH resistance in Bph32 transgenic rice.
(a) BPH resistance test of Bph32 transgenic and susceptible wild-type (WT) rice. TN1, susceptible variety (CK); Ka, susceptible WT rice; N65-7-4-3-5, empty-vector transgenic T2:3 line; N65-2-5-2-6 and N65-7-1-1-8, Bph32 transgenic T2:3 lines. (b) BPH resistance scores of Bph32 transgenic T2:3 lines using the modified standard seedbox screening. The data are presented as means ± SD (three replications). (c) RT-PCR analysis showing Bph32 expression in the transgenic T2:3 lines. (d) BPH resistance test of the Bph32 transgenic plants (N65-7-1-1-8) and susceptible wild-type (WT) plants (Ka) at the mature stage. Magnified views showed the locations of BPH feeding. (e) BPH survival number in the Bph32 transgenic plants (N65-7-1-1-8) and susceptible wild-type (WT) plants (Ka) from the first to the twelfth days after BPH infestation. (f) Comparison of the honeydew area in TN1, Ka, N65-7-1-1-8 and Ptb33 using the honeydew excretion test. **P < 0.01. One-way ANOVA was used to generate the P value.
Plants have evolved three defence mechanisms for responding to insect attack: antixenosis repels or disturbs insect settling, thereby reducing colonization or oviposition; antibiosis reduces insect feeding, survival or growth rate; and tolerance helps a crop to maintain high quality and yield after insect infestation48. To explore how Bph32 is involved in plant resistance to BPH, we conducted assays to compare nymph survival and honeydew excretion between the resistant Bph32 transgenic and susceptible wild-type plants (Table S3 and Fig. 2e and f). In the nymph survival test, the number of nymphs steadily decreased on the transgenic plants but began to stabilize 8 days after infestation on the wild-type controls (Fig. 2e). Honeydew excretion in the honeydew area of the transgenic plants was significantly lower than that of the wild-type controls or TN1 plants. However, the difference in honeydew excretion between the transgenic plants and the Ptb33 plants was insignificant (Fig. 2f). These results demonstrate that BPH feeding was inhibited on the resistant Bph32 transgenic plants and suggest that Bph32 conferred an antibiosis resistance that reduced BPH feeding and survival.
Expression analysis of Bph32 and subcellular localization of the Bph32 protein
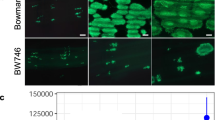
To reveal the molecular mechanisms underlying Bph32-mediated BPH resistance, we examined the expression profile of the Bph32 gene. Real-time (RT) PCR analysis showed that Bph32 was expressed in all investigated tissues at the flowering stage, and its expression level was highest in leaf sheaths followed by leaf blades, culms, panicles and roots (Fig. 3a), consistent with the preference of BPH to settle and probe in leaf sheaths at the flowering stage49. Bph32 expression was further analysed in more detail using transgenic plants carrying a Bph32 promoter-driven GUS reporter gene. GUS expression was observed in the root, leaf blade, leaf sheath, culm, glume, flower, immature seed and germinating seed (Fig. 3b), and GUS activity was strongly detected in parenchyma cells and the vascular bundle (Fig. 3b1 and 3b2). Notably, the expression levels of the Bph32 gene in some tissues of the susceptible line were different from that of the resistant line, which may be associated with the different in 5′ regulatory sequences (Figure S1a).

Bph32 expression analysis.
(a) Comparison of Bph32 expression as measured by RT-PCR in different organs in BPH-resistant 195B (red bar) and BPH-susceptible163B (blue bar). (b) Bph32 promoter-GUS expression pattern in transgenic rice plants. GUS expression profiles in root, culm, leaf blade, leaf sheath, glume, flower, immature seed and germinating seed, respectively (scale bars are as follows: 1–4, 500 μm; 5, 7 and 8, 800 μm; 6, 100 μm). (c) Bph32 subcellular localization. i–ii, Localization of the empty vector. Fluorescence (i) and merged image (ii) of the red fluorescence channel is shown in the top panel. iii–iv, Onion epithelial cells expressing the Bph-ptb33 fusion protein. Fluorescence (iii) and merged (iv) images showing that the Bph32 protein was localized mainly in the plasma membrane. (Scale bar: 100 μm) (d) Comparison of Bph32 expression by RT-PCR after BPH infestation in 195B (red bar) and 163B (blue bar). **P < 0.01; *P < 0.05. One-way ANOVA was used to generate the P value.
To confirm the subcellular localization of Bph32 protein, the Bph32 cDNA fused to the red fluorescent protein (RFP) gene at the N-terminal end and under the control of the CaMV 35S promoter was transiently expressed in onion epidermal cells. Consistent with our prediction, the Bph32 protein was observed in the plasma membrane of the cells (Fig. 3c).
To investigate how Bph32 responds to BPH infestation, we examined Bph32 expression at 0, 2, 4, 8, 24 and 48 h after infestation using RT-PCR. Bph32 expression in the resistant plant 195B was significantly induced compared with that in the susceptible plant 163B at 2 and 24 h after infestation (Fig. 3d). These results support the notion that the sequence variations in the coding and the promoter regions of the Bph32 genes between the resistant and susceptible varieties accounted for the difference in gene function regarding BPH resistance (Figure S1a).
Western blotting revealed that the Pph (Pyb33) and Tph (TN1) proteins were successfully expressed in Escherichia coli (E. coli) BL21T1R and could be detected using a Penta-His antibody (Fig. 4). The two proteins were insoluble and differed slightly in size.

Western blotting of Bph32 protein expression in E. coli BL21T1R.
M1, Precision Plus protein standard; 1, 2, and 3, whole cell lysate, supernatants and sediments of E. coli cells harbouring the control pClod l, respectively; 4, 5, and 6, whole cell lysate, supernatants and sediments of E. coli cells harbouring CDG0933-2(Ptb33, Pph), respectively; 7, 8, and 9, whole cell lysate, supernatants and sediments of E. coli cells harbouring CDG0934-1(TN1:Tph), respectively; M2, Perfect Protein Marker.
Phylogenetic relationship among the Bph32 proteins and the characterization of 195B, a Ptb33 introgression line with BPH resistance
Comparison of the Bph32 cDNA sequence of 123 rice varieties and accessions revealed the following 8 alleles of the Bph32 sequence (Table S4): the TN1 genotype: bph32 (64), the Ptb33 genotype: Bph32 (21), the 389B genotype: bph32389B (15), the Kasalash genotype: bph32ka (8), the Oryza rufipogon genotype: bph32Or (7), the TCHAMPA genotype: bph32TC (4), the P-35 genotype: bph32p (2) and the Safut Khosha genotype: bph32SK (2) (the numbers in brackets are the numbers of rice varieties and accessions with 100% identity in the Bph32 sequence). Bph32 in Ptb33 shares 100% sequence identity with its allele in Oryza latifolia (Table S4, Fig. 5, Figure S3), the wild rice species in South and Central America and an important contributor to BPH resistance17,50,51. The levels of BPH resistance of 119 rice varieties and accessions were investigated in our screen experiment, and BPH resistance scales of the Bph32-containing varieties and accessions were all below 5 (Table S4). BLAST searches performed on http://www.uniprot.org/blast/ showed that 34 Bph32 homologs are found in Oryza (Or), its outgroup (Leersia perrieri), Sorghum bicolor (Sb), Arundo donax (ARUDO), Aegilops tauschii (F775), Triticum urartu (TRIUA), Brachypodium distachyon (BRADI) and Setaria italica. Phylogenetic analysis revealed that Bph32 encodes a unique complement control module, or SCR domain-containing protein, and is closely related to other Oryza homologs and other Gramineae crops (Fig. 5).

Phylogenetic relationships of Bph32 homologs.
Among 123 rice varieties and accessions, Bph32 has eight alleles: the TN1 genotype: bph32 (64), the Ptb33 genotype: Bph32 (21), the 389B genotype: bph32389B (15), the Kasalash genotype: bph32ka (8), the Oryza rufipogon genotype: bph32Or (7), the TCHAMPA genotype: bph32TC (4), the P-35 genotype: bph32p (2) and the Safut Khosha genotype: bph32SK (2). The numbers in brackets represent the numbers of rice varieties and accessions with 100% identity in the Bph32 sequence. Or, Oryza; Sb, sorghum bicolor; ARUDO, Arundo donax; F775, Aegilops tauschii; TRIUA, Triticum urartu; BRADI, Brachypodium distachyon (scale bar: 0.1 amino acid substitutions per site).
To utilize Bph32 in BPH resistance breeding, the maintainer line 195B (F14) containing Bph32 was selected from the progeny of a backcross Ptb33/163B//163B through marker-assisted selection and insect identification. The BPH resistance of 195B and its combination was stable, and the average scores of BPH resistance were both 3.6 (Tables S3 and S4, Fig. 1a, Fig. 6a and b). The genetic background of the line 195B was analysed using 426 SSR markers and BPH resistance gene markers. Among the 426 SSR markers, 371 produced DNA fragments, but only164 SSR markers (44.2% of the markers) amplified polymorphic fragments in Ptb33 and 163B. The line 195B contained 36 Ptb33 polymorphic SSR loci or 21.95% with the Ptb33 genetic background (Table S5 and Fig. 7). In previous reports, Ptb33 has been reported to have two or three BPH resistance genes (bph2 and Bph3 or bph2, Bph3 and Bph9)52,53,54,55. Using gene markers and gene resequencing, we confirmed that Ptb33 contains the three genes Bph32, Bph3p (an allele of Bph3) and BPH26 (bph2), which are associated with BPH resistance (data to be reported elsewhere), whereas 195B contained only one gene, Bph32 (Fig. 6c,d and e, Fig. 7, Figure S4). The 115 F2:3 lines of a population derived from a cross between 195B and 106B(susceptible), showed a fit to a 1:2:1 (26:60:29) ratio (χ2 = 0.19, P = 0.91) for resistant, segregating and susceptible, further confirming that BPH resistance is under the control of a single gene. These results demonstrate that Bph32 is a stable and dominant BPH resistance gene and is functional in conferring BPH resistance (Table S3 and S4, Fig. 6a and b).

BPH resistance of the maintainer line 195B and its combinations by genotypic analysis and phenotypic reaction.
(a,b) BPH resistance test of 195B at the mature and seedling stages. 1, 2, 3, 4, 5 and 6 represent 195 A/Chenghui3203, Chenghui3203, 195B, 163 A/Chenghui3203, Ptb33 and TN1, respectively. (c–e) Gene marker analysis of 195B. (c,d), Gene markers Bph3-In1 and Bph3-In2 for Bph3; (e) gene marker Bp26-INd for BPH26.

Genotypic analysis of the 195B line.
The 195B line contained 36 Ptb33 polymorphic SSR loci (red font) or 21.95% Ptb33 genetic background, and one BPH resistance gene Bph32 (the 115 F2:3 lines of a 195B and 106B (susceptible) cross showed a fit to 1:2:1 (26:60:29) ratio (χ2 = 0.19, P = 0.91) for resistant, segregating and susceptible). Bph320, BPH2510, qBPH(t)57, BPH26(Bph2)11, and Bph982,83 are marked on this genetic map based on previous reports.
Discussion
Bph32, a novel BPH resistance gene, encodes an unknown SCR domain-containing protein
To date, some BPH resistance genes and a BPH quantitative resistance locus (QRL) have been found on the short arm of rice chromosome 616: Bph333,34, bph456, BPH2510 (bph20(t)15) and qBPH(t)57. The rice variety Ptb33 showed a higher degree and a broader spectrum of BPH resistance than has been described previously24,29,33,58. E.R. Angeles et al. first confirmed that Ptb33 contains two BPH resistance genes: Bph3 and bph229. The bph2 gene was mapped within a 1.0-cM region delimited by two AFLP markers, KAM3 and KAM5, and was identified as a single dominant gene59. Later research confirmed that the sequence of bph2 derived from ASD7 is completely identical to the sequence of BPH26 derived from ADR5211. We also confirmed that the sequence of bph2 derived from Ptb33 was completely identical to the sequence of BPH2611 using gene markers (Fig. 6e) and gene resequencing (data not shown). An allele of Bph320 was also identified in Ptb33 using the same approach (Fig. 6c and d) (data to be reported elsewhere). The quantitative resistance locus (QRL) qBPH(t), unlike Ptb33, containing the BPH resistance locus on chromosome 6, was identified in IR71033-121-15, and is flanked by the SSR markers RM469 and RM58657. The bph20(t) gene was renamed BPH25 by Myint et al.10 Although BPH25 and Bph32 are located in similar positions, they did not seem to be allelic because we did not find alleles against BPH (Figure S3). The BPH resistance gene flanked by the markers RM19291 and RM8072 was renamed Bph32 because Bph3 has already been designated.
Our results reveal that the BPH resistance gene Bph32 is a novel gene that encodes an unknown protein containing a signal peptide and a SCOP d1gkna2 domain (Table S2a and Fig. 1b). The SCOP d1gkna2 domain belongs to the complement control module/SCR domain family, a family of cell adhesion molecules (CAMs) that are considered to be types of lectin, or cell adhesion proteins41. Plant lectins have previously been reported to function as direct defence proteins that can act on insect glycoproteins or tissues to inhibit insect feeding42,60. In the present study, honeydew excretion was much lower in Bph32 transgenic plants than wild-type control plants (Fig. 2f). The 3D protein structure of Bph32 was successfully modeled based on a template (3DEE_A) from Neisseria gonorrhoeae Fa 109061 (Fig. 1c). The protein was predicted, and subsequently confirmed, by a subcellular localization experiment, to be localized to the plasma membrane (Fig. 3c). Expression analysis showed that Bph32 was highly expressed in the leaf sheaths, where BPH primarily settles and feeds49 (Fig. 3a). Bph32 was also highly expressed at 2 h and 24 h after BPH infestation (Fig. 3d), which suggested that Bph32 might inhibit BPH feeding (Fig. 2f). The interaction between rice and the BPH mirrors the co-evolution between plants and their natural enemies62,63. Unlike Bph14, BPH26, Bph3 and BPH29, Bph32 encodes an unkown SCR domain-containing protein, thus providing a new insight into the molecular mechanisms underlying plant defences against insect pests.
The Pph (Pyb33) and Tph (TN1) proteins were detected using a Penta-His antibody. The observed difference in migration between the two proteins might be due to differences in their insolubility (Fig. 4), and the two proteins are currently being purified for further analysis of their bioactivity.
Bph32, a stable and dominant BPH resistance gene, offers a resource for rice BPH resistance breeding
Since 1982, several Ptb33-derived, BPH-resistant rice varieties including IR60, IR62, and IR70, have been released, most of which presumably contain the Bph3 locus for resistance based on results from laboratory BPH biotypes in seedbox tests25. The Bph3 locus was successively mapped to rice chromosomes 730, 431 and 633,34 and was eventually cloned from rice chromosome 420. However, the BPH resistance locus, on the short arm of Ptb33 chromosome 6, was confirmed through SSR markers35,36,37 and was verified in the present study to be the Bph32 gene, which is stable and dominant for BPH resistance (Tables S3 and S4, Figs 1a, 2 and Fig. 6a and b).
The Bph32 gene shares 100% sequence identity with its allele in Oryza latifolia (Figure S3). Oryza latifolia, with a CCDD genome, is distributed throughout South and Central America and is an important contributor to BPH resistance17,50,51. Phylogenetic analysis of Bph32 showed that high levels of natural variation exist in Oryza (Fig. 5). However, only the Ptb33 genotype controls BPH resistance. Host genetic background influences the function of the resistance genes64. Different levels of BPH resistance were found for different materials (Table S4). Because of the frequent change in BPH biotypes and the occurrence of insecticide resistance in BPH, rice planthopper problems are intensifying. Thus, we need to develop rice varieties that exhibit stable and durable resistance to BPH by pyramiding multiple resistance genes using MAS. Characterization of the BPH resistance gene Bph32 should greatly assist efforts to develop and deploy rice varieties that exhibit stable and durable resistance to BPH.
Methods
Plant and insect materials
The 123 varieties of cultivated rice and rice accessions used in this study are listed in Table S4 together with their names, countries of origin, taxa and the levels of resistance to brown planthopper (BPH). Of these 123 varieties, the rice variety Ptb33 shows a higher degree and a broader spectrum of BPH resistance than has been described previously58, and the rice variety Ptb33 is the donor of a novel BPH resistance gene, Bph32. The maintainer line 195B containing Bph32 is an F14 selection from the progeny of the backcross Ptb33/163B//163B, and was developed using marker-assisted selection and insect identification. In this cross, the line 163B was found susceptible to BPH. The mixed biotype BPH populations used for infestation were collected from a rice field in Hainan Province, China, and were reared on plants of the susceptible variety Taichung Native (TN1) in a greenhouse maintained under a light regime of 15/9-h light/dark and day/night temperatures of 26–32 °C33.
Identification of Bph32 using bioinformatics and DNA sequencing
Ptb33, an local Indian variety carrying multiple BPH resistance genes, shows a broad-spectrum resistance against all BPH populations and has often been used to verify resistance25,26,27,33. Ptb33 carries the dominant BPH resistance gene Bph322,65. Jirapong Jairin et al. mapped the Bph3 locus between two flanking SSR markers, RM589 and RM588, on the short arm of chromosome 633. Subsequently, the researchers localized the Bph3 gene to a 190-kb interval flanked by the SSR markers RM19291 and RM807234. An approximately 190-kb nucleotide sequence of Nipponbare flanked by the SSR markers RM19291 and RM8072 was downloaded from the GenBank database (http://rice.plantbiology.msu.edu/cgi-bin/gbrowse/rice/). Another approximately 170-kb nucleotide sequence of the Oryza sativa Indica Group was obtained from the Gramene database (http://archive.gramene.org/Oryza_indica/Info/Index). Totals of 39 and 31 putative genes were identified from the two indicated nucleotide sequence fragments, respectively, using the Fgenesh gene-finder38 at http://linux1.softberry.com/berry.phtml?topic=fgenesh&group=programs&subgroup=gfind. Eight previously reported putative genes34 were removed from the predicted putative genes. The protein domains of the remaining genes were detected using SMART39,40 at http://smart.embl-heidelberg.de/. We found that LOC_Os06g03240 (MSU LOCUS ID) contained a SCOP dig6xa_ domain, which was described as a Kunitz/Bovine pancreatic trypsin inhibitor domain. The genomic sequences of LOC_Os06g03240 were further analysed using DNA sequencing in Ptb33, IR70 and IR60 (two Indica varieties known to harbour Bph3)7 and two newly identified rice lines, 195B and 121216, whose BPH resistance is derived from Ptb33, along with the susceptible variety TN1. The genomic DNA fragment (including the promoter regions, the entire CDS region and the downstream sequence) of LOC_Os06g03240 (~2.5 kb) was amplified from Ptb33 seedling DNA using specific primers (Table S6), and the PCR product was verified by DNA sequencing. The predicted protein sequence characteristics of the product, such as MW, pI, and binding sites, were calculated using ExPASy66 (http://web.expasy.org/cgi-bin/compute_pi/pi_tool) and the PredictProtein server44 (https://www.predictprotein.org/home). To model the 3D structure of LOC_Os06g03240, a BLAST search was first performed to identify suitable templates in the Protein Data Bank (pdb) (http://blast.ncbi.nlm.nih.gov/Blast.cgi); thereafter the 3D protein structure was modelled on the SWISS-MODEL workspace using the Alignment Mode67,68,69 (http://www.swissmodel.expasy.org/), and was visualized using the program DeepView70. LOC_Os06g03240 was ultimately considered a candidate gene and was named “Bph32” in this study because the Bph3 on chromosome 4 has previously been cloned by Yuqiang Liu et al.20, and the latest new BPH gene has been designated as Bph31 by G.D. Prahalada et al.71.
Plasmid construction and Western blotting
Total RNA was extracted from Ptb33 seedlings using the TaKaRa MiniBEST plant RNA Extraction Kit (TaKaRa Biotechnology (Dalian) Co., Ltd) according to the manufacturer’s instructions. To construct plasmids for the complementation test, the entire CDS fragment of Bph32 (585 bp) was amplified from Ptb33 seedling total RNA using gene-specific primers (Table S6) from Ptb33 seedling total RNA, and the PCR product was inserted into the binary vector PHB (provided by Shanghai Jiao Tong University) to generate PHB-Bph32.
For subcellular localization, the Bph32 CDS sequence was cloned into the vector pSAT6::RFP-N (purchased from https://www.arabidopsis.org/) and fused with the N-terminus of RFP, resulting in the Bph32-RFP fusion protein-expressing plasmid pSAT6::Bph32-RFP-N. Then, onion epidermal cells were transformed with the OsBph32-RFP gene via particle bombardment. Confocal images were taken 16 h after bombardment using a Nikon A1R-si laser scanning confocal microscope. The experiment was repeated three times.
To prepare the β-glucuronidase (GUS) reporter gene construct, a 786-bp genomic fragment, PtP, corresponding to the 5′ sequence upstream of the ATG start site in the Bph32 gene was amplified from Ptb33 genomic DNA using specific primers, and the PCR product was cloned into the P1300 + PB1101 vector (provided by Shanghai Jiao Tong University) to produce transcriptional fusion, P1300 + PB1101 + PtP, with the GUS gene. All constructs were verified by DNA sequencing.
To investigate the prokaryotic expression of the Bph32 gene, two 594-bp synthetic DNA sequences, Pph (Ptb33:Bph32) and Tph (TN1:Bph32; including the entire CDS region of Bph32), synthesized by TaKaRa Biotechnology (Dalian) Co., Ltd, were each subcloned into the prokaryotic expression vector pColdI and confirmed by DNA sequencing. The corresponding recombinant expression vectors were named CDG0933-2 (Pph) and CDG0934-1 (Tph). Then CDG0933-2, CDG0934-1 and control pColdI were introduced into the bacterial host Escherichia coli (E. coli) BL21T1R following standard protocols, and expression of the fusion genes was induced with 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) at 15 °C for 22 h or until the OD600 was approximately equal to two. E. coli cells that harboured the target plasmid were suspended in 320 μl of PBS in an appropriate final volume, ultrasonically crushed, and separated by centrifugation at 12,000 rpm for 10 min. Finally, three extract fractions (whole E. coli cell lysate, supernatant and sediment) were lysed in SDS sample buffer for 10 min at 99 °C. The lysates were analysed using 15% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE); the proteins were subsequently stained with Coomassie brilliant blue (CBB) R-250 for visualization.
To detect the Bph32 protein, Western blotting was performed as follows. The lysates were subjected to SDS-PAGE, after which the proteins were transferred to a polyvinylidene difluoride (PVDF) membrane. The membrane was incubated first overnight at 4 °C in 10 ml blocking buffer containing 1.5% bovine serum albumin (BSA) and then in 5 ml Penta-His antibody solution containing the primary antibody for 1 h. After washing twice using Tris-buffered saline-Tween (TBST) buffer and three times using TBS buffer, the membrane was incubated again for 1 h with the secondary antibody in 5 ml horseradish peroxidase (HRP)-rabbit anti-mouse IgG solution. The membrane was washed twice using TBST buffer and three times using TBS buffer. The membrane was stained with TrueBlue Peroxidase Substrate for visualization.
Rice transformation
The genetic complementation and Bph32 promoter-GUS fusion constructs were transformed into the Agrobacterium tumefaciens strain EHA105, and were then transformed into the receptor Kasalath (Ka). Plants regenerated from hygromycin-resistant calli (T0 plants) were self-pollinated to produce T1 and T2 seeds. The genotypes of each transgenic plant and their progenies were examined by PCR amplification using gene-specific primers (Table S6). Transgenic plants carrying Bph32 were evaluated for BPH resistance using the above-described methods. GUS activity was detected in transgenic plants by histochemical assay72,73. The experiment was repeated three times.
Evaluation of BPH resistance
BPH resistance was evaluated using the six-scale standard scoring system described by Heinrichs et al.74: 0 = no damage; 1 = very slight damage; 3 = first and second leaves partially yellowing; 5 = pronounced yellowing and stunting; 7 = mostly wilting, the plant still alive; and 9 = the plant completely wilted or dead. Five bioassay methods were used to evaluate BPH resistance in this study. Each experiment was repeated at least twice.
The modified standard seedbox screening method was used to measure the levels of resistance of the donor Ptb33, the susceptible TN1, the transgenic T2:3 plants, the receptor Kasalath (Ka), the introgression line 195B and its hybrid at the seedling stage under greenhouse conditions. Pre-germinated test seeds were soaked in Petri dishes (Ф10 cm) containing tissue paper to ensure that all seedlings were at the same growth stage before BPH infestation. Three days after soaking, the germinating seeds of each material were sown in a 40 × 20 × 2 cm seedbox at a row spacing of 20 × 4 cm with approximately 20 to 30 seeds in each row. At the third-leaf stage, the seedlings were infested with 2nd to 3rd instar BPH nymphs at a density of 8 to 10 insects per seedling. When all TN1 plants had died, the degrees of seedling damage in the varieties or lines were recorded. Each experiment was replicated three times.
Modified adult plant screening was used to evaluate the BPH resistance of the transgenic T2:3, Ka, 195B and 163B lines according to Suh et al.75. Three seedlings at the fourth-leaf stage of the transgenic T2:3, Ka, 195B and 163B lines were transplanted into 25-cm diameter plastic pots containing pulverized soil with compound fertilizer (15-13-12, N-P2O5-K2O) in three replicates. At the heading stage, each adult rice plant was infested with 2nd to 3rd instar nymphs at a density of 200 to 210 insects per plant. The transgenic T2:3 and 195B plants were evaluated based on the degree of damage in the susceptible Ka and 163B control plants, and the plants were recorded as resistant or susceptible once the susceptible controls had died.
In the seedling test, the BPH survival rate was determined to assess the reaction to BPH preference for 195B and 163B. To ensure that all seedlings were at the same growth stage, the seeds were pre-germinated on Petri dishes containing tissue paper. After seven days, the germinating seeds were sown into 15 × 12 cm plastic pots at the rate of one seed per pot. Ten pots were replicated for each treatment. Thirty days after transplanting, the plants were trimmed to one tiller. Then, each plant was infested by 20 2nd instar nymphs and each pot was enclosed in a nylon mesh cage. The number of BPH on each plant was recorded at 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, and 13 days post-infestation.
The honeydew excretion test, as a measure of feeding rate, was conducted to determine the phenotypic reaction of the plants to BPH feeding as described in Heinrichs et al.74. The specific steps used were as follows: seeds were first pregerminated on Petri dishes containing tissue paper for seven days. Three germinating seeds of each material were transplanted into one plastic pot, which was filled with clay as the medium. Ten plastic pots were replicated for each material. The tillers and pollutants were removed from the rice plants 30 days after the rice seeds were sown. Single tillers of the 30-day plants were transferred into 10-cm-diameter plastic cups equipped with honeydew-deposit chambers, which formed a semi-circular dome-like cover and a flat cover, at the rate of one plant per pot. Ten pots were replicated for each treatment. The base of each plant was encircled by a filter paper disk containing 0.02% bromocresol green. Female 4th instar nymphs were starved for 2 h in a nylon mesh cage containing a moist filter paper and were transferred to the chambers (one nymph in each chamber), and the filter papers were collected 24 h later. This process was repeated three times, and the total area of blue-green spots was measured using the Image-Pro Plus 6 program.
A modified bulk seedling screening was used to evaluate the BPH resistance of 119 varieties of cultivated rice, rice accessions, and the 115 F2:3 lines of a population derived from a cross between 195B and 106B (susceptible) at the seedling stage under greenhouse conditions. Approximately 20 to 30 seeds of each material were pre-germinated in small nylon mesh bags for three days and were then sown (8 × 12 cm) in 100 cm × 150 cm pots. Each material was replicated twice. At the third-leaf stage, the seedlings were infested with 2nd and 3rd instar nymphs at a density of 8 to 10 insects per seedling. Each pot was then covered with a 100 × 150 cm nylon mesh cage. When all of the TN1 plants had died, the degrees of seedling damage in the other varieties or accessions were recorded.
Sequence variation analysis
The exons of Bph32 from 123 varieties of cultivated rice and rice accessions were obtained by PCR amplification and DNA sequencing using gene-specific primers (Table S6). The DNA sequences were translated using the Fgenesh gene-finder38 at http://linux1.softberry.com/berry.phtml?topic=fgenesh&group=programs&subgroup=gfind. Homologous proteins of Bph32 were screened by BLAST searching at http://www.uniprot.org/blast/. Phylogenetic analysis was performed using the Clustal Omega program76,77,78 (http://www.uniprot.org/align/ or http://www.ebi.ac.uk/Tools/msa/clustalo/). The phylogenetic tree was built using Mega 6.0579.
Genotypic analysis of line 195B
Four hundred twenty-six SSR markers that were distributed evenly on 12 rice chromosomes were chosen. The SSR primers and the InDel marker primers Bp3-In1and Bp3-In2 for Bph3 and Bp26-INd for BPH26 (Table S6) were synthesized by Sangon Biotech (Shanghai) Co., Ltd. The genetic characterization of 195B was analysed using these SSR and InDel markers. A genetic linkage map of 195B was constructed using the Mapdraw program80 based on the results of PCR amplification and the physical genetic distance of these SSR markers as reported by http://www.gramene.org/.
RNA isolation and qPCR analysis
Pre-germinated seeds of 195B and 163B were sown in 15-cm- diameter pots at the density of one seed per pot. Each plant was infested with 20 BPHs 45 days after sowing and each pot was then covered with a nylon mesh cage. The leaves and sheaths of each plant were collected and frozen immediately in liquid nitrogen at 0, 2, 4, 8, 24 and 48 h after infestation. All treatments, each with three biological replicates, were terminated at the same time. At the heading stage, the flag leaves, culms, sheaths, and roots of 195B and 163B (each line with three biological replicates) were collected and immediately frozen in liquid nitrogen before total RNA isolation. Total RNA was extracted using a TaKaRa MiniBEST Plant RNA Extraction Kit according to the manufacturer’s instructions. Total RNA was then converted into first-strand cDNA using an AMV First Strand cDNA Synthesis Kit following the manufacturer’s instructions. Quantitative RT-PCR was performed using a LightCycler480 Software system (Roche) using the primers listed in Table S6 for the expression analysis of Bph32. The RT-PCR data were analyzed using the 2−ΔΔCT method81.
Additional Information
How to cite this article: Ren, J. et al. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 6, 37645; doi: 10.1038/srep37645 (2016).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Cheng, J. Rice planthopper problems and relevant causes in China. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia, 157–178 (Int. Rice Res. Ins., Manila, Philippines, 2009).
Reissig, W. Illustrated guide to integrated pest management in rice in tropical Asia. (Int. Rice Res. Ins., Manila, Philippines, 1985).
Catindig, J. et al. Situation of planthoppers in Asia. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia, 191–220 (Int. Rice Res. Ins., Manila, Philippines, 2009).
Cheng, J. Rice planthoppers in the past half century in China in Rice Planthoppers. 1–32 (Springer, 2015).
Sheth, P. R. Rice planthoppers strike back. Curr. Sci. 95, 441 (2008).
Cheng, J. & Zhu, Z. Analysis on the key factors causing the outbreak of brown planthopper in Yangtze Area, China in 2005. Plant Protection 32, 1–4 (2005).
Brar, D., Virk, P., Jena, K. & Khush, G. Breeding for resistance to planthoppers in rice. In Planthoppers: new threats to the sustainability of intensive rice production systems in Asia, 401–409 (Int. Rice Res. Inst., Manila, Philippines, 2009).
Athwal, D., Pathak, M., Bacalangco, E. & Pura, C. Genetics of resistance to brown planthoppers and green leafhoppers in Oryza sativa L. Crop Sci. 11, 747–750 (1971).
Huang, D. et al. Fine mapping and characterization of BPH27, a brown planthopper resistance gene from wild rice (Oryza rufipogon Griff.). Theor. Appl. Genet. 126, 219–229 (2013).
Myint, K. K. et al. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor. Appl. Genet. 124, 495–504 (2012).
Tamura, Y. et al. Map-based cloning and characterization of a brown planthopper resistance gene BPH26 from Oryza sativa L. ssp. indica cultivar ADR52. Sci. Rep. 4, 5872 (2014).
Wang, Y. et al. Map-based cloning and characterization of BPH29, a B3 domain-containing recessive gene conferring brown planthopper resistance in rice. J. Exp. Bot. 66, 6035–6045 (2015).
Wu, H. et al. Fine mapping of brown planthopper (Nilaparvata lugens Stål) resistance gene Bph28 (t) in rice (Oryza sativa L.). Mol. Breed. 33, 909–918 (2014).
Rahman, M. L. et al. High-resolution mapping of two rice brown planthopper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minuta. Theor. Appl. Genet. 119, 1237–1246 (2009).
Yang, L. et al. Genetic mapping of bph20 (t) and bph21 (t) loci conferring brown planthopper resistance to Nilaparvata lugens Stål in rice (Oryza sativa L.). Euphytica 183, 161–171 (2012).
Fujita, D., Kohli, A. & Horgan, F. G. Rice resistance to planthoppers and leafhoppers. Crit. Rev. Plant Sci. 32, 162–191 (2013).
Jena, K. K. & Kim, S.-M. Current status of brown planthopper (BPH) resistance and genetics. Rice 3, 161–171 (2010).
Sarao, P. S. et al. Donors for Resistance to Brown Planthopper Nilaparvata lugens (Stål) from Wild Rice Species. Rice Sci. 23, 219–224 (2016).
Du, B. et al. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 106, 22163–22168 (2009).
Liu, Y. et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 33, 301–305 (2015).
Cha, Y. S. et al. Fine mapping of the rice Bph1 gene, which confers resistance to the brown planthopper (Nilaparvata lugens Stål), and development of STS markers for marker-assisted selection. Mol. Cells 26, 146–151 (2008).
Lakshminarayana, A. & Khush, G. S. New genes for resistance to the brown planthopper in rice. Crop Sci. 17, 96–100 (1977).
Pathak, P. & Heinrichs, E. Selection of biotype populations 2 and 3 of Nilaparvata lugens by exposure to resistant rice varieties. Environ. Entomol. 11, 85–90 (1982).
Khush, G. Studies of varietal resistance in rice to the brown planthopper at the International Rice Research Institute. In Brown planthopper: threat to rice production in Asia, 285–301 (IRRI, 1979).
Peñalver Cruz, A., Arida, A., Heong, K. L. & Horgan, F. G. Aspects of brown planthopper adaptation to resistant rice varieties with the Bph3 gene. Entomol. Exp. Appl. 141, 245–257 (2011).
Bhogadhi, S. C. et al. Screening of Rice Genotypes for Resistance to Brown Plant Hopper Biotype 4 and Detection of BPH Resistance Genes. Int. J. Life Sci. Biotechnol. Pharma Res. 4, 90 (2015).
Jena, M., Panda, R., Sahu, R., Mukherjee, A. K. & Dhua, U. Evaluation of rice genotypes for rice brown plant hopper resistance through phenotypic reaction and genotypic analysis. Crop Protect. 78, 119–126 (2015).
Sidhu, G. & Khush, G. Linkage relationships of some genes for disease and insect resistance and semidwarf stature in rice. Euphytica 28, 233–237 (1979).
Angeles, E., Khush, G. & Heinrichs, E. Inheritance of resistance to planthoppers and leafhoppers In Rice genetic. Proceedings of the International Rice Genetics Symposium, 27–31 May, 1985, Manila, Philippines. 537–549 (IRRI, 1986).
Ikeda, R. & Kaneda, C. Genetic analysis of resistance to brown planthopper, Nilaparvata lugens Stål, in rice. Jpn. J. Breed. 31, 279–285 (1981).
Yan, H., Qin, R., Jin, W., He, G. & Song, Y. Comparative physical mapping of Bph3 with BAC-FISH in Oryza officinalis and O. sativa. Acta Bot. Sin. 44, 583–587 (2001).
Sun, L., Su, C., Wang, C., Zhai, H. & Wan, J. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed. Sci. 55, 391–396 (2005).
Jairin, J., Phengrat, K., Teangdeerith, S., Vanavichit, A. & Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breed. 19, 35–44 (2007).
Jairin, J. et al. Physical mapping of Bph3, a brown planthopper resistance locus in rice. Maejo Int. J. Sci. Technol. 1, 166–177 (2007).
Jairin, J. et al. Development of rice introgression lines with brown planthopper resistance and KDML105 grain quality characteristics through marker-assisted selection. Field Crops Res. 110, 263–271 (2009).
Haining, Y., Improvement, G. C., Shaoli, W. & Xiaoqiong, L. Pyramiding lines conferring resistances to brown planthopper and bacterial leaf blight by marker-assisted selection. Mol. Plant Breed. (2010).
Singh, A. et al. Marker assisted selection: a paradigm shift in Basmati breeding. Indian J.Genet. Plant Breed. 71, 120–128 (2011).
Solovyev, V., Kosarev, P., Seledsov, I. & Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 7, S10 (2006).
Letunic, I., Doerks, T. & Bork, P. SMART: recent updates, new developments and status in 2015. Nucleic Acids Res. 43, D257–D260 (2015).
Letunic, I., Doerks, T. & Bork, P. SMART 7: recent updates to the protein domain annotation resource. Nucleic Acids Res. 40, D302–D305 (2012).
Parham, P. The immune system (2nd ed.). New York:: Garland Science,, 244–245 (2005).
Vandenborre, G., Smagghe, G. & Van Damme, E. J. Plant lectins as defense proteins against phytophagous insects. Phytochemistry 72, 1538–1550 (2011).
Ofran, Y. & Rost, B. ISIS: interaction sites identified from sequence. Bioinformatics 23, e13–e16 (2007).
Rost, B., Yachdav, G. & Liu, J. The predictprotein server. Nucleic Acids Res. 32, W321–W326 (2004).
Rost, P. H. E. K. B. Improvement of DNA- and RNA-Protein Binding Prediction . undefined undefined: undefined-undefined (2012).
Goldberg, T., Hamp, T. & Rost, B. LocTree2 predicts localization for all domains of life. Bioinformatics 28, i458–i465 (2012).
Benkert, P., Biasini, M. & Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 27, 343–350 (2011).
Alam, S. & Cohen, M. Detection and analysis of QTLs for resistance to the brown planthopper, Nilaparvata lugens, in a doubled-haploid rice population. Theor. Appl. Genet. 97, 1370–1379 (1998).
Wang, Y. et al. Penetration into rice tissues by brown planthopper and fine structure of the salivary sheaths. Entomol. Exp. Appl. 129, 295–307 (2008).
Jung-Tsung, W., Heinrichs, E. & Medrano, F. Resistance of wild rices, Oryza spp., to the brown planthopper, Nilaparvata lugens (Homoptera: Delphacidae). Environ. Entomol. 15, 648–653 (1986).
Velusamy, R., Kumar, M. G. & Edward, Y. Mechanisms of resistance to the brown planthopper Nilaparvata lugens in wild rice (Oryza spp.) cultivars. Entomol. Exp. Appl. 74, 245–251 (1995).
Balaravi, P., Sharma, R., Dass, A. & Shenoy, V. Evaluation of rice genotypes for brown planthopper (BPH) resistance using molecular markers and phenotypic methods. Afr. J. Biotechnol. 12, 2515 (2013).
Jannoey, P., Pongprasert, W., Lumyong, S., Roytrakul, S. & Nomura, M. Comparative proteomic analysis of two rice cultivars (Oryza sativa L.) contrasting in Brown Planthopper (BPH) stress resistance. Plant Omics 8, 96–105 (2015).
Madurangi, S. & Ratnasekera, D. Evaluation of resistance of wild rice; Oryza nivara to feeding by Brown planthopper [Nilaparvata lugens (Stål)] populations in Proceedings of International Forestry and Environment Symposium 17 (2012).
Manuwoto, S. Respon biologi wereng batang coklat terhadap biokimia tanaman padi. Jurnal Penelitian Pertanian Tanaman Pangan (2012).
Jairin, J., Sansen, K., Wongboon, W. & Kothcharerk, J. Detection of a brown planthopper resistance gene bph4 at the same chromosomal position of Bph3 using two different genetic backgrounds of rice. Breed. Sci. 60, 71–75 (2010).
Jairin, J. et al. Detection of brown planthopper resistance genes from different rice mapping populations in the same genomic location. Sci. Asia 33, 347–352 (2007).
Saxena, R. C. & Okech, S. H. Role of plant volatiles in resistance of selected rice varieties to brown planthopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae). J. Chem. Ecol. 11, 1601–1616 (1985).
Murai, H. et al. Construction of a high-resolution linkage map of a rice brown planthopper (Nilaparvata lugens Stål) resistance gene bph2. Theor. Appl. Genet. 103, 526–532 (2001).
Michiels, K., Van Damme, E. J. & Smagghe, G. Plant‐insect interactions: what can we learn from plant lectins? Arch. Insect Biochem. Physiol. 73, 193–212 (2010).
Das, D. et al. The structure of the first representative of Pfam family PF09836 reveals a two-domain organization and suggests involvement in transcriptional regulation. Acta Crystallogr. F. 66, 1174–1181 (2010).
Rausher, M. D. Co-evolution and plant resistance to natural enemies. Nature 411, 857–864 (2001).
Ryan, M. & Byrne, O. Plant-insect coevolution and inhibition of acetylcholinesterase. J. Chem. Ecol. 14, 1965–1975 (1988).
Cao, Y. et al. The expression pattern of a rice disease resistance gene Xa3/Xa26 is differentially regulated by the genetic backgrounds and developmental stages that influence its function. Genetics 177, 523–533 (2007).
Ikeda, R. Studies of the inheritance of resistance to the rice brown planthopper (Nilaparvata lugens Stål) and the breeding of resistance rice cultivar. Bull Natl. Agric. Res. Cent. 3, 1–54 (1985).
Gasteiger, E. et al. Protein identification and analysis tools on the ExPASy server. (Springer, 2005).
Arnold, K., Bordoli, L., Kopp, J. & Schwede, T. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics 22, 195–201 (2006).
Guex, N., Peitsch, M. C. & Schwede, T. Automated comparative protein structure modeling with SWISS‐MODEL and Swiss‐PdbViewer: A historical perspective. Electrophoresis 30, S162–S173 (2009).
Schwede, T., Kopp, J., Guex, N. & Peitsch, M. C. SWISS-MODEL: an automated protein homology-modeling server. Nucleic Acids Res. 31, 3381–3385 (2003).
Guex, N. & Peitsch, M. C. SWISS‐MODEL and the Swiss‐Pdb Viewer: an environment for comparative protein modeling. Electrophoresis 18, 2714–2723 (1997).
Prahalada, G. D. et al. Indentification of a novel broad-spectrum resistance gene, BPH31 conferring resistance to BPH biotype of India. International Rice Symposium-IRS2015 NOV 18–20 Hyderabad, pp269 (2015).
Martin, T. et al. The GUS reporter system as a tool to study plant gene expression. GUS protocols: using the GUS gene as a reporter of gene expression (ed. Gallagher, S. R. ) 23–43 (1992).
Gallagher, S. R. GUS protocols: using the GUS gene as a reporter of gene expression. (Academic Press, 2012).
Heinrichs, E., Medrano, F. & Rapusas, H. Genetic evaluation for insect resistance in rice. (Int. Rice Res. Inst., Manila, Philippines, 1985).
Suh, J.-P. et al. Development of elite breeding lines conferring Bph18 gene-derived resistance to brown planthopper (BPH) by marker-assisted selection and genome-wide background analysis in japonica rice (Oryza sativa L.). Field Crops Res. 120, 215–222 (2011).
Sievers, F. et al. Fast, scalable generation of high‐quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7, 539 (2011).
Goujon, M. et al. A new bioinformatics analysis tools framework at EMBL–EBI. Nucleic Acids Res. 38, W695–W699 (2010).
McWilliam, H. et al. Analysis tool web services from the EMBL-EBI. Nucleic Acids Res. 41, W597–W600 (2013).
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729 (2013).
Liu, R. & Meng, J. MapDraw: a microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi chuan= Hereditas/Zhongguo yi chuan xue hui bian ji 25, 317–321 (2003).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. methods 25, 402–408 (2001).
Murata, K. et al. Mapping of a brown planthopper (Nilaparvata lugens Stål) resistance gene Bph9 on the long arm of rice chromosome 12. Cereal Res. Commun. 245–250 (2001).
Chang-Chao, S., Hu-Qu, Z., Chun-Ming, W., Li-Hong, S. & Jian-Min, W. SSR mapping of brown planthopper resistance gene Bph9 in Kaharamana, an indica rice (Oryza sativa L.). Acta Genet. Sin. 33, 262–268 (2006).
Acknowledgements
We thank Dr. Theerayut Toojinda for brown planthopper identification training. This work was supported by the 863 Program of China (2014AA10A600), the National System of Modern Agriculture Industrial Technology(CARS-01-08), the Sichuan Innovation Ability Rising Government Financial Project, and the Sichuan Rice Breeding Project for Tackling Key Problems.
Author information
Authors and Affiliations
Contributions
J.S.R. performed the genetic analysis, constructed the vectors, conducted the sequence and expression analysis and wrote the manuscript; F.Y.G. performed the genetic transformation and Western blotting; X.T.W. performed the subcellular localization; X.J.L., L.H.Z., J.Q.L. and X.W.S. perfromed the phenotypic analysis; and G.J.R. and H.L. directed the project and wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Ren, J., Gao, F., Wu, X. et al. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci Rep 6, 37645 (2016). https://doi.org/10.1038/srep37645
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep37645
This article is cited by
-
Mapping of a novel recessive brown planthopper resistance gene bph46 from wild rice (Oryza nivara)
Euphytica (2024)
-
Dissecting brown planthopper resistance genes in Oryza and its wild relatives: A review
Euphytica (2024)
-
Phenotypic and genotypic assessment of elite rice varieties for brown plant hopper (Nilaparvata lugens Stål.) resistance
Cereal Research Communications (2023)
-
In silico analysis of cloned brown planthopper genes unveiled OsJ_28113 as a key regulator in triggering resistance response in rice
Journal of Proteins and Proteomics (2023)
-
Large scale rice germplasm screening for identification of novel brown planthopper resistance sources
Molecular Breeding (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
